
Abstract
The use of human organotypic models for biomedical research is experiencing a significant increase due to their biological relevance, the possibility to perform high-throughput analyses, and their cost efficiency. In the field of anti-infective research, comprising the search for novel antipathogenic treatments including vaccines, efforts have been made to reduce the use of animal models. That is due to two main reasons: unreliability of data obtained with animal models and the increasing willingness to reduce the use of animals in research for ethical reasons. Human three-dimensional (3-D) models may substitute and/or complement in vivo studies, to increase the translational value of preclinical data. Here, we provide an overview of recent studies utilizing human organotypic models, resembling features of the cervix, intestine, lungs, brain, and skin in the context of anti-infective research. Furthermore, we focus on the future applications of human skin models and present methodological protocols to culture human skin equivalents and human skin explants.
Astrid Hendriks, Ana Rita Cruz, These authors contributed equally.
The original version of this chapter was revised: OpenAccess text has been included. The erratum to this chapter is available at https://doi.org/10.1007/82_2019_174.
You have full access to this open access chapter, Download chapter PDF
Similar content being viewed by others


1 Introduction
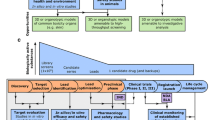
Overall mortality caused by infectious diseases has dramatically decreased in the past decades due to the positive impact of antibiotics, antivirals, antifungals, vaccines, and more recently monoclonal antibodies. The use of in vitro and/or in vivo models in preclinical research aims to predict the level of safety, immunogenicity, and protection of novel anti-infective treatments. Unfortunately, animal models often fail in this respect, as shown by the high number of unsuccessful clinical trials. There is an abundant amount of pathogens with exclusive human-specific properties, thus underlining the need for human in vitro or ex vivo models to study these microbes and to obtain reliable results in preclinical studies. Furthermore, great efforts are being made to reduce, refine, and replace the use of animal models in various fields of biomedical research. Human organotypic models are becoming more popular in both the academic sector as well as industry, producing valuable information to help find novel therapeutic targets. Here, we will present an overview of human organ-specific models used in anti-infective research, focusing on cervical, intestinal, respiratory, brain, and skin models. Moreover, we will dedicate a section of this review to human skin models commenting on their future applications and presenting a methodological protocol for the culture of human skin explants and skin equivalents that can be applied to anti-infective research.
2 Part 1—State of the Art of Organotypic Models for Anti-infective Research
2.1 Cervical Models
Human 3-D organotypic cultures, also known as raft cultures, have been developed in order to mimic human cervical epithelium. These models are frequently composed of primary cervical keratinocytes or cervical carcinoma cell lines seeded on top of a collagen matrix filled with fibroblasts (Delvenne et al. 2001). Organotypic raft cultures are then grown at air–liquid interface in order to promote differentiation and consequent stratification of the epithelium.
Epithelial differentiation is essential for human papillomavirus (HPV) replication (Meyers et al. 1992); thus, cell monolayers are limited models to study the life cycle of HPV and, consequently, to develop antiviral therapies against its replication. For this reason, organotypic epithelial raft cultures have been used to evaluate the efficacy of antivirals in blocking replication of different HPV types. Two commercial preparations of interferon-α (IFN-α) (Sen et al. 2005) and an antisense oligonucleotide targeting the mRNA of an essential protein (ORI-1001) for HPV6 and HPV11 replication (Alam et al. 2005) were tested in this model. These antivirals were able to impair the replication of specific HPV types, and the optimal concentration of the two IFN-α preparations was shown to be different for each of the HPV types. Additionally, the efficacy of bi-specific monoclonal antibodies anti-CD3/anti-EGFR, designed to target epidermal growth factor receptor (EGFR)-positive neoplastic keratinocytes by activating T cells (Ferrini et al. 1993), was also evaluated in a human 3-D model of the cervical epithelium (Renard et al. 2002). After the demonstration that lymphocytes can penetrate in 3-D cultures composed by a cervical carcinoma cell line (Jacobs et al. 1998), the authors proved that T cells, previously incubated with the bi-specific antibodies, induce apoptosis of HPV16-transformed primary keratinocytes, but not of healthy keratinocytes (Renard et al. 2002). These studies highlighted the feasibility and preclinical relevance of organotypic cultures of keratinocytes to test different classes of anti-HPV agents.
Human cervical explants have also been extensively used to screen the efficacy and toxicity of topical antiviral candidates, mainly for human immunodeficiency virus (HIV). The compounds tested include polyanionic molecules, such as the synthetic naphthalenesulfonate polymer PRO2000 and dextrin sulfate (DxS); cellulose acetate 1,2-benzenedicarboxylate (CAP); the carrageenan-based product PC-515; the lysine dendrimer SPL7013; the antimicrobial peptide D2A21; cyanovirin-N; palmitic acid; nonoxynol-9; gramicidin; the pyrimidinedione IQP-0528; and the non-nucleoside reverse transcriptase inhibitor UC781 (Greenhead et al. 2000; Zussman et al. 2003; Fletcher et al. 2005, 2006; Lu et al. 2006; Cummins et al. 2007; Buffa et al. 2009; Lin et al. 2011; Mahalingam et al. 2011). The combinatorial effect of these compounds in preventing HIV-1 infection has also been evaluated in polarized ectocervical explants (Zhang et al. 2015).
Infections caused by HIV are frequently accompanied by other sexually transmitted infections, such as herpes simplex virus (HSV) (Wald and Link 2002; Buve 2010). Highly specific drugs against each of these viruses have been designed, including acyclovir (HSV-specific) and tenofovir (HIV-specific). Surprisingly, clinical trials revealed that acyclovir could reduce HIV-1 levels in HSV-2/HIV-1 coinfected patients (Nagot et al. 2007; Zuckerman et al. 2007; Baeten et al. 2008) and that tenofovir reduced the risk of HSV-2 infection (Cates 2010). The ability of acyclovir to suppress HIV-1 infection was confirmed in human tonsils, lymph nodes, cervicovaginal and colorectal tissues inoculated with HIV-1 and HSV-2 (Lisco et al. 2008), and the effectiveness of tenofovir to impair HSV replication was evaluated in several in vitro and ex vivo models such as organotypic epithelial 3-D rafts, human lymphoid and cervical tissues (Andrei et al. 2011). Therefore, cervical and other types of tissues showed to be suitable models to assess the antiviral effect of microbicides against two different viruses. In a more recent study, the compound PMEO-DAPym (6-phosphonylmethoxyethoxy-2,4-diaminopyrimidine) was also identified to target both HSV and HIV infections (Balzarini et al. 2013). Its potency was also tested in multiple models, including organotypic epithelial raft cultures of human primary keratinocytes and human lymphoid and cervical tissues, showing an overall significant advantage over the existing compounds tenofovir and adefovir, which have been described to impair both HIV and HSV.
In summary, human cervical raft cultures and explants have been exploited as models to test the efficacy of various treatments, to optimize the concentration of microbicides and to assess the benefit of combining antivirals to tackle different viruses. Importantly, organotypic models have been used to overcome the limitations of cell monolayers in reproducing in vivo conditions essential for viral replication.
2.2 Intestinal Models
In the last years, several studies started to use colorectal explant tissues to evaluate the antiretroviral efficacy of topical microbicides marketed or under evaluation in clinical trials for vaginal use. The safety and effectiveness of many antivirals such as CAP, PRO2000, SPL7013, Vena Gel, and the class of reverse transcriptase inhibitors (RTI), which includes the nucleotide-RTI tenofovir, the nucleoside-RTI emtricitabine (FTC) and the non-nucleoside-RTI UC-781, dapivirine (TMC120), and IQP-0528 was assessed in colorectal tissues (Abner et al. 2005; Fletcher et al. 2006; Herrera et al. 2009; Rohan et al. 2010; Herrera et al. 2011; Dezzutti et al. 2012, 2014). When tested in combination, RTI were shown to induce a greater inhibitory potential compared with the compounds tested alone (Herrera et al. 2009, 2011). The effectiveness of other types of drugs, such as the polyanion candidates PRO2000 and DxS, has been also tested in parallel with the RTI activity (Fletcher et al. 2006). More recently, a study combining the non-nucleoside-RTI TMC120 with the HIV entry inhibitor maraviroc proved a synergistic antiretroviral effect in both ectocervical and colonic tissue (Dezzutti et al. 2015). Also, in this case, the use of explant tissue models allows the preclinical assessment of both toxicity and efficacy of topical microbicides, and it facilitates the screening of multiple candidates, alone or in combination, against different viral subtypes, thereby being less laborious than in vivo models.
