
Abstract
Renal tissue hypoperfusion and hypoxia are early key elements in the pathophysiology of acute kidney injury of various origins, and may also promote progression from acute injury to chronic kidney disease. Here we describe test interventions that are used to study the control of renal hemodynamics and oxygenation in experimental animals in the context of kidney-specific control of hemodynamics and oxygenation. The rationale behind the use of the individual tests, the physiological responses of renal hemodynamics and oxygenation, the use in preclinical studies, and the possible application in humans are discussed.
This chapter is based upon work from the COST Action PARENCHIMA, a community-driven network funded by the European Cooperation in Science and Technology (COST) program of the European Union, which aims to improve the reproducibility and standardization of renal MRI biomarkers.
You have full access to this open access chapter, Download protocol PDF
Similar content being viewed by others

Key words
1 Introduction
Kidney diseases are a global health burden with steadily increasing incidence and prevalence [1,2,3,4,5]. Animal studies indicate that acute kidney injuries (AKI ) of various origins share one common link in the pathophysiological chain of events, ultimately leading to AKI , as well as to progression from AKI to chronic kidney diseases (CKD ): imbalance between renal oxygen delivery and oxygen demand [6,7,8,9,10,11,12,13,14]. Renal tissue hypoperfusion and hypoxia have also been suggested to play a pivotal role in the pathophysiology of other kidney diseases including diabetic kidney disease [15,16,17,18,19]. These pathophysiological concepts have largely been generated by preclinical studies that used either invasive quantitative probes or noninvasive functional magnetic resonance imaging (MRI) techniques to gain insight into renal hemodynamics and oxygenation. Thus, making ultimate statements on the role of renal hypoperfusion and hypoxia for these renal disorders is elusive because in vivo assessment of renal hemodynamics and oxygenation constitutes a challenge.
All modalities available in today’s experimental and translational research practice have inherent shortcomings and methodological constraints. Invasiveness is the major disadvantage of the gold standard physiological probes such as perivascular flow probes for measurement of total renal blood flow, laser-Doppler-optodes for assessment of local tissue perfusion, and Clark-type electrodes or fluorescence-quenching optodes for measurements of local tissue partial pressure of oxygen (pO2), which precludes their use in humans. While functional MRI including blood oxygenation-sensitized T2* (aka blood oxygenation level-dependent MRI; BOLD-MRI) offers noninvasive techniques to obtain insight into renal perfusion and oxygenation, its major weakness is its qualitative nature. Before it can be used for quantitative characterization of renal tissue perfusion and oxygenation, it needs to be calibrated with the gold standard invasive techniques in various (patho)physiological scenarios [20,21,22,23,24].
The control of renal hemodynamics and oxygenation under physiological as well as under pathophysiological conditions is complex and differs considerably from nonrenal tissue [8, 21, 23, 25,26,27]. Due to the considerable capacity of the organism’s homeostatic control systems to—at least partially—compensate for disturbances of, or injury to, certain control elements, these alterations are often not easily detectable when studied by measuring baseline data only. In order to disentangle these complexities, dedicated reversible test interventions are conceptually appealing. In fact, such interventions can serve three main purposes. First, they are used to gain more insight into the control of renal hemodynamics and oxygenation in healthy animals and in animal models of various kidney diseases [8, 12, 25, 28,29,30,31,32,33,34,35,36,37]. Second, the tests are used to assess whether a given drug or contrast agent has beneficial or unwarranted effects on the control of renal hemodynamics and oxygenation [38,39,40]. Finally, dedicated reversible tests are used to achieve calibration of functional MRI data [22, 23].
In this chapter, specifics of the control of renal hemodynamics and oxygenation are outlined first. Then, the individual test procedures are described, and the rationale behind their use, the physiological response of renal hemodynamics and oxygenation, the use in preclinical studies and the possible application in humans are discussed.
This chapter is part of the book Pohlmann A, Niendorf T (eds) (2020) Preclinical MRI of the Kidney—Methods and Protocols. Springer, New York.
2 Specifics of Renal Hemodynamics and Oxygenation
Renal hemodynamics and oxygenation offer a number of striking differences when compared to nonrenal tissue. First, total renal blood flow (RBF) is huge when compared to virtually all other organs on a per gram basis: the kidneys receive about 20% of the cardiac output under resting conditions. Yet the distribution of blood perfusion differs substantially between the layers: while 100% of blood flowing into the kidney reaches the cortex, only 15% of blood that previously passes through the cortex, will reach the medulla. Even intralayer (cortex, outer medulla, and inner medulla) perfusion is quite heterogeneous [8, 41, 42]. In accordance with the high total RBF, the kidneys’ oxygen extraction (the difference between the O2 content in the renal arterial and the renal venous blood) is low as compared to the majority of nonrenal tissues. Yet the partial pressure of oxygen (pO2) is low in the medulla and also varies considerably within the respective layers, in accordance with the different blood flow distribution [21, 23, 26, 43,44,45,46,47].
Second, the kidney differs from all other organs with regard to the relationship between metabolism and perfusion. More than 26 thousand millimoles of sodium (Na+) are filtered in the human glomeruli every day, equivalent to more than 1.5 kg of table salt. To achieve sodium balance, the amount of salt excreted by the kidneys must exactly match the amount of ingested salt minus the amount of extrarenal loss. Thus, more than 99% of the filtered sodium must usually be reabsorbed from the tubules. Tubular resorption relies on active transport processes, which account for about 85% of the kidney’s energy expenditure and therefore its O2 consumption. The more sodium is filtered in the glomeruli, the more must be reabsorbed. As glomerular filtration rate (GFR), under the majority of circumstances, increases with increasing RBF, renal O2 consumption also usually increases with increasing renal perfusion. This is in contradistinction to all other organs, where metabolism determines perfusion [8, 48].
