
Abstract
Reactivity is an important feature of every stable state to measure the short-term instability of such stable state in prey–predator models. Asymptotic stability is not a powerful tool to study the dynamics immediately after an equilibrium has disturbed. We are interested in studying the effect of disturbance made by harvesting effort to the stable state of a set of prey–predator models. We consider different scenarios of disturbance made by prey harvesting only, predator harvesting only and selective harvesting on both. We show that disturbance made by prey harvesting causes reactivity of stable state but not in a sustainable way (non-existence of maximum sustainable yield). Further, it is shown that disturbance made by predator harvesting causes reactivity of stable state but in a sustainable way. We study how disturbance made by different harvesting strategies to the asymptotically stable state in the prey–predator system where intraspecific competition rule applied to predator species affects transient dynamics. We find that the transient dynamics properties like reactivity and non-reactivity switch through disturbance made by harvesting but in a sustainable way (maximum sustainable yield exists). Further, we find that asymptotically stable state remains reactive following multiple disturbances made by selective harvesting on both but in a sustainable way (maximum sustainable total yield exists)










Similar content being viewed by others
References
K.E. Anderson, R.M. Nisbet, E. McCauley, Transient responses to spatial perturbations in advective systems. Bull. Math. Biol. 70(5), 1480–1502 (2008)
J.-F. Arnoldi, M. Loreau, B. Haegeman, Resilience, reactivity and variability: a mathematical comparison of ecological stability measures. J. Theor. Biol. 389, 47–59 (2016)
H. Caswell, Matrix Population Models: Construction, Analysis, and Interpretation, 2nd edn. (Sinauer Associates. Inc., Sunderland, MA, 2001)
H. Caswell, M.G. Neubert, Reactivity and transient dynamics of discrete-time ecological systems. J. Differ. Equ. Appl. 11(4–5), 295–310 (2005)
X.I.N. Chen, J.E. Cohen, Global stability, local stability and permanence in model food webs. J. Theor. Biol. 212(2), 223–235 (2001)
C.W. Clark, Mathematical bioeconomics, in Mathematical Problems in Biology. (Springer, 1974), pp. 29–45
J.A. Estes, D.O. Duggins, Sea otters and kelp forests in alaska: generality and variation in a community ecological paradigm. Ecol. Monogr. 65(1), 75–100 (1995)
B. Ghosh, T.K. Kar, Sustainable use of prey species in a prey–predator system: jointly determined ecological thresholds and economic trade-offs. Ecol. Model. 272, 49–58 (2014)
B. Ghosh, P. Paul, T.K. Kar, Extinction scenarios in exploited system: combined and selective harvesting approaches. Ecol. Complex. 19, 130–139 (2014)
M. Kot, Elements of Mathematical Ecology (Cambridge University Press, Cambridge, 2001)
T. Legović, S. Geček, Impact of maximum sustainable yield on independent populations. Ecol. Model. 221(17), 2108–2111 (2010)
T. Legović, J. Klanjšček, S. Geček, Maximum sustainable yield and species extinction in ecosystems. Ecol. Modell. 221(12), 1569–1574 (2010)
L. Mari, R. Casagrandi, A. Rinaldo, M. Gatto, A generalized definition of reactivity for ecological systems and the problem of transient species dynamics. Methods Ecol. Evolut. 8(11), 1574–1584 (2017)
M. Marvier, P. Kareiva, M.G. Neubert, Habitat destruction, fragmentation, and disturbance promote invasion by habitat generalists in a multispecies metapopulation. Risk Anal. Int. J. 24(4), 869–878 (2004)
H. Matsuda, P.A. Abrams, Maximal yields from multispecies fisheries systems: rules for systems with multiple trophic levels. Ecol. Appl. 16(1), 225–237 (2006)
H. Matsuda, P.A. Abrams, Is feedback control effective for ecosystem-based fisheries management? J. Theor. Biol. 339, 122–128 (2013)
R.M. May, G.R. Conway, M.P. Hassell, T.R.E. Southwood, Time delays, density-dependence and single-species oscillations. J. Anim. Ecol. 747–770 (1974)