Another type of human intestinal organotypic model is the 3-D cultured primary intestinal organoid, which is an organ-like tissue generated from pluripotent stem cells isolated from intestinal tissue (Lancaster and Knoblich 2014). Organoids contain various types of cells and recapitulate most aspects of in vivo tissue architecture, like spatial organization and overall microenvironment. Intestinal organoids have been used to study host-pathogen interactions; nevertheless, the number of studies that uses this model to evaluate the efficacy of antimicrobial agents is still limited. Yin et al. showed that primary human intestinal organoids were permissive to rotavirus infection, both to laboratory and patient-derived rotavirus strains. Importantly, clinical isolates demonstrated different sensitivities to the antivirals IFN-α and ribavirin, suggesting that this organotypic model could be valuable to predict the individual responsiveness to antiviral therapy, thus supporting personalized medicine (Yin et al. 2015). Another study implemented a functional assay to determine the efficacy of cholera toxin inhibitors by measuring the swelling of primary human intestinal organoids (Zomer-van Ommen, Pukin et al. 2016).
2.3 Respiratory Models
The respiratory system has a complex architecture and consists of different distinct structures such as the nasal mucosa, the upper and lower bronchial tracts and alveoli. Due to the direct contact with the air, natural exposure to various pathogens is very common. Subsequently, pneumonia remains one of the leading causes of death worldwide, thereby making the respiratory system a key target model for anti-infective research (Global Burden of Disease Study 2015). Certain aspects of the respiratory system, such as the structural complexity and the different cell types involved, make it difficult to fully simulate all features in vitro. Different 3-D models, which mimic specific components or processes of the respiratory system, have been extensively used to test various novel anti-infective agents. For example, the protective role of nasal sprays in maintaining the barrier function of nasal epithelial cells has been shown in vitro using a commercially available 3-D nasal mucosal model (De Servi et al. 2017). The ability of a novel antimicrobial peptide, derived from frog skin, to affect the permeability of lung epithelium has been shown using primary bronchial epithelial cells differentiated in vitro, where the barrier integrity was measured with an electrode determining the transepithelial electrical resistance (TEER) (Chen et al. 2017). A 3-D co-culture system consisting of the human airway epithelial cell line BEAS-2B, immature dendritic cells (DCs) derived from human circulating CD14+ monocytes and human lung fibroblast cell line MRC-5, was recently used to discriminate skin sensitizers from respiratory sensitizers. In this model, OX40L upregulation in DCs was preferentially induced by respiratory sensitizers and was found to be associated with the initiation of the T helper type 2 (Th2) cell response leading to respiratory allergy (Mizoguchi et al. 2017). Similarly, a 3-D lung model, including an epithelial cell line in combination with a human lung fibroblast cell line, was used to test the effect of pooled immunoglobulin (IVIG) in preventing S. aureus toxins mediated tissue damage. In addition, live imaging analysis of GFP-expressing epithelial cells within these tissue cultures confirmed the protective effect of IVIG (Mairpady Shambat et al. 2015).
Ex vivo studies of lung-related samples are proving useful in anti-infective research as well. For example, the benefit of the vaccine adjuvant CpG to promote T follicular helper cells, which are important in eliciting a robust anti-influenza antibody response, has been shown in nasopharynx-associated lymphoid tissues (NALT) derived from healthy children and adults (Aljurayyan et al. 2016). Furthermore, analysis of Pseudomonas aeruginosa biofilm aggregates in sputum from cystic fibrosis patients, treated with low concentrations of nitric oxide (NO), showed an enhanced efficacy of antibiotics in treating chronic infection (Howlin et al. 2017). Although beyond the scope of this review, it should be mentioned that numerous in vitro and ex vivo 3-D models are widely being used to study infection of bacteria as well as viruses and fungi and could be important models to test novel anti-infective treatments in the future. In brief, ex vivo models such as human nasal respiratory mucosa have been used in the context of viral infection for studying herpes simplex virus type 1 invasion (Glorieux et al. 2011; Wang et al. 2012). Besides this, infections with Mycobacterium tuberculosis, Aspergillus fumigatus, and Burkholderia cenocepacia have also been extensively studied (Chandorkar et al. 2017; Fonseca et al. 2017; Van den Driessche et al. 2017). A future potential application of these models can be to test the efficacy of nasal nanovaccines, which represent a promising novel anti-infective (Bernocchi et al. 2017). In conclusion, 3-D organotypic human lung cell culture models can provide important information on the efficacy of novel antimicrobials.
2.4 Brain Models
Due to the large anatomical differences between the human brain and other mammals, there is a great need to establish experimental human brain models. However, since the brain contains a dense network of various cell types such as neurons, glial cells, and astrocytes, with complex interaction patterns, it is very challenging to recapitulate this system in vitro. Over the past years, a lot of effort has been made to this cause, e.g., in trying to develop neurospheroids and cerebral organoids (Jorfi et al. 2017). Many studies utilized 3-D human brain models to study the pathogenesis and identify potential therapeutic targets of infectious agents causing damage to the brain. A major advantage of using these models is that they allow for the high-throughput screening of potential anti-infectives, a very important feature in the response to the fast emergence of Zika virus. A pool of existing drugs has been tested for efficacy against Zika and Japanese encephalitis virus infection in in vitro models including fetal-like organoids, human fetal-derived neuronal stem cells, human cortical organoids, and human pluripotent stem cell-derived cortical neural progenitor cells (hNPCs) (Xu et al. 2016; Bullard-Feibelman et al. 2017; Li et al. 2017; Wang et al. 2017; Zhou et al. 2017). Here, it should be noted that the in vitro efficacy of antivirals has been confirmed by additional in vivo experiments in these studies. However, in the case of human-specific pathogens such as varicella zoster virus (VZV), the use of 3-D human brain models is even more critical. Human neuronal systems, derived from embryonic and neural stem cells, have been established to study the various stages of VZV infection and might function as a tool to investigate novel antiviral targets (Markus et al. 2015; Kurapati et al. 2017). Another study aimed to mimic the human blood–brain barrier (BBB) by culturing a human brain microvascular endothelial cell line (HBMEC) in a rotating wall vessel (RWV) bioreactor. This model can be used to study the invasion of pathogens into the central nervous system (Bramley et al. 2017). This system may enable the screening of novel or existing antivirals for their protective effect. Collectively, human brain models are extensively being used as a high-throughput screening tool for anti-infective research, in particular for antivirals. However, data from currently used 3-D models still require additional studies in animal models, due to the very complex characteristics of the human brain.
3 Part 2—in-Depth Overview of Skin Models for Anti-infective Research
The skin is the largest human organ and functions as a barrier against physical injury, infections, and environmental cues (e.g., UV light) and contributes to body temperature regulation (Proksch et al. 2008; Olaniyi et al. 2017). The skin is composed of two major layers: the epidermis and the dermis. Keratinocytes are the main cellular component of the epidermis and its stage of differentiation subdivide the epidermis into four strata: stratum corneum, stratum granulosum, stratum spinosum, and stratum basale. The outermost layer, stratum corneum, is populated by terminally differentiated keratinocytes, while stratum basale is the inner layer of the epidermis, which is composed of undifferentiated keratinocytes. Although less abundant, Langerhans cells, melanocytes, and cytotoxic T cells can also be found in the epidermis. The dermis is mainly composed of collagen and elastin fibers and is rich in immune cells such as dendritic cells, T cells, macrophages, and mast cells. Fibroblasts, nerves, as well as blood and lymphatic vessels are also present in this layer. In addition, epidermal appendages, such as sweat and sebaceous glands as well as hair follicles, are embedded in the dermal layer. Hair follicles transverse the two compartments of the skin, representing a major route of penetration for everything coming into contact with the skin, including microbes.