Third, hormones such as angiotensin II and epinephrine, sympathetic vasomotor nerves, and paracrine mediators such as nitric oxide or adenosine, that control resistance vessels in nonrenal tissues, impinge on intrarenal resistance vessels too, thereby altering renal O2 delivery. However, in the kidney, they additionally affect tubular sodium resorption and thus O2 consumption. Furthermore, their effect on postglomerular vessels can result in divergent responses of RBF and GFR . Finally, adenosine exerts vasodilation in virtually all nonrenal vascular beds, but vasoconstriction in the renal cortex [8, 25, 48, 49].
Fourth, the kidney is equipped with efficient mechanisms of autoregulation, that is, the ability to dampen or even to abolish the effects that changes in renal arterial pressure would otherwise inevitably have on RBF and GFR . The almost perfect autoregulation of RBF and GFR probably relies on the fact that not just one, but three mechanisms are involved. The first one, the myogenic response (aka Bayliss effect) acts not only on renal resistance vessels but also on brain and gut vessels. The second mechanism, the tubuloglomerular feedback (TGF), and the third one, hitherto just named “third mechanism”, are kidney-specific. Renal autoregulatory mechanisms, in particular the TGF and the third mechanism, have been suggested to serve the purpose of balancing O2 delivery, that is, RBF with metabolic and O2 demands arising from tubular reabsorption. The outer medulla is particularly prone to imbalance between O2 delivery and demand since this layer exhibits a high O2 demand but low pO2 [21, 34, 35, 50,51,52].
Fifth, intrarenal perfusion is also affected by changes in tubular volume . The tubular volume fraction is quite large and can rapidly change due to alterations in GFR , in tubular outflow toward the pelvis, in tubular fluid resorption, and modulation of the transmural pressure gradient. Since the renal capsule is rather tough, changes in tubular volume will result in circular distension or compression of intrarenal vessels [23, 36].
Finally, in addition to the heterogeneous intrarenal blood perfusion, three other factors substantially contribute to the low tissue pO2 and, in particular, to the “physiological hypoxia” in the medulla. First, there is a considerable shunt diffusion of O2 from arteries to veins in the cortex and from descending to ascending vasa recta in the medulla [53,54,55]. Second, the Fåhræus–Lindqvist effect lowers the hematocrit in the vasa recta supplying the medulla, which lowers the O2 content of blood perfusing parts of the medulla [41, 42]. Third, plasma skimming at intrarenal vessel branches results in different hematocrit and therefore O2 content of blood perfusing the daughter vessels [41, 56].
3 Dedicated Reversible Test Interventions
3.1 Short Periods of Occlusion of the Renal Artery or Renal Vein
Occlusions of the renal artery (alternatively: the suprarenal aorta) or of the renal vein emulate clinical conditions in which deficient renal perfusion results in deterioration of intrarenal oxygenation. If maintained for longer periods of time these conditions can cause AKI [57,58,59]. The rationale for performing both of these tests is that renal arterial occlusion and renal venous occlusion have similar effects with regard to renal perfusion and oxygenation, yet opposing effects with regard to intrarenal blood volume . With the onset of aortic occlusion, the inflow of blood into the kidney is abruptly stopped while outflow via the renal vein continues until pressures in intrarenal vessels and in the vena cava are equalized. With the onset of renal venous occlusion, outflow of blood is abruptly stopped while inflow via the artery does not cease until the arterial pressure-induced distension of intrarenal vessels is counterbalanced by the resistance of the renal tissue including the rather tough capsule [22, 39, 60].
In both cases, renal tissue perfusion rapidly decreases and eventually approaches zero flow. As renal O2 consumption remains unaltered at the early stage of occlusions, a rapid and massive decline in renal tissue pO2 results, which, in turn, also reduces blood pO2 and the O2 saturation of hemoglobin (StO2) in the intrarenal (micro)vasculature. This intrarenal deoxygenation of hemoglobin (Hb) is aggravated by a progressive rightward shift of the oxyHb dissociation curve during the occlusion due to the intrarenal accumulation of carbon dioxide (CO2) [22, 39, 60].
Yet the opposing changes of renal blood volume have an impact on renal tissue oxygenation. The decrease in tissue pO2 at the onset of the venous occlusion is much slower than at the onset of the arterial occlusion. While renal O2 consumption is similar during both kinds of occlusions, the transiently maintained inflow of oxygenated blood at the onset of venous occlusion increases the intrarenal reservoir of O2 [22, 39, 60].
The opposing changes in renal blood volume have a massive impact on the changes in blood oxygenation-sensitized T2* (and its reciprocal value, R2*), because T2* reflects the amount of deoxygenated Hb (deoxyHb) per tissue volume (voxel) [23]. In case of the venous occlusion with its increase in the vascular volume fraction and thus the increasing amount of deoxyHb per volume , tissue T2* massively decreases [60]. With the arterial occlusion’s decrease of deoxyHb per volume , the decrease in T2* is small. In fact, it was found significantly smaller than the decrease in T2* measured during hypoxemia (8% inspiratory oxygen fraction), which is diametrically opposed to the effects of arterial occlusion versus hypoxemia on tissue pO2 (see Fig. 1) [22, 23, 36].