R.M. May, J.R. Beddington, C.W. Clark, S.J. Holt, R.M. Laws, Management of multispecies fisheries. Science 205(4403), 267–277 (1979)
M.G. Neubert, H. Caswell, Alternatives to resilience for measuring the responses of ecological systems to perturbations. Ecology 78(3), 653–665 (1997)
M.G. Neubert, T. Klanjscek, H. Caswell, Reactivity and transient dynamics of predator–prey and food web models. Ecol. Model. 179(1), 29–38 (2004)
A.J. Nicholson, An outline of the dynamics of animal populations. Aust. J. Zool. 2(1), 9–65 (1954)
P. Paul, B. Ghosh, T.K. Kar, Impact of species enrichment and fishing mortality in three species food chain models. Commun. Nonlinear Sci. Numer. Simul. 29(1–3), 208–223 (2015)
P. Paul, T.K. Kar, A. Ghorai, Ecotourism and fishing in a common ground of two interacting species. Ecol. Model. 328, 1–13 (2016)
S.L. Pimm, J.H. Lawton, Number of trophic levels in ecological communities. Nature 268(5618), 329–331 (1977)
M.L. Rosenzweig, R.H. MacArthur, Graphical representation and stability conditions of predator-prey interactions. Am. Nat. 97(895), 209–223 (1963)
P.J. Schmid, D.S. Henningson, D.F. Jankowski, Stability and transition in shear flows. applied mathematical sciences, vol. 142. Appl. Mech. Rev. 55(3), B57–B59 (2002)
I. Stott, Perturbation analysis of transient population dynamics using matrix projection models. Methods Ecol. Evolut. 7(6), 666–678 (2016)
Y. Tian, T. Zhang, K. Sun, Dynamics analysis of a pest management prey–predator model by means of interval state monitoring and control. Nonlinear Anal. Hybrid Syst. 23, 122–141 (2017)
A. Verdy, H. Caswell, Sensitivity analysis of reactive ecological dynamics. Bull. Math. Biol. 70(6), 1634–1659 (2008)
R. Vesipa, L. Ridolfi, Impact of seasonal forcing on reactive ecological systems. J. Theor. Biol. 419, 23–35 (2017)
C.J. Walters, V. Christensen, S.J. Martell, J.F. Kitchell, Possible ecosystem impacts of applying MSY policies from single-species assessment. ICES J. Mar. Sci. 62(3), 558–568 (2005)
H. Wang, J. Zhang, X.-S. Yang, The maximum amplification of perturbations in ecological systems. Int. J. Biomath. 10(01), 1750009 (2017)
S. Yuan, D. Wu, G. Lan, H. Wang, Noise-induced transitions in a nonsmooth producer-grazer model with stoichiometric constraints. Bull. Math. Biol. 82(5), 1–22 (2020)
Y. Zhao, L. You, D. Burkow, S. Yuan, Optimal harvesting strategy of a stochastic inshore-offshore hairtail fishery model driven by lévy jumps in a polluted environment. Nonlinear Dyn. 95(2), 1529–1548 (2019)
Acknowledgements
Prosenjit Paul is thankful to Council of Scientific and Industrial Research (CSIR), New Delhi, India, for providing financial support in form of Research Associate fellowship (NO.25(0300)/19/EMR-II, dated: 16th May, 2019). The research of T.K. Kar is supported by the Council of Scientific and Industrial Research (CSIR), No. 25(0300)/19/EMR-II, dated: 16th May, 2019.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Declarations
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Authorship
This manuscript is prepared by Prosenjit Paul, T.K.Kar and Esita Das; Prosenjit Paul and T.K.Kar are taken the main role to design the approach, perform the analysis and write the manuscript, with substantial input from Esita Das.
Appendix
Appendix
Appendix A. Proof of Proposition 1.
These parts give the technical proof of the results of the analysis conducted in Sect. 3
The interior equilibrium point \((P_{x1}, P_{y1})\) is given by \(P_{x1}=\frac{m_{y}}{\alpha _{y}}\) and \(P_{y1}=\frac{r_{x}}{\alpha _{x}}(1-\frac{m_{y}}{\alpha _{y}k_{x}})-\frac{q_{x}e_{x}}{\alpha _{x}}\) provided \(e_{x}<\frac{r_{x}}{q_{x}}(1-\frac{m_{y}}{k_{x}\alpha _{y}})\).
The Hermitian matrix is given by at \((P_{x1}, P_{y1})\)
and its characteristic equation is \(det(H(J_{1})-\lambda I)=\lambda ^2-(trac(H(J_{1})))\lambda +det(H(J_{1}))=0\) where \(trac(H(J_{1}))=-\frac{r_{x} P_{x1}}{k_{x}}\) and \(det(H(J_{1}))= \frac{(-\alpha _{x} P_{x1}+\alpha _{y} P_{y1})^2}{4}\).