Keratinocytes are the first players to act when the skin barrier is challenged. Therefore, 2-D monolayers of keratinocytes have been used to study the primary immune response induced by keratinocytes in multiple fields, ranging from cosmetics to injury and pathogen infection. Nevertheless, monolayers of keratinocytes do not resemble the stratification characteristic of the epidermis. In order to overcome this limitation, reconstructed human epidermis (RHE) models can be built by culturing keratinocytes at the air–liquid interface (Netzlaff et al. 2005).
However, a complete model of the skin is needed to study host–pathogen interactions and anti-infective interventions, including their toxicity, efficacy, and induced immune response. Since animal experimentation is time-consuming, expensive, and its predictive value is limited when applied to humans, the ideal model should reproduce both histology and architecture of the human skin. Full-thickness skin models, such as human skin equivalents and human skin explants, fulfill these requirements.
3.1 Skin Equivalent Versus Skin Explants
Many studies describe the development of skin equivalent models where the different layers of the skin are rebuilt using primary fibroblasts within a matrix of collagen followed by seeding with keratinocytes. Further differentiation of keratinocytes at air–liquid interface results in stratified layers, closely resembling the human skin. De-vitalized de-epidermalized dermis (DED), obtained by discarding both epidermis and adipose tissue from human skin, can also be used as a more physiological support to grow keratinocytes (Lamb and Ambler 2014; Ventress et al. 2016). In addition, either primary or immortalized keratinocytes can be used to construct the epidermal layer, although the widely used HaCaT cell line does not reach the terminal differentiation stage (Boelsma et al. 1999; Schoop et al. 1999). Human skin explants, commonly collected from cosmetic surgery procedures, represent the most complete skin model.
When selecting a model to evaluate the efficacy or toxicity of an anti-infective compound, it should be taken into account that each of the full-thickness human skin models, which include skin equivalents and skin explants, presents advantages and disadvantages (Table 1).
In order to compensate for the lack of immune cells in the skin equivalent models, Langerhans cells (Dezutter-Dambuyant et al. 2006; Laubach et al. 2011; Ouwehand et al. 2011), dendritic cells (Bechetoille et al. 2007) and macrophages (Bechetoille et al. 2011) have been successfully incorporated into skin equivalent models. Nevertheless, the fact that skin equivalent is built using cells from multiple donors makes the model genetically heterogeneous. On the other hand, the possibility of modulating the cellular components of the skin equivalents allows for studying the role of each cell type. Another advantage of skin equivalent models is the possibility of employing genetic engineering to unveil the function of a specific factor in the context of an infection and immune response, by inducing or depleting its expression in a particular cell type before its introduction into the skin matrix.
Skin explants preserve the histology and the complexity of the skin, being composed of several different resident cells (including immune cells) and appendages. This feature enables the study of the cutaneous local immune response raised against a pathogen or triggered by an anti-infective immune intervention at this site. However, skin explants have a short-term viability and limited availability. Additionally, the explant is not connected to circulation and consequently lacks the ability to recruit immune cells from the blood to the explant, such as neutrophils that are important for microbial clearance. Further, the inter-donor variability of the human skin explants makes interpretation of results more complex, thus requiring a greater number of samples from different donors to be analyzed in order to guarantee validity of the results. On the other hand, this feature is a key to better understand the host-inherent variability leading to differential individual responses, opening the door to personalized medicine.
3.2 Use of Human Skin Models in Anti-infective Research
Similar to the respiratory system, the skin has direct contact with the outside environment and therefore acts as a physical and chemical barrier for external threats. The skin is a common initial site of infection, where the first encounter of the immune system with a pathogen often occurs. Consequently, skin models are important for studying infections and anti-infective agents.
Although human skin models have been used to study microbial infection and pathogenesis by a number of authors, there are only a few studies focusing on anti-infective research. Since mouse skin does not resemble human skin in either histology or immunology (Pasparakis et al. 2014), efforts have been made to use human skin equivalent and human skin explant models to test safety and efficacy of antibacterial and antiviral compounds.
Poxviruses have a tropism for skin and mucosal tissues. Thereby, an organotypic culture system, built with human primary keratinocytes derived from neonatal foreskin, was used to prove the inhibitory effect of some acyclic nucleoside phosphonates on the viral cytopathic effect (Snoeck et al. 2002). The same model was also used to evaluate the antiviral activity of the compounds acyclovir, penciclovir, brivudin, foscarnet, and cidofovir against wild-type and mutant alphaherpesviruses HSV type 1 and type 2, and VZV (Andrei et al. 2005). In both studies, the antivirals were added to the medium to mimic systemic delivery of the compounds.
Due to the fact that Staphylococcus aureus is the major cause of bacterial skin and soft tissue infections in humans (McCaig et al. 2006) and the widespread emergence of antibiotic-resistant strains, recent studies have tested strategies to block this pathogen at the initial site of the infection. A study performed in a human full-thickness skin equivalent, constructed by keratinocytes and fibroblasts seeded on top of a DED scaffold, showed that a tetraspanin-derived peptide can halve the number of S. aureus adherent to keratinocytes as compared to a scrambled control peptide, without impairing keratinocyte viability (Ventress et al. 2016). Although it is still not clear if S. aureus infection is the cause or the consequence of atopic dermatitis (AD), the low expression of cutaneous antimicrobial peptides (AMPs) in AD patients is thought to contribute to their increased susceptibility to infections (Ong et al. 2002). Therefore, different strategies have been employed to increase AMP levels to prevent S. aureus infection. A study published this year used heat-killed Lactobacillus johnsonii NCC 533, a non-replicating probiotic, as a topical treatment to increase the expression of AMP. This treatment reduced the adhesion of S. aureus to the RHE model by up to 74% (Rosignoli et al. 2018). Synthetic antimicrobial peptides derived from the natural AMP LL-37, namely P60.4, peptide 10, and SAAP-148, have been tested for their efficiency against multi-drug resistance strains on RHE models (Haisma et al. 2016), human skin equivalents (Haisma et al. 2014), and split-thickness human skin explants (de Breij et al. 2012), respectively. All peptides proved to have antimicrobial and antibiofilm properties against methicillin-resistant S. aureus (MRSA), and SAAP-148 was also effective against multi-drug resistant (MDR) Acinetobacter baumannii (de Breij et al. 2012). LL-37 formulated in a nano-sized liposome has also been proved to be effective against herpes simplex virus 1 (HSV-1) infection in RHE models (Ron-Doitch et al. 2016). Interestingly, the cytotoxicity induced by the formulation was greater in the monolayer of HaCaT cells than in the RHE model, which could be due to the presence of stratum corneum, acting as a protective layer.
Due to the increasing interest in this field of research, we focused on possible future directions with the skin model based on current trends in anti-infective research. The skin, in particular the dermis, can serve as an efficient delivery system of vaccines due to the easy access and presence of many immune cells which can elicit a strong response to vaccine antigens. Intradermal vaccination has already shown to alter Langerhans cell distribution within human skin explants (Pearton et al. 2010), as well as to induce DC maturation (de Gruijl et al. 2006). Similarly, vaccination by microneedles can affect dendritic cell migration (Pearton et al. 2013). Since more evidence is emerging that the gut microbiome can influence immune responses, and hence vaccine efficacy (Littman 2017; Lynn and Pulendran 2017), one could argue that this applies to the skin microbiome as well. This hypothesis could be evaluated using the human skin model, whereby investigating the effect of known skin commensals on the cutaneous immune system. The application of topical compounds, e.g., in treating psoriasis (Kusuba et al. 2017) or other inflammatory diseases, and its effect on susceptibility to infections can be studied in depth with skin models, especially skin explants since the skin surface closely resembles the in vivo situation. In addition, novel anti-infective approaches, such as nano-sponges that absorb secreted bacterial toxins (Hu et al. 2013; Escajadillo et al. 2017) or other innovative antimicrobial delivery systems (Hasan et al. 2017), can be tested for their efficacy with the skin model. Similarly, our group recently showed the protective effect of antibodies against S. aureus toxins within the context of the human skin explant model (Olaniyi et al. 2018).