Comparison of relative changes in renal cortical and medullary tissue pO2 quantified by invasive gold standard fluorescence quenching optodes (left panel) versus relative changes in renal cortical and medullary T2* (so-called BOLD-MRI, right panel), during short-term occlusion of the suprarenal aorta and short-term hypoxia (8% inspiratory O2 fraction), respectively, in anesthetized rats. Data are mean ± SEM, redrawn from Refs. 22, 36
Short-time (1–3 min) occlusions of the renal artery (or of the suprarenal aorta) have been used in several studies for different scientific purposes. In order to gauge the effects on T2* and T2 of bolus injections of an X-ray contrast medium into the thoracic aorta of healthy rats, the effect of arterial occlusion (and that of hypoxemia) was quantified in the same rats [36]. En route to calibration of T2* with quantitative physiological measurements by means of a dedicated hybrid MR-PHYSIOL setup (see the chapter by Cantow K et al. “Monitoring Renal Hemodynamics and Oxygenation by Invasive Probes: Experimental Protocol”), suprarenal aortic occlusion was used [22]. In order to ascertain that the superparamagnetic iron oxide nanoparticle (USPIO) preparation, ferumoxytol is suitable as a contrast medium for MR-based assessment of the renal blood volume fraction; its possible unwarranted effects on control of renal hemodynamics and oxygenation were tested by interventions including suprarenal aortic occlusion in rats (see Fig. 2) [38]. Implementing a setup that combines classical invasive probes for RBF, tissue perfusion, and pO2 with newly developed near infrared spectroscopy (NIRS) techniques that enable monitoring of the amount of Hb per tissue volume and the O2 saturation of Hb (StO2) of intrarenal blood (termed PHYSIOL-NIRS), aortic occlusion was used as one of the test interventions [39]. By means of a dedicated deconvolution procedure developed by our group, the time course of RBF upon the release of the occlusion can be analyzed. This “step-response” analysis allows us to determine the strength of each of the three mechanisms of RBF autoregulation in the whole kidney in vivo, in both healthy rats and rat models of AKI [30, 34, 35].

In order to study whether the USPIO preparation, ferumoxytol (FO), exerts unwarranted effects on regulation of renal hemodynamics and oxygenation, a short-term suprarenal aortic occlusion was employed as test intervention in anesthetized rats [38]. Here, the relative changes (mean ± SEM) in hemodynamics and tissue oxygenation are depicted with FO dosages of 6, 10, and 41 mg Fe/kg body mass, or vehicle (Control)
Short-time (1–3 min) occlusions of the renal vein were also used for different purposes. In order to establish an optimum dose of the USPIO ferumoxytol in rats for the purpose of T2*-based quantification of the renal blood volume fraction in a 9.4T small animal scanner, renal venous occlusion was chosen as the combined effects of the decrease in O2 delivery and the increase in the blood volume fraction; thus, deoxyHb per volume was expected to result in a most prominent decrease in T2* (see Fig. 3) [60].

Left panel: T2*-weighted images (echo time = 3.6 ms, spatial resolution = 226 × 422 μm) obtained by a 9.4 T small animal MR scanner (Bruker Biospin, Biospec 94/20) of a rat kidney in vivo at baseline, during occlusion of the renal vein (v.o.), and at the beginning of the recovery phase (rec.) without the USPIO ferumoxytol (top row), and with four increasing doses of ferumoxytol [60]. Right panels: Comparison of the renal cortical and medullary T2* sensitivity to USPIO injection and the T2* sensitivity to the venous occlusion at different USPIO doses. Data are mean ± SEM (n = 4 rats) of cortical and medullary ROIs [60]
Both renal arterial occlusion and renal venous occlusion was performed in the same healthy rats in the PHYSIOL-NIRS setup in order to directly compare their effects [39].
As the implementation of vascular occluders necessitates invasive techniques, these tests can be performed in preclinical studies only.
3.2 Servocontrolled Changes in Renal Arterial Pressure
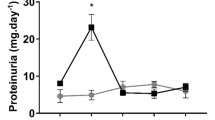
Dynamic changes in renal arterial pressure according to different time courses of pressure reduction followed by pressure restoration—be it staircasewise or rampwise changes—enable insights into control of renal hemodynamics and oxygenation including the degree of autoregulation’s efficiency and the contributions of the three autoregulatory mechanisms in vivo. This is achieved by a servocontrol system developed by our group, that was utilized to help disentangle the complexities of renal physiology and pathophysiology [30, 31, 33, 34]. Moreover, a study that employed such an intervention in a rat model that emulates an early stage of diabetic kidney disease (a type 1 diabetes mellitus–like model induced by administration of streptozotocin 4 weeks before obtaining the data on renal perfusion and oxygenation) unmasked alterations in the control of renal perfusion and oxygenation that would have gone undetected when only baseline data had been obtained [32]. Data on medullary tissue pO2 obtained by invasive probes in this model have been inconsistent, which, among other reasons, may be caused by the spatial heterogeneity of pO2 within the renal medulla [15,16,17]. A recent study did not find any significant differences in baseline data on RBF and cortical and medullary tissue pO2 among healthy control rats, diabetic rats, and diabetic rats treated with the antidiabetic liraglutide (a glucagon-like peptide 1 agonist approved for patients suffering from type 2 diabetes). However, as depicted by Fig. 4, the response to ramp-wise reduction and restoration of renal arterial pressure differed considerably among these groups [32].

Changes of invasively measured parameters of renal hemodynamics and oxygenation during ramp-wise reduction in renal perfusion pressure followed by ramp-wise pressure restoration in anesthetized rats. Conductance values (the reciprocal of vascular resistance) were calculated by dividing the respective perfusion values by renal perfusion pressure, in order to distinguish flow changes that result from passive circular distension/compression of vessels from those actively exerted by vascular smooth muscles. Three groups were studied: a healthy control group, a group in which a diabetes mellitus type 1-like disorder (DM) was induced by streptozotocin 4 weeks before obtaining the data, and a third group in which DM was induced and the antidiabetic liraglutide administered for 3 weeks (DM + LIRA). Values (mean ± SEM) are given as relative changes from baseline [32]
Again, as the implementation of a vascular occluder necessitates invasive techniques, such studies can be performed in preclinical studies only.