The eigenvalues are: \(\lambda =\frac{\sqrt{ \left( trac(H(J_{1})) \right) ^2-4 Det(H(J_{1}))}+trac(H(J_{1}))}{2}\)
The largest part of the eigenvalues of the matrix \(H(J_{1})\) is given by
Therefore, the stable equilibrium point \((P_{x1}, P_{y1})\) is reactive under the disturbance made by prey harvesting if \(\lambda _{1}(H(J_{1}))>0\), i.e., either \(\alpha _{x} P_{x1}-\alpha _{y} P_{y1}<0\) or, \(\alpha _{x} P_{x1}-\alpha _{y} P_{y1}>0\)
i.e., either \(e_{x}<e_{x1}\) or, \(e_{x}>e_{x1}\) where \(e_{x1}=\frac{\alpha _{x}}{\alpha _{y}q_{x}}\left[ \frac{\alpha _{y}r_{x}}{\alpha _{x}}(1-\frac{m_{y}}{k_{x}\alpha _{y}})-\frac{\alpha _{x}m_{y}}{\alpha _{y}} \right] \). Clearly, \(P_{y1}=0\implies e_{xext}=\frac{r_{x}}{q_{x}}(1-\frac{m_{y}}{k_{x}\alpha _{y}})\)
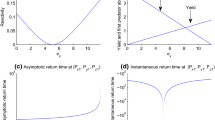
Hence, predator species goes to extinction at \(e_{xext}=\frac{r_{x}}{q_{x}}(1-\frac{m_{y}}{k_{x}\alpha _{y}})\) Now \(\frac{\hbox {d}\lambda _1(H(J_{1}))}{\hbox {d}e_{x}}<0\) when \(e_{x}\in (0, e_{x1})\) and \(\frac{\hbox {d}\lambda _1(H(J_{1}))}{\hbox {d}e_{x}}>0\) when \(e_{x}\in (e_{x1}, e_{xext})\). The yield \(Y_{x}=\frac{q_{x}e_{x}m_{y}}{\alpha _{y}}\) has no maximum in \(e_{x}\in (e_{x1}, e_{xext})\).
Therefore, the statements contain in Proposition 1 is proved and given below:
-
1.
If \(e_{x}\in (0, e_{x1})\), the stable equilibrium point \((P_{x1},P_{y1})\) remains reactive (\(\lambda _1(H(J_{1}))>0\)) under the disturbance made by prey harvesting and the strength of reactivity decreases with \(e_{x}\).
-
2.
If \(e_{x}\in (e_{x1}, e_{xext})\), the stable equilibrium point \((P_{x1},P_{y1})\) remains also reactive (\(\lambda _1(H(J_{1}))>0\)) under the disturbance made by prey harvesting and the strength of reactivity increases with \(e_{x}\) but MSY rule does not hold good. (see Fig. 1)
Appendix B. Proof of Proposition 2.
The interior equilibrium of model (2) is given by \((P_{x2}, P_{y2})\):
where \(e_{y}<\frac{\alpha _{y}k_{x}-m_{y}}{q_{y}}\).
Following the method which already applied in “Appendix A”, we get the largest eigenvalue of the Hermitian matrix is given by at (\((P_{x2}, P_{y2})\)):
Therefore, the stable equilibrium point \((P_{x2}, P_{y2})\) is reactive under the disturbance made by prey harvesting if \(\lambda _{2}(H(J_{2}))>0\), i.e., either \(\alpha _{x} P_{x2}-\alpha _{y} P_{y2}<0\) or, \(\alpha _{x} P_{x2}-\alpha _{y} P_{y2}>0\)
i.e., either \(e_{y}<e_{y1}\) or, \(e_{y}>e_{y1}\) where \(e_{y1}=\frac{1}{q_{y}}\left( \frac{\alpha _{y}^{2}r_{x}k_{x}}{\alpha _{y}r_{x}+\alpha _{x}^{2}k_{x}}-m_{y} \right) \)
Clearly, \(P_{y2}=0\implies e_{yext}=\frac{\alpha _{y}k_{x}-m_{y}}{q_{y}}\)
Hence, predator species goes to extinction at \(e_{yext}=\frac{\alpha _{y}k_{x}-m_{y}}{q_{y}}\)
Now \(\frac{\hbox {d}\lambda _2(H(J_{2}))}{\hbox {d}e_{y}}<0\) when \(e_{y}\in (0, e_{y1})\) and \(\frac{\hbox {d}\lambda _2(H(J_{2}))}{\hbox {d}e_{y}}>0\) when \(e_{y}\in (e_{y1}, e_{yext})\).
The yield \(Y_{y}=\frac{q_{y}e_{y}r_{x}}{\alpha _{x}}(1-\frac{m_{y}+q_{y}e_{y}}{\alpha _{y}k_{x}})\) has maximum at \(e_{ymax}=\frac{e_{yext}}{2}\) in \(e_{y}\in (e_{y1}, e_{yext})\).
since \(\frac{\hbox {d}Y_{y}}{\hbox {d}e_{y}}=0\implies e_{ymax}=\frac{\alpha _{y}k_{x}-m_{y}}{2 q_{y}}\) and \(\frac{\hbox {d}^2Y_{y}}{\hbox {d}e_{y}^2}<0\) at \(e_{y}=e_{ymax}\)
Therefore, the statement contained in Proposition 2 is proved and given below:
-
1.