Despite their many advantages, most of the currently used skin models have some limitations, as mentioned before. One of the most prominent limitations is the lack of a systemic flow, thereby leaving out essential immune processes which are key in anti-infective research, i.e., neutrophil recruitment, complement activation, and T cell migration (Pozzi et al. 2015). One way to overcome this deficit is the introduction of the organ on a chip technology, where microfluidics are incorporated in a culture model (Ahadian et al. 2017). Many skin-on-chip models have been established by using different cell types, as reviewed in (van den Broek et al. 2017), and may provide novel insights on therapeutic approaches in the future (Wufuer et al. 2016). Another technique to mimic skin perfusion is an isolated perfused human skin flap model, which has been developed to test drug delivery to the skin. In brief, a human skin explant was linked to an automated perfusion system via one vessel, thereby inducing a controlled, continuous flow. An infrared (IR) camera was used to constantly monitor the skin surface temperature, and thus the degree of skin perfusion (Ternullo et al. 2017). In conclusion, skin models provide a new platform to study the effectiveness of various novel anti-infective compounds, as summarized in Fig. 1.

Proposed future applications of human skin models in anti-infective research. a Human skin models can be used to study mechanism of action of intradermal vaccination and its efficacy against dermal pathogens and its interaction with commensal bacteria. b Novel topical agents may be tested for efficacy and potential side effects in a human skin model, either skin equivalent or skin explant. c Incorporation of microfluidics in the human skin model would mimic the systemic blood flow in the culture model enabling critical immune processes, such as neutrophil recruitment or tissue distribution of antibodies

Schematic overview of the protocol for the culture of human skin equivalents and human skin explants. a Human skin equivalent can be cultured in two steps, starting with the dermal component consisting of collagen and fibroblasts. Other immune cells, such as macrophages, can be included at this step. To allow for the fibroblasts to contract, keratinocytes can be seeded after 7 days. Thereafter, differentiation of keratinocytes at air–liquid interface will require 7–14 days, depending on the origin of the keratinocytes. b When handling fresh human skin explants four different critical steps are involved. The first is the removal of the adipose tissue. To create a superficial wound for infection protocols, the stratum corneum can be removed by repeated tape-stripping followed by the preparation of punches with disposable surgical biopsy punches. The biopsy punches can be washed and cultured at air–liquid interface
3.3 Conclusion
Taken together, over the last decade, human 3-D organotypic models have acquired increasing importance and great efforts have been made to improve the culture, handling, and reproducibility of these in vitro models. In contrast, a trend may be observed in which the use of animal models is becoming obsolete. One explanation could be the overall consensus that the use of animal models has to be reduced from an ethical point of view. Secondly, the occurrence of failed studies on novel anti-infectives in the clinical phase underlines the complexity of predicting human response using animal models. Despite the remaining limitations, the use of human organotypic models can be a valuable tool to facilitate the transition from preclinical to clinical studies.
3.4 Protocol: Culture of Human Skin Equivalents and Skin Explants for Use in Anti-infective Research
Figure 2 summarizes the procedures to culture full-thickness human skin models for use in anti-infective research.
3.4.1 Procedure for the Preparation of Human Skin Equivalents
-
1.
Materials
-
1.1
Cells
-
Primary human dermal fibroblasts
-
Human keratinocyte cell line/primary human keratinocytes
-
-
1.2
Reagents
-
Collagen type I from bovine hides (3 mg/ml)
-
Sterile phosphate buffered saline (PBS)
-
Trypsin-EDTA (0.05%)
-
2N NaOH
-
HBSS 10x with phenol-red
-
Fibroblast culture medium:
-
DMEM High glucose
-
100 U/mL of penicillin and 100 μg/mL of streptomycin
-
10% fetal bovine serum (FBS) heat inactivated at 56 °C for 30 min
-
-
Keratinocyte culture medium:
-
DMEM high glucose
-
100 U/mL of penicillin and 100 μg/mL of streptomycin
-
-
FAD medium:
-
DMEM/F12 (1:1)
-
0.4 ug/mL hydrocortisone
-
50 ug/mL L-ascorbic acid
-
1 × 10−10 M Cholera toxin
-
1% antibiotic-antimycotic
-
10% FBS heat inactivated
-
-
FAD airlift interface medium (ALI)
-
FAD medium
-
0.5 mM Na-pyruvate
-
1.6 mM CaCl2
-
20 ng/mL TGF-alpha
-
100 ng/mL GM-CSF
-
All media must be filtered and stored at 4 °C, no longer than 2 weeks.
-
-
1.3
Equipment
-
Cell culture incubator (37 °C, 5% CO2)
-
Cell culture incubator (37 °C, 0% CO2)
-
Laminar flow class II cabinet
-
Hemocytometer
-
Pipet-aid
-
Sterile disposable pipettes
-
Single pipette with sterile tips
-
Polypropylene tubes, 15 mL and 50 mL
-
Glass Pasteur pipettes
-
12 transwell inserts preloaded in 12-well plates; pore size: 0.4 µm, sterile (Transwell®)
-
Sterile filter bottles (0.2 µm)
-
Ice buckets
-
Water bath
-
Centrifuge
-
-
1.1
-
2.
Methods
-
2.1
Dermal component
Before starting the protocol:
-
Place 10xHBSS, collagen and 2N NaOH on ice and disposable pipettes in the fridge;
-
Set an incubator at 37 °C, 0% CO2;
-
Calculate the amount of collagen mix needed to fill each insert with 500 μL. Considering that collagen mix is composed of collagen: 10xHBSS: fibroblast solution in the ratio 8:1:1, calculate the required amount of each of the components.
The following procedure must be performed under a laminar flow cabinet:
-
1.
Harvest fibroblasts and count the number of cells using a hemocytometer
-
2.
Suspend fibroblasts in FBS to a concentration of 1 × 106 cells/mL
-
3.
Keep it at room temperature (RT)
From this step forward work always on ice:
-
4.
Slowly add one part of chilled 10xHBSS to eight parts of chilled collagen in a 50 mL tube, with gentle swirling
-
5.
Adjust the pH of the solution to pH 7.4 using 2N NaOH, dropwise until the color changes to orange/red
It usually requires 50 μL of 2N NaOH per 10 mL of collagen mix
-
6.
Add one part of the fibroblasts solution to the collagen mix and homogenize the gel solution carefully, using a disposable pipette.
-
7.
Add 500 μL of the complete collagen mix onto the filter inserts
Prevent bubbles by reverse pipetting
-
8.
To allow the gel to solidify, place the 12-well plates for 1 h at 37 °C, 0% CO2
-
9.
Add 2 mL FAD medium (1.5 mL outside, 0.5 mL inside the insert) and incubate for 7 days at 37 °C, 5% CO2. Replace the medium every 2-3 days.
-
-
2.2
Epidermal component
-
10.
Harvest keratinocytes and count the number of cells using a hemocytometer
-
11.
Prepare keratinocytes at 5 × 105 cell/ml in keratinocyte culture medium
-
12.
Remove the medium inside the inserts
-
13.
Add 20 μL of keratinocyte solution to the dermal equivalent
-
14.
Incubate for 2 h at 37 °C, 5% CO2
-
15.
Add 500 μL of FAD medium inside the inserts
-
16.
Incubate overnight
-
17.
Remove all medium from the wells
-
18.
Add 500 μL FAD ALI medium outside the insert, to allow culture at air–liquid interface
-
19.
Incubate up to 7–14 days (dependent on differentiation status) and change FAD ALI medium every 2–3 days.
-
10.
-
2.1
3.4.2 Procedure for the Preparation of Human Skin Explants (Olaniyi et al. 2018)
The outermost stratum of the epidermis, stratum corneum, is the major responsibility for maintaining the physical barrier function of the skin (Baroni et al. 2012). Therefore, this barrier has to be breached so that a pathogen can invade the tissue. Here, we described the use of the tape-stripping strategy as we consider that this method guarantees reproducibility between experiments.