3.3 Short Periods of Changes in the Inspiratory Gas Mixture
Hyperoxia, hypoxia, and hypercapnia primarily alter blood oxygenation. Renal O2 delivery is determined by renal perfusion and by the arterial O2 content. The latter is determined, among other factors, by the inspiratory fraction of oxygen (FiO2), and, due to the effect of CO2 on the oxyHb dissociation curve, also by the inspiratory fraction of CO2 (FiCO2).
Increasing the FiO2 from 21% (normoxia) to 100% (hyperoxia) results in a substantial increase in arterial pO2 (usually four- to fivefold), whereas the increase in arterial O2 content is very small, because most of the Hb in arterial blood is already O2 saturated under normoxic conditions. Yet the increase in arterial pO2 enhances the driving force for diffusion of O2 from intrarenal vessels to tissue as well as from intrarenal arteries to veins. As a consequence, the increase in renal tissue pO2 is substantial, whereby medullary pO2 increases less than cortical pO2, due to arteriovenous diffusive O2 shunting, which reduces the O2 content of arterial blood that perfuses the medulla [22, 26, 39, 53,54,55]. Renal T2* changes exerted by hyperoxia are small [22]. While the amount of deoxyHb in arterial blood is barely changed, the increase in blood pO2 in intrarenal veins that results from the higher arteriovenous pO2 difference decreases venous deoxyHb. While primarily altering blood oxygenation, the hyperoxic stimulus has also secondary effects: it results in vasoconstriction, preferentially in nonrenal vascular beds, which leads to an increase in arterial pressure [22, 31, 39].
The primary effect of reducing the FiO2 (typically to either 8% or 10%, with durations of 3–12 min, in rat studies) is a decrease in oxygenation of arterial blood (hypoxemia) with the consequent reduction in renal O2 supply. With ongoing O2 consumption, this does per se result in a decrease in renal tissue pO2. Yet renal O2 supply is further diminished by hypoxia-induced extrarenal vasodilation that results in a drop in arterial pressure with ensuing decrease in RBF [22, 31, 39, 59]. Whether this is aggravated of alleviated by constriction or dilation, respectively, of the renal vasculature depends on the degree of hypoxia: in anesthetized rats, FiO2 of 8% results in renal vasoconstriction while 10% results in vasodilation [22, 31, 39]. The combined effect of hypoxemia and reduced RBF on renal O2 supply leads to a major mismatch with O2 consumption, that massively reduces tissue pO2 as well as T2* (see Fig. 1) [22, 39]. A further secondary effect of arterial hypoxemia is increased ventilation triggered by arterial chemoreceptors. The ensuing decrease in arterial pCO2 shifts the oxyHb dissociation curve to the left, that is, O2 is hindered from being released by Hb, which further aggravates the tissue hypoxia [61, 62].
With the hypercapnic stimulus (increasing FiCO2 to 5%) the opposite effect is achieved, namely a rightward shift of the oxyHb dissociation curve. This would per se result in a decrease in StO2 and an increase in blood and tissue pO2. However, while the increase in tissue pO2 is substantial, the StO2 decrease is meagre [39]. The major reason that StO2 does not decrease much is that increased pCO2 of arterial blood is a very strong stimulus for ventilation, again mediated by arterial chemoreceptors [61, 62].
Hyperoxic, hypoxic, and hypercapnic tests have been used in a multitude of preclinical in vivo studies. With regard to the kidney this includes but is not limited to studies on the control of renal hemodynamics and oxygenation in healthy animals and models of kidney diseases [26, 27, 31, 53,54,55, 59], experiments that aimed at calibration of T2* by means of the MR-PHYSIOL setup (see Fig. 5), [22] studies on the T2* effect of an X-ray contrast medium, [36] assessment of possible unwarranted effects of the USPIO ferumoxytol, [38] and experiments en route to the PHYSIOL-NIRS setup [39].

Time courses of selected invasively measured data and MR parameters acquired simultaneously throughout baseline, a period of hypoxia (FiO2 = 8%), and recovery in anesthetized rats by means of a dedicated MR-PHYSIOL hybrid setup (for details see text). Data (mean ± SEM) are relative changes from baseline, redrawn from Ref. 22
Given the broad therapeutic use of pure oxygen, short-term tests with 100% FiO2 should pose no problem for studies in humans, whereas hypoxic challenges are precluded in humans, for obvious ethical reasons. Hypercapnia has been used for decades in humans, in particular, for the study of cerebrovascular reactivity, and should thus be employed in preclinical and clinical studies on renal hemodynamics and oxygenation as well [63, 64].
3.4 Administration of Drugs and Endogenous Vasoactive Substances
Furosemide is the “classic” loop diuretic: its major action is the inhibition of the sodium –potassium–two-chloride cotransporter in the apical membrane of tubular epithelial cells of the thick ascending limb of Henle’s loop. The primary effect is an increase in urine flow rate and in urinary sodium and potassium excretion. As less tubular resorption necessitates less renal O2 consumption, administration of furosemide leads to an increase in renal tissue pO2 [65, 66]. In accordance, increases in renal T2* (or decreases in its reciprocal value R2*) have been observed in a multitude of preclinical as well as clinical studies (see Fig. 6) [37, 66,67,68,69]. It must be noted, however, that the increase in T2* upon furosemide does not solely rely on improved oxygenation. First, the increase in tubular fluid downstream of the thick ascending limb will increase the transmural pressure gradient, thereby compressing intrarenal vessels with the ensuing decrease in the amount of deoxyHb per tissue volume . Second, furosemide inhibits the TGF, thereby compromising renal autoregulation with the possible consequence of an increase in RBF [34, 35, 50].