-
1.
Materials
-
1.1
Reagents
-
Fresh human skin explants
-
Sterile phosphate buffered saline (PBS)
-
Culture medium:
-
Advanced DMEM supplemented with 4 mM L-glutamine
-
All media should be filtered and stored at 4 °C, no longer than 2 weeks.
-
-
1.2
Equipment
-
Cell culture incubator (37 °C, 5% CO2)
-
Laminar flow class II cabinet
-
Pipet-aid
-
Sterile disposable pipettes
-
Single pipette with sterile tips
-
Glass Pasteur pipettes
-
12 transwell inserts preloaded in 12-well plates; pore size: 0.4 µm, sterile (Transwell®)
-
Disposable surgical biopsy punches
-
Disposable scalpels
-
Sterile forceps
-
Sterile scissors
-
Sterile filter bottles (0.2 µm)
-
Water bath
-
Adhesive tape
-
Dissection board
-
Disinfected pins
-
-
1.1
-
2.
Methods
-
2.1
Skin processing
Process the tissue immediately after its delivery.
The following procedure must be performed under a laminar flow cabinet:
-
1.
Remove the excess of adipose tissue, using disposable scalpels or sterile scissors
-
2.
Pin the skin in the dissection board
-
3.
Strip the stratum corneum 30 times with an adhesive tape
Make sure that the surface of the skin is dry before starting
-
4.
Take 8 mm (in diameter) punch biopsies using disposable surgical biopsy punches
-
5.
Place the punches inside a six-well plate, previously filled in with culture medium to prevent them to dry
-
6.
Remove the remaining adipose tissue from the punches using a scalpel
-
7.
Add 1 mL of culture medium in the 12-well plate
-
8.
Place the skin punches in the inserts using sterile forceps
-
9.
Culture the explants at air–liquid interface, at 37 °C, 5% CO2.
-
1.
-
2.1
Change history
19 July 2019
∎∎∎
References
Abner SR, Guenthner PC, Guarner J, Hancock KA, Cummins JE Jr, Fink A, Gilmore GT, Staley C, Ward A, Ali O, Binderow S, Cohen S, Grohskopf LA, Paxton L, Hart CE, Dezzutti CS (2005) A human colorectal explant culture to evaluate topical microbicides for the prevention of HIV infection. J Infect Dis 192(9):1545–1556
Ahadian S, Civitarese R, Bannerman D, Mohammadi MH, Lu R, Wang E, Davenport-Huyer L, Lai B, Zhang B, Zhao Y, Mandla S, Korolj A, Radisic M (2017) Organ-on-a-chip platforms: a convergence of advanced materials, cells, and microscale technologies. Adv Health Mater
Alam S, Bromberg-White J, McLaughlin-Drubin M, Sen E, Bodily JM, Meyers C (2005) Activity and therapeutic potential of ORI-1001 antisense oligonucleotide on human papillomavirus replication utilizing a model of dysplastic human epithelium. Anticancer Res 25(2A):765–777
Aljurayyan AN, Sharma R, Upile N, Beer H, Vaughan C, Xie C, Achar P, Ahmed MS, McNamara PS, Gordon SB, Zhang Q (2016) A critical role of T follicular helper cells in human mucosal anti-influenza response that can be enhanced by immunological adjuvant CpG-DNA. Antiviral Res 132:122–130
Andrei G, Lisco A, Vanpouille C, Introini A, Balestra E, van den Oord J, Cihlar T, Perno CF, Snoeck R, Margolis L, Balzarini J (2011) Topical tenofovir, a microbicide effective against HIV, inhibits herpes simplex virus-2 replication. Cell Host Microbe 10(4):379–389
Andrei G, van den Oord J, Fiten P, Opdenakker G, De Wolf-Peeters C, De Clercq E, Snoeck R (2005) Organotypic epithelial raft cultures as a model for evaluating compounds against alphaherpesviruses. Antimicrob Agents Chemother 49(11):4671–4680
Baeten JM, Strick LB, Lucchetti A, Whittington WL, Sanchez J, Coombs RW, Magaret A, Wald A, Corey L, Celum C (2008) Herpes simplex virus (HSV)-suppressive therapy decreases plasma and genital HIV-1 levels in HSV-2/HIV-1 coinfected women: a randomized, placebo-controlled, cross-over trial. J Infect Dis 198(12):1804–1808
Balzarini J, Andrei G, Balestra E, Huskens D, Vanpouille C, Introini A, Zicari S, Liekens S, Snoeck R, Holy A, Perno CF, Margolis L, Schols D (2013) A multi-targeted drug candidate with dual anti-HIV and anti-HSV activity. PLoS Pathog 9(7):e1003456
Baroni A, Buommino E, De Gregorio V, Ruocco E, Ruocco V, Wolf R (2012) Structure and function of the epidermis related to barrier properties. Clin Dermatol 30(3):257–262
Bechetoille N, Dezutter-Dambuyant C, Damour O, Andre V, Orly I, Perrier E (2007) Effects of solar ultraviolet radiation on engineered human skin equivalent containing both Langerhans cells and dermal dendritic cells. Tissue Eng 13(11):2667–2679
Bechetoille N, Vachon H, Gaydon A, Boher A, Fontaine T, Schaeffer E, Decossas M, Andre-Frei V, Mueller CG (2011) A new organotypic model containing dermal-type macrophages. Exp Dermatol 20(12):1035–1037
Bernocchi B, Carpentier R, Betbeder D (2017) Nasal nanovaccines. Int J Pharm
Boelsma E, Verhoeven MC, Ponec M (1999) Reconstruction of a human skin equivalent using a spontaneously transformed keratinocyte cell line (HaCaT). J Invest Dermatol 112(4):489–498
Bramley JC, Drummond CG, Lennemann NJ, Good CA, Kim KS, Coyne CB (2017) A three-dimensional cell culture system to model RNA virus infections at the blood-brain barrier. mSphere 2(3)
Buffa V, Stieh D, Mamhood N, Hu Q, Fletcher P, Shattock RJ (2009) Cyanovirin-N potently inhibits human immunodeficiency virus type 1 infection in cellular and cervical explant models. J Gen Virol 90(Pt 1):234–243
Bullard-Feibelman KM, Govero J, Zhu Z, Salazar V, Veselinovic M, Diamond MS, Geiss BJ (2017) The FDA-approved drug sofosbuvir inhibits Zika virus infection. Antiviral Res 137:134–140
Buve A (2010) Can we reduce the spread of HIV infection by suppressing herpes simplex virus type 2 infection? F1000 Med Rep 2
Cates W Jr (2010) After CAPRISA 004: time to re-evaluate the HIV lexicon. Lancet 376(9740):495–496
Chandorkar P, Posch W, Zaderer V, Blatzer M, Steger M, Ammann CG, Binder U, Hermann M, Hortnagl P, Lass-Florl C, Wilflingseder D (2017) Fast-track development of an in vitro 3D lung/immune cell model to study Aspergillus infections. Sci Rep 7(1):11644
Chen C, Mangoni ML, Di YP (2017) In vivo therapeutic efficacy of frog skin-derived peptides against Pseudomonas aeruginosa-induced pulmonary infection. Sci Rep 7(1):8548
Cummins JE Jr, Guarner J, Flowers L, Guenthner PC, Bartlett J, Morken T, Grohskopf LA, Paxton L, Dezzutti CS (2007) Preclinical testing of candidate topical microbicides for anti-human immunodeficiency virus type 1 activity and tissue toxicity in a human cervical explant culture. Antimicrob Agents Chemother 51(5):1770–1779
de Breij A, Haisma EM, Rietveld M, El Ghalbzouri A, van den Broek PJ, Dijkshoorn L, Nibbering PH (2012) Three-dimensional human skin equivalent as a tool to study Acinetobacter baumannii colonization. Antimicrob Agents Chemother 56(5):2459–2464
de Gruijl TD, Ophorst OJ, Goudsmit J, Verhaagh S, Lougheed SM, Radosevic K, Havenga MJ, Scheper RJ (2006) Intradermal delivery of adenoviral type-35 vectors leads to high efficiency transduction of mature, CD8 + T cell-stimulating skin-emigrated dendritic cells. J Immunol 177(4):2208–2215
De Servi B, Ranzini F, Pique N (2017) Protective barrier properties of Rhinosectan(R) spray (containing xyloglucan) on an organotypic 3D airway tissue model (MucilAir): results of an in vitro study. Allergy Asthma Clin Immunol 13:37
Delvenne P, Hubert P, Jacobs N, Giannini SL, Havard L, Renard I, Saboulard D, Boniver J (2001) The organotypic culture of HPV-transformed keratinocytes: an effective in vitro model for the development of new immunotherapeutic approaches for mucosal (pre)neoplastic lesions. Vaccine 19(17–19):2557–2564
Dezutter-Dambuyant C, Black A, Bechetoille N, Bouez C, Marechal S, Auxenfans C, Cenizo V, Pascal P, Perrier E, Damour O (2006) Evolutive skin reconstructions: from the dermal collagen-glycosaminoglycan-chitosane substrate to an immunocompetent reconstructed skin. Biomed Mater Eng 16(4 Suppl):S85–S94
Dezzutti CS, Russo J, Wang L, Abebe KZ, Li J, Friend DR, McGowan IM, Rohan LC (2014) Development of HIV-1 rectal-specific microbicides and colonic tissue evaluation. PLoS ONE 9(7):e102585
Dezzutti CS, Shetler C, Mahalingam A, Ugaonkar SR, Gwozdz G, Buckheit KW, Buckheit RW Jr (2012) Safety and efficacy of tenofovir/IQP-0528 combination gels—a dual compartment microbicide for HIV-1 prevention. Antiviral Res 96(2):221–225
Dezzutti CS, Yandura S, Wang L, Moncla B, Teeple EA, Devlin B, Nuttall J, Brown ER, Rohan LC (2015) Pharmacodynamic activity of Dapivirine and Maraviroc single entity and combination topical gels for HIV-1 prevention. Pharm Res 32(11):3768–3781
Escajadillo T, Olson J, Luk BT, Zhang L, Nizet V (2017) A red blood cell membrane-camouflaged nanoparticle counteracts streptolysin O-Mediated virulence phenotypes of invasive group a streptococcus. Front Pharmacol 8:477
Ferrini S, Cambiaggi A, Sforzini S, Marciano S, Canevari S, Mezzanzanica D, Colnaghi MI, Grossi CE, Moretta L (1993) Targeting of T lymphocytes against EGF-receptor + tumor cells by bispecific monoclonal antibodies: requirement of CD3 molecule cross-linking for T-cell activation. Int J Cancer 55(6):931–937
Fletcher P, Kiselyeva Y, Wallace G, Romano J, Griffin G, Margolis L, Shattock R (2005) The nonnucleoside reverse transcriptase inhibitor UC-781 inhibits human immunodeficiency virus type 1 infection of human cervical tissue and dissemination by migratory cells. J Virol 79(17):11179–11186
Fletcher PS, Elliott J, Grivel JC, Margolis L, Anton P, McGowan I, Shattock RJ (2006a) Ex vivo culture of human colorectal tissue for the evaluation of candidate microbicides. AIDS 20(9):1237–1245
Fletcher PS, Wallace GS, Mesquita PM, Shattock RJ (2006b) Candidate polyanion microbicides inhibit HIV-1 infection and dissemination pathways in human cervical explants. Retrovirology 3:46
Fonseca KL, Rodrigues PNS, Olsson IAS, Saraiva M (2017) Experimental study of tuberculosis: from animal models to complex cell systems and organoids. PLoS Pathog 13(8):e1006421
Global Burden of Disease Study C (2015) Global, regional, and national incidence, prevalence, and years lived with disability for 301 acute and chronic diseases and injuries in 188 countries, 1990–2013: a systematic analysis for the Global Burden of Disease Study 2013. Lancet 386(9995):743–800
Glorieux S, Bachert C, Favoreel HW, Vandekerckhove AP, Steukers L, Rekecki A, Van den Broeck W, Goossens J, Croubels S, Clayton RF, Nauwynck HJ (2011) Herpes simplex virus type 1 penetrates the basement membrane in human nasal respiratory mucosa. PLoS ONE 6(7):e22160
Greenhead P, Hayes P, Watts PS, Laing KG, Griffin GE, Shattock RJ (2000) Parameters of human immunodeficiency virus infection of human cervical tissue and inhibition by vaginal virucides. J Virol 74(12):5577–5586
Grivel JC, Margolis L (2009) Use of human tissue explants to study human infectious agents. Nat Protoc 4(2):256–269
Grulich AE, Zablotska I (2010) Commentary: probability of HIV transmission through anal intercourse. Int J Epidemiol 39(4):1064–1065
Haisma EM, de Breij A, Chan H, van Dissel JT, Drijfhout JW, Hiemstra PS, El Ghalbzouri A, Nibbering PH (2014) LL-37-derived peptides eradicate multidrug-resistant Staphylococcus aureus from thermally wounded human skin equivalents. Antimicrob Agents Chemother 58(8):4411–4419
Haisma EM, Goblyos A, Ravensbergen B, Adriaans AE, Cordfunke RA, Schrumpf J, Limpens RW, Schimmel KJ, den Hartigh J, Hiemstra PS, Drijfhout JW, El Ghalbzouri A, Nibbering PH (2016) Antimicrobial Peptide P60.4Ac-Containing Creams and Gel for Eradication of Methicillin-Resistant Staphylococcus aureus from Cultured Skin and Airway Epithelial Surfaces. Antimicrob Agents Chemother 60(7):4063–4072
Hasan S, Thomas N, Thierry B, Prestidge CA (2017) Controlled and Localized Nitric Oxide Precursor Delivery from Chitosan Gels to Staphylococcus aureus Biofilms. J Pharm Sci
Herrera C, Cranage M, McGowan I, Anton P, Shattock RJ (2009) Reverse transcriptase inhibitors as potential colorectal microbicides. Antimicrob Agents Chemother 53(5):1797–1807
Herrera C, Cranage M, McGowan I, Anton P, Shattock RJ (2011) Colorectal microbicide design: triple combinations of reverse transcriptase inhibitors are optimal against HIV-1 in tissue explants. AIDS 25(16):1971–1979
Howlin RP, Cathie K, Hall-Stoodley L, Cornelius V, Duignan C, Allan RN, Fernandez BO, Barraud N, Bruce KD, Jefferies J, Kelso M, Kjelleberg S, Rice SA, Rogers GB, Pink S, Smith C, Sukhtankar PS, Salib R, Legg J, Carroll M, Daniels T, Feelisch M, Stoodley P, Clarke SC, Connett G, Faust SN, Webb JS (2017) Low-Dose Nitric Oxide as targeted anti-biofilm adjunctive therapy to treat Chronic Pseudomonas aeruginosa Infection in Cystic Fibrosis. Mol Ther
Hu CM, Fang RH, Copp J, Luk BT, Zhang L (2013) A biomimetic nanosponge that absorbs pore-forming toxins. Nat Nanotechnol 8(5):336–340
Jacobs N, Moutschen MP, Franzen-Detrooz E, Boniver V, Boniver J, Delvenne P (1998) Organotypic culture of HPV-transformed keratinocytes: a model for testing lymphocyte infiltration of (pre)neoplastic lesions of the uterine cervix. Virchows Arch 432(4):323–330
Jorfi M, D’Avanzo C, Kim DY, Irimia D (2017) Three-dimensional models of the human brain development and diseases. Adv Health Mater
Kurapati S, Sadaoka T, Rajbhandari L, Jagdish B, Shukla P, Ali MA, Kim YJ, Lee G, Cohen JI, Venkatesan A (2017) Role of the JNK pathway in varicella-zoster virus lytic infection and reactivation. J Virol 91(17)
Kusuba N, Kitoh A, Dainichi T, Honda T, Otsuka A, Egawa G, Nakajima S, Miyachi Y, Kabashima K (2017) Inhibition of IL-17-committed T cells in a murine psoriasis model by a vitamin D analogue. J Allergy Clin Immunol
Lamb R, Ambler CA (2014) Modified methods for growing 3-D skin equivalents: an update. Methods Mol Biol 1195:171–182