Effect of injections of furosemide (5 mg/kg body mass), hydralazine (5 mg/kg), angiotensin II (0.5 μg/min/kg), and saline (repeatability) on renal BOLD as recorded by means of a 1.5 T clinical MR scanner (Magnetom Avanto, Siemens Healthcare), using a multiple gradient echo sequence (TR = 300 ms, TE = 5, 10, 20, 30, and 40 ms, voxel size 0.6 × 0.6 mm in-plane and 5 mm slice thickness) in rats in vivo. Data are mean ± SEM of median ROI values [37]
The use of the furosemide test in both preclinical and clinical MR studies is nowadays as widespread that it is almost regarded as a gold standard. However, whether it fulfils all expectations regarding its use as a diagnostic tool in patients suffering from various kidney diseases, remains to be seen [68].
Administration of furosemide is a reversible intervention insofar as its direct effects vanish with the excretion of the drug. However, it leaves the organism with deficits in water, sodium , and potassium. These should ideally be replenished—be it per os or by means of infusions of a balanced electrolyte solution.
Bolus injections of adenosine cause a rapid drop in arterial pressure due to its vasodilatory effect on nonrenal resistance vessels. In the renal cortical vascular bed, it exerts vasoconstriction [39, 49]. The consequence of these two effects is a substantial decrease in RBF followed by a smaller decrease in cortical tissue pO2. All these effects vanish rapidly, lasting less than a minute for the hemodynamics and less than 2 min for the cortical pO2 in rats [39].
Whereas the role of adenosine in various renal control mechanisms including the TGF as well as the potentially beneficial effect of adenosine receptor antagonists for prevention of X-ray contrast media-induced AKI have been intensively studied, [49, 70] the adenosine test has seldom been used to study renal hemodynamics and oxygenation. This may appear surprising, as injections of adenosine—be it intravenously or into coronary arteries—in patients suffering from coronary disease is quite established [71, 72]. While the risk for a decrease in renal cortical pO2 in patients must not be ignored—even if it is lasting less than 2 min, the test should at least find wider use in preclinical studies.
References
Fortrie G, de Geus HRH, Betjes MGH (2019) The aftermath of acute kidney injury: a narrative review of long-term mortality and renal function. Crit Care 23(1):24. https://doi.org/10.1186/s13054-019-2314-z
Selby NM, Taal MW (2019) Long-term outcomes after AKI-a major unmet clinical need. Kidney Int 95(1):21–23. https://doi.org/10.1016/j.kint.2018.09.005
Zuk A, Bonventre JV (2019) Recent advances in acute kidney injury and its consequences and impact on chronic kidney disease. Curr Opin Nephrol Hypertens 28(4):397–405. https://doi.org/10.1097/mnh.0000000000000504
Levin A, Tonelli M, Bonventre J, Coresh J, Donner JA, Fogo AB, Fox CS, Gansevoort RT, Heerspink HJL, Jardine M, Kasiske B, Kottgen A, Kretzler M, Levey AS, Luyckx VA, Mehta R, Moe O, Obrador G, Pannu N, Parikh CR, Perkovic V, Pollock C, Stenvinkel P, Tuttle KR, Wheeler DC, Eckardt KU (2017) Global kidney health 2017 and beyond: a roadmap for closing gaps in care, research, and policy. Lancet 390(10105):1888–1917. https://doi.org/10.1016/s0140-6736(17)30788-2
Bello AK, Levin A, Tonelli M, Okpechi IG, Feehally J, Harris D, Jindal K, Salako BL, Rateb A, Osman MA, Qarni B, Saad S, Lunney M, Wiebe N, Ye F, Johnson DW (2017) Assessment of global kidney health care status. JAMA 317(18):1864–1881. https://doi.org/10.1001/jama.2017.4046
Brezis M, Rosen S (1995) Hypoxia of the renal medulla--its implications for disease. N Engl J Med 332:647–655
Seeliger E, Sendeski M, Rihal CS, Persson PB (2012) Contrast-induced kidney injury: mechanisms, risk factors, and prevention. Eur Heart J 33(16):2007–2015
Evans RG, Ince C, Joles JA, Smith DW, May CN, O'Connor PM, Gardiner BS (2013) Haemodynamic influences on kidney oxygenation: the clinical implications of integrative physiology. Clin Exp Pharmacol Physiol 40:106–122
Evans RG, Ow CP, Bie P (2015) The chronic hypoxia hypothesis: the search for the smoking gun goes on. Am J Physiol Renal Physiol 308(2):F101–F102
Shu S, Wang Y, Zheng M, Liu Z, Cai J, Tang C, Dong Z (2019) Hypoxia and hypoxia-inducible factors in kidney injury and repair. Cell 8(3):207. https://doi.org/10.3390/cells8030207
Hultstrom M, Becirovic-Agic M, Jonsson S (2018) Comparison of acute kidney injury of different etiology reveals in-common mechanisms of tissue damage. Physiol Genomics 50(3):127–141. https://doi.org/10.1152/physiolgenomics.00037.2017
Calzavacca P, Evans RG, Bailey M, Bellomo R, May CN (2015) Cortical and medullary tissue perfusion and oxygenation in experimental septic acute kidney injury. Crit Care Med 43(10):e431–e439
Fähling M, Seeliger E, Patzak A, Persson PB (2017) Understanding and preventing contrast-induced acute kidney injury. Nat Rev Nephrol 13(3):169–180