Lancaster MA, Knoblich JA (2014) Organogenesis in a dish: modeling development and disease using organoid technologies. Science 345(6194):1247125
Laubach V, Zoller N, Rossberg M, Gorg K, Kippenberger S, Bereiter-Hahn J, Kaufmann R, Bernd A (2011) Integration of Langerhans-like cells into a human skin equivalent. Arch Dermatol Res 303(2):135–139
Li C, Deng YQ, Wang S, Ma F, Aliyari R, Huang XY, Zhang NN, Watanabe M, Dong HL, Liu P, Li XF, Ye Q, Tian M, Hong S, Fan J, Zhao H, Li L, Vishlaghi N, Buth JE, Au C, Liu Y, Lu N, Du P, Qin FX, Zhang B, Gong D, Dai X, Sun R, Novitch BG, Xu Z, Qin CF, Cheng G (2017) 25-Hydroxycholesterol protects host against Zika virus infection and its associated microcephaly in a mouse model. Immunity 46(3):446–456
Lin X, Paskaleva EE, Chang W, Shekhtman A, Canki M (2011) Inhibition of HIV-1 infection in ex vivo cervical tissue model of human vagina by palmitic acid; implications for a microbicide development. PLoS ONE 6(9):e24803
Lisco A, Vanpouille C, Tchesnokov EP, Grivel JC, Biancotto A, Brichacek B, Elliott J, Fromentin E, Shattock R, Anton P, Gorelick R, Balzarini J, McGuigan C, Derudas M, Gotte M, Schinazi RF, Margolis L (2008) Acyclovir is activated into a HIV-1 reverse transcriptase inhibitor in herpesvirus-infected human tissues. Cell Host Microbe 4(3):260–270
Littman DR (2017) Do the microbiota influence vaccines and protective immunity to pathogens? If so, is there potential for efficacious microbiota-based vaccines?” Cold Spring Harb Perspect Biol
Lu H, Zhao Q, Wallace G, Liu S, He Y, Shattock R, Neurath AR, Jiang BS (2006) Cellulose acetate 1,2-benzenedicarboxylate inhibits infection by cell-free and cell-associated primary HIV-1 isolates. AIDS Res Hum Retroviruses 22(5):411–418
Lynn DJ, Pulendran B (2017) The potential of the microbiota to influence vaccine responses. J Leukoc Biol
Mahalingam A, Simmons AP, Ugaonkar SR, Watson KM, Dezzutti CS, Rohan LC, Buckheit RW Jr, Kiser PF (2011) Vaginal microbicide gel for delivery of IQP-0528, a pyrimidinedione analog with a dual mechanism of action against HIV-1. Antimicrob Agents Chemother 55(4):1650–1660
Mairpady Shambat S, Chen P, Nguyen Hoang AT, Bergsten H, Vandenesch F, Siemens N, Lina G, Monk IR, Foster TJ, Arakere G, Svensson M, Norrby-Teglund A (2015) Modelling staphylococcal pneumonia in a human 3D lung tissue model system delineates toxin-mediated pathology. Dis Model Mech 8(11):1413–1425
Markus A, Lebenthal-Loinger I, Yang IH, Kinchington PR, Goldstein RS (2015) An in vitro model of latency and reactivation of varicella zoster virus in human stem cell-derived neurons. PLoS Pathog 11(6):e1004885
McCaig LF, McDonald LC, Mandal S, Jernigan DB (2006) Staphylococcus aureus-associated skin and soft tissue infections in ambulatory care. Emerg Infect Dis 12(11):1715–1723
Meyers C, Frattini MG, Hudson JB, Laimins LA (1992) Biosynthesis of human papillomavirus from a continuous cell line upon epithelial differentiation. Science 257(5072):971–973
Mizoguchi I, Ohashi M, Chiba Y, Hasegawa H, Xu M, Owaki T, Yoshimoto T (2017) Prediction of chemical respiratory and contact sensitizers by OX40L expression in dendritic cells using a novel 3D coculture system. Front Immunol 8:929
Nagot N, Ouedraogo A, Foulongne V, Konate I, Weiss HA, Vergne L, Defer MC, Djagbare D, Sanon A, Andonaba JB, Becquart P, Segondy M, Vallo R, Sawadogo A, Van de Perre P, Mayaud P, Group AS (2007) Reduction of HIV-1 RNA levels with therapy to suppress herpes simplex virus. N Engl J Med 356(8):790–799
Netzlaff F, Lehr CM, Wertz PW, Schaefer UF (2005) The human epidermis models EpiSkin, SkinEthic and EpiDerm: an evaluation of morphology and their suitability for testing phototoxicity, irritancy, corrosivity, and substance transport. Eur J Pharm Biopharm 60(2):167–178
Olaniyi RO, Pancotto L, Grimaldi L, Bagnoli F (2018) Deciphering the pathological role of Staphylococcal alpha-Toxin and Panton-Valentine Leukocidin using a novel ex vivo human skin model. Front Immunol 9:951
Olaniyi R, Pozzi C, Grimaldi L, Bagnoli F (2017) Staphylococcus aureus-associated skin and soft tissue infections: anatomical localization, epidemiology, therapy and potential prophylaxis. Curr Top Microbiol Immunol 409:199–227
Ong PY, Ohtake T, Brandt C, Strickland I, Boguniewicz M, Ganz T, Gallo RL, Leung DY (2002) Endogenous antimicrobial peptides and skin infections in atopic dermatitis. N Engl J Med 347(15):1151–1160
Ouwehand K, Spiekstra SW, Waaijman T, Scheper RJ, de Gruijl TD, Gibbs S (2011) Technical advance: langerhans cells derived from a human cell line in a full-thickness skin equivalent undergo allergen-induced maturation and migration. J Leukoc Biol 90(5):1027–1033
Pasparakis M, Haase I, Nestle FO (2014) Mechanisms regulating skin immunity and inflammation. Nat Rev Immunol 14(5):289–301
Pearton M, Kang SM, Song JM, Anstey AV, Ivory M, Compans RW, Birchall JC (2010) Changes in human Langerhans cells following intradermal injection of influenza virus-like particle vaccines. PLoS ONE 5(8):e12410
Pearton M, Pirri D, Kang SM, Compans RW, Birchall JC (2013) Host responses in human skin after conventional intradermal injection or microneedle administration of virus-like-particle influenza vaccine. Adv Healthc Mater 2(10):1401–1410
Popov L, Kovalski J, Grandi G, Bagnoli F, Amieva MR (2014) Three-dimensional human skin models to understand staphylococcus aureus skin colonization and infection. Front Immunol 5:41
Pozzi C, Lofano G, Mancini F, Soldaini E, Speziale P, De Gregorio E, Rappuoli R, Bertholet S, Grandi G, Bagnoli F (2015) Phagocyte subsets and lymphocyte clonal deletion behind ineffective immune response to Staphylococcus aureus. FEMS Microbiol Rev 39(5):750–763
Proksch E, Brandner JM, Jensen JM (2008) The skin: an indispensable barrier. Exp Dermatol 17(12):1063–1072
Renard I, Mezzanzanica D, Canevari S, Ferrini S, Boniver J, Delvenne P, Jacobs N (2002) Anti-CD3/anti-epidermal growth factor receptor-bispecific antibody retargeting of lymphocytes against human neoplastic keratinocytes in an autologous organotypic culture model. Am J Pathol 160(1):113–122
Rohan LC, Moncla BJ, Ayudhya RPKN, Cost M, Huang Y, Gai F, Billitto N, Lynam JD, Pryke K, Graebing P, Hopkins N, Rooney JF, Friend D, Dezzutti CS (2010) In vitro and ex vivo testing of tenofovir shows it is effective as an HIV-1 microbicide. PLoS ONE 5(2):e9310
Ron-Doitch S, Sawodny B, Kuhbacher A, David MM, Samanta A, Phopase J, Burger-Kentischer A, Griffith M, Golomb G, Rupp S (2016) Reduced cytotoxicity and enhanced bioactivity of cationic antimicrobial peptides liposomes in cell cultures and 3D epidermis model against HSV. J Control Release 229:163–171
Rosignoli C, Thibaut de Menonville S, Orfila D, Beal M, Bertino B, Aubert J, Mercenier A, Piwnica D (2018) A topical treatment containing heat-treated Lactobacillus johnsonii NCC 533 reduces Staphylococcus aureus adhesion and induces antimicrobial peptide expression in an in vitro reconstructed human epidermis model. Exp Dermatol
Schoop VM, Mirancea N, Fusenig NE (1999) Epidermal organization and differentiation of HaCaT keratinocytes in organotypic coculture with human dermal fibroblasts. J Invest Dermatol 112(3):343–353
Sen E, McLaughlin-Drubin M, Meyers C (2005) Efficacy of two commercial preparations of interferon-alpha on human papillomavirus replication. Anticancer Res 25(2A):1091–1100
Snoeck R, Holy A, Dewolf-Peeters C, Van Den Oord J, De Clercq E, Andrei G (2002) Antivaccinia activities of acyclic nucleoside phosphonate derivatives in epithelial cells and organotypic cultures. Antimicrob Agents Chemother 46(11):3356–3361
Ternullo S, de Weerd L, Flaten GE, Holsaeter AM, Skalko-Basnet N (2017) The isolated perfused human skin flap model: A missing link in skin penetration studies? Eur J Pharm Sci 96:334–341
van den Broek LJ, Bergers L, Reijnders CMA, Gibbs S (2017) Progress and future prospectives in skin-on-chip development with emphasis on the use of different cell types and technical challenges. Stem Cell Rev 13(3):418–429
Van den Driessche F, Vanhoutte B, Brackman G, Crabbe A, Rigole P, Vercruysse J, Verstraete G, Cappoen D, Vervaet C, Cos P, Coenye T (2017) Evaluation of combination therapy for Burkholderia cenocepacia lung infection in different in vitro and in vivo models. PLoS ONE 12(3):e0172723
Ventress JK, Partridge LJ, Read RC, Cozens D, MacNeil S, Monk PN (2016) Peptides from Tetraspanin CD9 Are Potent Inhibitors of Staphylococcus Aureus Adherence to Keratinocytes. PLoS ONE 11(7):e0160387
Wald A, Link K (2002) Risk of human immunodeficiency virus infection in herpes simplex virus type 2-seropositive persons: a meta-analysis. J Infect Dis 185(1):45–52
Wang S, Liu Y, Guo J, Wang P, Zhang L, Xiao G, Wang W (2017) Screening of FDA-Approved Drugs for Inhibitors against Japanese Encephalitis Virus Infection. J Virol
Wang X, Zhang N, Glorieux S, Holtappels G, Vaneechoutte M, Krysko O, Zhang L, Han D, Nauwynck HJ, Bachert C (2012) Herpes simplex virus type 1 infection facilitates invasion of Staphylococcus aureus into the nasal mucosa and nasal polyp tissue. PLoS ONE 7(6):e39875
Wufuer M, Lee G, Hur W, Jeon B, Kim BJ, Choi TH, Lee S (2016) Skin-on-a-chip model simulating inflammation, edema and drug-based treatment. Sci Rep 6:37471
Xu M, Lee EM, Wen Z, Cheng Y, Huang WK, Qian X, Tcw J, Kouznetsova J, Ogden SC, Hammack C, Jacob F, Nguyen HN, Itkin M, Hanna C, Shinn P, Allen C, Michael SG, Simeonov A, Huang W, Christian KM, Goate A, Brennand KJ, Huang R, Xia M, Ming GL, Zheng W, Song H, Tang H (2016) Identification of small-molecule inhibitors of Zika virus infection and induced neural cell death via a drug repurposing screen. Nat Med 22(10):1101–1107
Yin Y, Bijvelds M, Dang W, Xu L, van der Eijk AA, Knipping K, Tuysuz N, Dekkers JF, Wang Y, de Jonge J, Sprengers D, van der Laan LJ, Beekman JM, Ten Berge D, Metselaar HJ, de Jonge H, Koopmans MP, Peppelenbosch MP, Pan Q (2015) Modeling rotavirus infection and antiviral therapy using primary intestinal organoids. Antiviral Res 123:120–131
Zhang W, Hu M, Shi Y, Gong T, Dezzutti CS, Moncla B, Sarafianos SG, Parniak MA, Rohan LC (2015) Vaginal microbicide film combinations of two reverse transcriptase inhibitors, EFdA and CSIC, for the prevention of HIV-1 sexual transmission. Pharm Res 32(9):2960–2972
Zhou T, Tan L, Cederquist GY, Fan Y, Hartley BJ, Mukherjee S, Tomishima M, Brennand KJ, Zhang Q, Schwartz RE, Evans T, Studer L, Chen S (2017) High-content screening in hPSC-neural progenitors identifies drug candidates that inhibit Zika virus infection in fetal-like organoids and adult brain. Cell Stem Cell
Zomer-van Ommen DD, Pukin AV, Fu O, Quarles van Ufford LH, Janssens HM, Beekman JM, Pieters RJ (2016) Functional characterization of cholera toxin inhibitors using human intestinal organoids. J Med Chem 59(14):6968–6972
Zuckerman RA, Lucchetti A, Whittington WL, Sanchez J, Coombs RW, Zuniga R, Magaret AS, Wald A, Corey L, Celum C (2007) Herpes simplex virus (HSV) suppression with valacyclovir reduces rectal and blood plasma HIV-1 levels in HIV-1/HSV-2-seropositive men: a randomized, double-blind, placebo-controlled crossover trial. J Infect Dis 196(10):1500–1508
Zussman A, Lara L, Lara HH, Bentwich Z, Borkow G (2003) Blocking of cell-free and cell-associated HIV-1 transmission through human cervix organ culture with UC781. AIDS 17(5):653–661
Acknowledgements
We would like to acknowledge Reuben Olaniyi and Proinnsias Fox at GSK for critical reading of the manuscript.
Funding
This project is funded by the European Union’s Horizon 2020 research and innovation program under the Marie Skłodowska-Curie grant agreement No 675106.
Authorship
Fabio Bagnoli had the initial idea of the topic, content, and structure of the review article. Astrid Hendriks and Ana Rita Cruz defined the content and wrote the manuscript. Fabio Bagnoli, Andrea Manetti, and Elisabetta Soldaini reviewed and wrote the manuscript. All authors approved the manuscript before it was submitted.
Declaration of interest
Astrid Hendriks and Ana Rita Cruz are Ph.D. fellows and are enrolled in the Infection and Immunity Ph.D. program, part of the Graduate school of Life Sciences at the University of Utrecht. Fabio Bagnoli, Andrea Manetti, and Elisabetta Soldaini are employees of GSK group of companies. Fabio Bagnoli owns patents on S. aureus vaccine candidates and GSK stocks. The authors have no other relevant affiliations or financial involvement with any organization or entity with a financial interest in or financial conflict with the subject matter or materials discussed in the manuscript apart from those disclosed.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Open Access This chapter is licensed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license and indicate if changes were made.
The images or other third party material in this chapter are included in the chapter's Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the chapter's Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder.
Copyright information
© 2018 The Author(s)
About this chapter

Cite this chapter
Hendriks, A., Cruz, A.R., Soldaini, E., Manetti, A.G.O., Bagnoli, F. (2018). Human Organotypic Models for Anti-infective Research. In: Bagnoli, F., Rappuoli, R. (eds) Three Dimensional Human Organotypic Models for Biomedical Research. Current Topics in Microbiology and Immunology, vol 430. Springer, Cham. https://doi.org/10.1007/82_2018_130
Download citation
DOI: https://doi.org/10.1007/82_2018_130
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-030-62451-4
Online ISBN: 978-3-030-62452-1
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)