Ma S, Evans RG, Iguchi N, Tare M, Parkington HC, Bellomo R, May CN, Lankadeva YR (2019) Sepsis-induced acute kidney injury: a disease of the microcirculation. Microcirculation 26(2):e12483. https://doi.org/10.1111/micc.12483
Palm F, Carlsson PO, Hansell P, Hellberg O, Nygren A, Liss P (2003) Altered response in renal blood flow and oxygen tension to contrast media in diabetic rats. Acta Radiol 44(3):347–353
Palm F, Cederberg J, Hansell P, Liss P, Carlsson PO (2003) Reactive oxygen species cause diabetes-induced decrease in renal oxygen tension. Diabetologia 46(8):1153–1160
dos Santos EA, Li LP, Ji L, Prasad PV (2007) Early changes with diabetes in renal medullary hemodynamics as evaluated by fiberoptic probes and BOLD magnetic resonance imaging. Investig Radiol 42(3):157–162. https://doi.org/10.1097/01.rli.0000252492.96709.36
Calvin AD, Misra S, Pflueger A (2010) Contrast-induced acute kidney injury and diabetic nephropathy. Nat Rev Nephrol 6(11):679–688
Hansell P, Welch WJ, Blantz RC, Palm F (2013) Determinants of kidney oxygen consumption and their relationship to tissue oxygen tension in diabetes and hypertension. Clin Exp Pharmacol Physiol 40(2):123–137
Evans RG, Gardiner BS, Smith DW, O'Connor PM (2008) Methods for studying the physiology of kidney oxygenation. Clin Exp Pharmacol Physiol 35(12):1405–1412
Pohlmann A, Cantow K, Hentschel J, Arakelyan K, Ladwig M, Flemming B, Hoff U, Persson PB, Seeliger E, Niendorf T (2013) Linking non-invasive parametric MRI with invasive physiological measurements (MR-PHYSIOL): towards a hybrid and integrated approach for investigation of acute kidney injury in rats. Acta Physiol (Oxf) 207(4):673–689
Pohlmann A, Arakelyan K, Hentschel J, Cantow K, Flemming B, Ladwig M, Waiczies S, Seeliger E, Niendorf T (2014) Detailing the relation between renal T2* and renal tissue pO2 using an integrated approach of parametric magnetic resonance imaging and invasive physiological measurements. Investig Radiol 49(8):547–560
Niendorf T, Pohlmann A, Arakelyan K, Flemming B, Cantow K, Hentschel J, Grosenick D, Ladwig M, Reimann H, Klix S, Waiczies S, Seeliger E (2015) How BOLD is blood oxygenation-dependent (BOLD) magnetic resonance imaging of the kidney? Opportunities, challenges and future directions. Acta Physiol (Oxf) 213(1):19–38
Hirakawa Y, Tanaka T, Nangaku M (2017) Renal hypoxia in CKD; pathophysiology and detecting methods. Front Physiol 8:99. https://doi.org/10.3389/fphys.2017.00099
Calzavacca P, Evans RG, Bailey M, Bellomo R, May CN (2015) Variable responses of regional renal oxygenation and perfusion to vasoactive agents in awake sheep. Am J Physiol Regul Integr Comp Physiol 309(10):R1226–R1233
Evans RG, Gardiner BS, Smith DW, O'Connor PM (2008) Intrarenal oxygenation: unique challenges and the biophysical basis of homeostasis. Am J Physiol Renal Physiol 295(5):1259–1270
Evans RG, Goddard D, Eppel GA, O'Connor PM (2011) Factors that render the kidney susceptible to tissue hypoxia in hypoxemia. Am J Physiol Regul Integr Comp Physiol 300(4):R931–R940
Emans TW, Janssen BJ, Pinkham MI, Ow CP, Evans RG, Joles JA, Malpas SC, Krediet CT, Koeners MP (2016) Exogenous and endogenous angiotensin-II decrease renal cortical oxygen tension in conscious rats by limiting renal blood flow. J Physiol 594(21):6287–6300. https://doi.org/10.1113/jp270731
Evans RG, Madden AC, Denton KM (2000) Diversity of responses of renal cortical and medullary blood flow to vasoconstrictors in conscious rabbits. Acta Physiol Scand 169(4):297–308
Seeliger E, Flemming B, Wronski T, Ladwig M, Arakelyan K, Godes M, Mockel M, Persson PB (2007) Viscosity of contrast media perturbs renal hemodynamics. J Am Soc Nephrol 18(11):2912–2920
Flemming B, Seeliger E, Wronski T, Steer K, Arenz N, Persson PB (2000) Oxygen and renal hemodynamics in the conscious rat. J Am Soc Nephrol 11(1):18–24
Ferrara F, Cantow K, Flemming B, Skalweit A, Ladwig M, Fähling M, Seeliger E (2017) Effects of liraglutide on control of renal hemodynamics and oxygenation in diabetic rats. Acta Physiol (Oxf) 219(Suppl. 711):38
Flemming B, Arenz N, Seeliger E, Wronski T, Steer K, Persson PB (2001) Time-dependent autoregulation of renal blood flow in conscious rats. J Am Soc Nephrol 12(11):2253–2262
Seeliger E, Wronski T, Ladwig M, Dobrowolski L, Vogel T, Godes M, Persson PB, Flemming B (2009) The renin-angiotensin system and the third mechanism of renal blood flow autoregulation. Am J Physiol Renal Physiol 296(6):F1334–F1345
Wronski T, Seeliger E, Persson PB, Forner C, Fichtner C, Scheller J, Flemming B (2003) The step response: a method to characterize mechanisms of renal blood flow autoregulation. Am J Physiol Renal Physiol 285(4):F758–F764
Arakelyan K, Cantow K, Hentschel J, Flemming B, Pohlmann A, Ladwig M, Niendorf T, Seeliger E (2013) Early effects of an x-ray contrast medium on renal T2*/T2 MRI as compared to short-term hyperoxia, hypoxia and aortic occlusion in rats. Acta Physiol 208(2):202–213
Jerome NP, Boult JK, Orton MR, d'Arcy J, Collins DJ, Leach MO, Koh DM, Robinson SP (2016) Modulation of renal oxygenation and perfusion in rat kidney monitored by quantitative diffusion and blood oxygen level dependent magnetic resonance imaging on a clinical 1.5T platform. BMC Nephrol 17(1):142. https://doi.org/10.1186/s12882-016-0356-x
Cantow K, Pohlmann A, Flemming B, Ferrara F, Waiczies S, Grosenick D, Niendorf T, Seeliger E (2016) Acute effects of ferumoxytol on regulation of renal hemodynamics and oxygenation. Sci Rep 6:29965. https://doi.org/10.1038/srep29965
Grosenick D, Cantow K, Arakelyan K, Wabnitz H, Flemming B, Skalweit A, Ladwig M, Macdonald R, Niendorf T, Seeliger E (2015) Detailing renal hemodynamics and oxygenation in rats by a combined near-infrared spectroscopy and invasive probe approach. Biomed Opt Express 6(2):309–323
Seeliger E, Cantow K, Arakelyan K, Ladwig M, Persson PB, Flemming B (2014) Low-dose nitrite alleviates early effects of an X-ray contrast medium on renal hemodynamics and oxygenation in rats. Investig Radiol 49(2):70–77
Edwards A, Silldforff EP, Pallone TL (2000) The renal medullary microcirculation. Front Biosci 5:E36–E52
Zimmerhackl BL, Robertson CR, Jamison RL (1987) The medullary microcirculation. Kidney Int 31(2):641–647
Schurek HJ (1988) Kidney medullary hypoxia: a key to understanding acute renal failure? die Nierenmarkhypoxie: ein Schlussel zum Verstandnis des akuten Nierenversagens? Klin Wochenschr 66(18):828–835
Baumgartl H, Leichtweiss HP, Lubbers DW, Weiss C, Huland H (1972) The oxygen supply of the dog kidney: measurements of intrarenal pO2. Microvasc Res 4(3):247–257
Lubbers DW, Baumgartl H (1997) Heterogeneities and profiles of oxygen pressure in brain and kidney as examples of the pO2 distribution in the living tissue. Kidney Int 51(2):372–380
Evans RG, Ow CPC (2018) Heterogeneity of renal cortical oxygenation: seeing is believing. Kidney Int 93(6):1278–1280. https://doi.org/10.1016/j.kint.2018.01.039
Hirakawa Y, Mizukami K, Yoshihara T, Takahashi I, Khulan P, Honda T, Mimura I, Tanaka T, Tobita S, Nangaku M (2018) Intravital phosphorescence lifetime imaging of the renal cortex accurately measures renal hypoxia. Kidney Int 93(6):1483–1489. https://doi.org/10.1016/j.kint.2018.01.015
Evans RG, Harrop GK, Ngo JP, Ow CP, O'Connor PM (2014) Basal renal O2 consumption and the efficiency of O2 utilization for Na+ reabsorption. Am J Physiol Renal Physiol 306(5):F551–F560
Vallon V, Muhlbauer B, Osswald H (2006) Adenosine and kidney function. Physiol Rev 86(3):901–940
Just A (2007) Mechanisms of renal blood flow autoregulation: dynamics and contributions. Am J Physiol Regul Integr Comp Physiol 292(1):R1–R17
O'Connor PM (2006) Renal oxygen delivery: matching delivery to metabolic demand. Clin Exp Pharmacol Physiol 33(10):961–967
Blantz RC, Weir MR (2004) Are the oxygen costs of kidney function highly regulated? Curr Opin Nephrol Hypertens 13(1):67–71
Schurek HJ, Jost U, Baumgartl H, Bertram H, Heckmann U (1990) Evidence for a preglomerular oxygen diffusion shunt in rat renal cortex. Am J Physiol 259(6 Pt 2):F910–F915
Ngo JP, Ow CP, Gardiner BS, Kar S, Pearson JT, Smith DW, Evans RG (2016) Diffusive shunting of gases and other molecules in the renal vasculature: physiological and evolutionary significance. Am J Physiol Regul Integr Comp Physiol 311(5):R797–r810. https://doi.org/10.1152/ajpregu.00246.2016
Leong CL, Anderson WP, O'Connor PM, Evans RG (2007) Evidence that renal arterial-venous oxygen shunting contributes to dynamic regulation of renal oxygenation. Am J Physiol Renal Physiol 292:F1726–F1733
Pappenheimer JR, Kinter WB (1956) Hematocrit ratio of blood within mammalian kidney and its significance for renal hemodynamics. Am J Physiol 185:377
Hoff U, Lukitsch I, Chaykovska L, Ladwig M, Arnold C, Manthati VL, Fuller TF, Schneider W, Gollasch M, Muller DN, Flemming B, Seeliger E, Luft FC, Falck JR, Dragun D, Schunck WH (2011) Inhibition of 20-HETE synthesis and action protects the kidney from ischemia/reperfusion injury. Kidney Int 79(1):57–65
Hosszu A, Antal Z, Lenart L, Hodrea J, Koszegi S, Balogh DB, Banki NF, Wagner L, Denes A, Hamar P, Degrell P, Vannay A, Szabo AJ, Fekete A (2017) sigma1-receptor Agonism protects against renal ischemia-reperfusion injury. J Am Soc Nephrol 28(1):152–165. https://doi.org/10.1681/asn.2015070772
Cantow K, Flemming B, Ladwig-Wiegard M, Persson PB, Seeliger E (2017) Low dose nitrite improves reoxygenation following renal ischemia in rats. Sci Rep 7(1):14597–15058
Pohlmann A, Cantow K, Huelnhagen T, Grosenick D, Dos Santos PJ, Boehmert L, Gladytz T, Waiczies S, Flemming B, Seeliger E, Niendorf T (2017) Experimental MRI monitoring of renal blood volume fraction variations En route to renal magnetic resonance oximetry. Tomography 3(4):188–200. https://doi.org/10.18383/j.tom.2017.00012
Kumar P (2009) Systemic effects resulting from carotid body stimulation-invited article. Adv Exp Med Biol 648:223–233. https://doi.org/10.1007/978-90-481-2259-2_26
Lahiri S, Forster RE 2nd (2003) CO2/H(+) sensing: peripheral and central chemoreception. Int J Biochem Cell Biol 35(10):1413–1435
Catchlove SJ, Pipingas A, Hughes ME, Macpherson H (2018) Magnetic resonance imaging for assessment of cerebrovascular reactivity and its relationship to cognition: a systematic review. BMC Neurosci 19(1):21. https://doi.org/10.1186/s12868-018-0421-4
Corfield DR, McKay LC (2016) Regional cerebrovascular responses to hypercapnia and hypoxia. Adv Exp Med Biol 903:157–167. https://doi.org/10.1007/978-1-4899-7678-9_11
Brezis M, Agmon Y, Epstein FH (1994) Determinants of intrarenal oxygenation. I. Effects of diuretics. Am J Physiol 267(6 Pt 2):F1059–F1062. https://doi.org/10.1152/ajprenal.1994.267.6.F1059
Warner L, Glockner JF, Woollard J, Textor SC, Romero JC, Lerman LO (2011) Determinations of renal cortical and medullary oxygenation using blood oxygen level-dependent magnetic resonance imaging and selective diuretics. Investig Radiol 46(1):41–47. https://doi.org/10.1097/RLI.0b013e3181f0213f
Prasad PV, Edelman RR, Epstein FH (1996) Noninvasive evaluation of intrarenal oxygenation with BOLD MRI. Circulation 94(12):3271–3275. https://doi.org/10.1161/01.cir.94.12.3271
Palmucci S, Mammino L, Caltabiano DC, Costanzo V, Foti PV, Mauro LA, Farina R, Profitta ME, Sinagra N, Ettorre GC, Veroux M, Basile A (2019) Diffusion-MR in kidney transplant recipients: is diuretic stimulation a useful diagnostic tool for improving differentiation between functioning and non-functioning kidneys? Clin Imaging 53:97–104. https://doi.org/10.1016/j.clinimag.2018.10.003
Haddock B, Larsson HBW, Francis S, Andersen UB (2019) Human renal response to furosemide: simultaneous oxygenation and perfusion measurements in cortex and medulla. Acta Physiol (Oxf) 227:e13292. https://doi.org/10.1111/apha.13292
Welch WJ (2002) Adenosine A1 receptor antagonists in the kidney: effects in fluid-retaining disorders. Curr Opin Pharmacol 2(2):165–170
Gili S, Barbero U, Errigo D, De Luca G, Biondi-Zoccai G, Leone AM, Iannaccone M, Montefusco A, Omede P, Moretti C, D'Amico M, Gaita F, D'Ascenzo F (2018) Intracoronary versus intravenous adenosine to assess fractional flow reserve: a systematic review and meta-analysis. J Cardiovasc Med (Hagerstown) 19(6):274–283. https://doi.org/10.2459/jcm.0000000000000652
Solerno R, Pedroni P, Mariani J, Sarmiento R (2018) Comparison of sodium nitroprusside and adenosine for fractional flow reserve assessment: a systematic review and meta-analysis. Expert Rev Cardiovasc Ther 16(10):765–770. https://doi.org/10.1080/14779072.2018.1513789
Acknowledgments
This work was funded, in part (Kathleen Cantow and Erdmann Seeliger) by the German Research Foundation (Gefoerdert durch die Deutsche Forschungsgemeinschaft (DFG), Project number/ Projektnummer 394046635, SFB 1365, RENOPROTECTION).The authors wish to thank Ariane Anger and Andrea Gerhardt for expert technical assistance.
This chapter is based upon work from COST Action PARENCHIMA, supported by European Cooperation in Science and Technology (COST). COST (www.cost.eu) is a funding agency for research and innovation networks. COST Actions help connect research initiatives across Europe and enable scientists to enrich their ideas by sharing them with their peers. This boosts their research, career, and innovation.
PARENCHIMA (renalmri.org) is a community-driven Action in the COST program of the European Union, which unites more than 200 experts in renal MRI from 30 countries with the aim to improve the reproducibility and standardization of renal MRI biomarkers.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Open Access This chapter is licensed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license and indicate if changes were made.
The images or other third party material in this chapter are included in the chapter's Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the chapter's Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder.
Copyright information
© 2021 The Author(s)
About this protocol

Cite this protocol
Cantow, K., Ladwig-Wiegard, M., Flemming, B., Fekete, A., Hosszu, A., Seeliger, E. (2021). Reversible (Patho)Physiologically Relevant Test Interventions: Rationale and Examples. In: Pohlmann, A., Niendorf, T. (eds) Preclinical MRI of the Kidney. Methods in Molecular Biology, vol 2216. Humana, New York, NY. https://doi.org/10.1007/978-1-0716-0978-1_4
Download citation
DOI: https://doi.org/10.1007/978-1-0716-0978-1_4
Published:
Publisher Name: Humana, New York, NY
Print ISBN: 978-1-0716-0977-4
Online ISBN: 978-1-0716-0978-1
eBook Packages: Springer Protocols