If \(e_{y}\in (0, e_{y1})\), the stable equilibrium point \((P_{x2},P_{y2})\) remains reactive (\(\lambda _1(H(J_{2}))>0\)) under the predator harvesting and the strength of reactivity decreases with \(e_{y}\).
-
2.
If \(e_{y}{\in } (e_{y1}, e_{yext})\), the stable equilibrium point \((P_{x2},P_{y2})\) remains reactive (\(\lambda _1(H(J_{2}))>0\)) under the disturbance made by predator harvesting and the strength of reactivity increases with \(e_{y}\) and MSY rule holds good. (see Fig. 3)
Appendix C. Proof of Proposition 3.
The interior equilibrium of model (3) is given by \((P_{x3}, P_{y3})\):
where \(\alpha _{y}k_{x}q_{x}e_{x}+r_{x}q_{y}e_{y}<r_{x}\left( \alpha _{y}k_{x}-m_{y}\right) \).
Following the method which already applied in “Appendix A”, we get the largest eigenvalue of the Hermitian matrix is given by at \((P_{x3}, P_{y3})\):
Therefore, the stable equilibrium point \((P_{x3}, P_{y3})\) is reactive under the disturbance made by selective harvesting on both (prey and predator) if \(\lambda _{3}(H(J_{3}))>0\), i.e., either \(\alpha _{x} P_{x3}-\alpha _{y} P_{y3}<0\) or, \(\alpha _{x} P_{x3}-\alpha _{y} P_{y3}>0\)
i.e., either \(\left( \alpha _{x}^2k_{x} +\alpha _{y} r_{x}\right) q_{y}e_{y}+\alpha _{y}^2 k_{x} q_{x} e_{x}\le \alpha _{y}r_{x}\left( \alpha _{y}k_{x}-m_{y}\right) -\alpha _{x}^2 k_{x}m_{y} \) or, \(\left( \alpha _{x}^2 +\alpha _{y} r_{x}\right) q_{y}e_{y}+\alpha _{y}^2 k_{x} q_{x} e_{x}\ge \alpha _{y}r_{x}\left( \alpha _{y}k_{x}-m_{y}\right) -\alpha _{x}^2 k_{x}m_{y} \).
Clearly, \(P_{y3}=0\implies \alpha _{y}k_{x}q_{x}e_{x}+r_{x}q_{y}e_{y}=r_{x}\left( \alpha _{y}k_{x}-m_{y}\right) \)
Hence, predator species goes to extinction when \(\alpha _{y}k_{x}q_{x}e_{x}+r_{x}q_{y}e_{y}=r_{x}\left( \alpha _{y}k_{x}-m_{y}\right) \)
The yield \(Y_{xy}=q_{x}e_{x}\left( \frac{m_{y}+q_{y}e_{y}}{\alpha _{y}}\right) +q_{y}e_{y}\left( \frac{\alpha _{y}k_{x}( r_{x}-q_{x}e_{x})-(q_{y}e_{y}+m_{y})r_{x}}{\alpha _{x}\alpha _{y}k_{x}}\right) \) has no maximum.
Let
Clearly, \(\frac{\partial ^2Y_{xy}}{\partial e_{x}^{2}}=0\) and implies the Hessian matrix corresponding to \(Y_{xy}\) is not negative definite. Consequently, \(Y_{xy}\) has no maximum and MSTY does not exist.
Therefore, the statements contained in Proposition 3 are proved and given below:
-
1.
If \((e_{x}, e_{y})\in R_{xy}^{-}\) and \(R_{xy}^{+}\), the stable equilibrium point \((P_{x3},P_{y3})\) remains reactive (\(\lambda _3(H(J_{3}))>0\)) under the multiple disturbances made by selective harvesting on both.
-
2.
If \((e_{x}, e_{y})\in R_{xy1}\subseteq R_{xy}^{+}\), the reactivity of stable equilibrium point \((P_{x3},P_{y3})\) may be implied extinction of predator species and consequently MSTY rule not holds good. (see Fig. 5)
Appendix A*. Proof of Proposition 1*.
These parts give the technical proof of the results of the analysis conducted in Sect. 3
The interior equilibrium point \((P_{x1*}, P_{y1*})\) is given by
The Hermitian matrix is given by at \((P_{x1*}, P_{y1*})\)
and its characteristic equation is \(det(H(J_{1*})-\lambda I)=\lambda ^2-(trac(H(J_{1*})))\lambda +det(H(J_{1*}))=0\) where \(trac(H(J_{1*}))=-\left( \frac{r_{x} P_{x1}}{k_{x}}+\gamma _{y}P_{y1*}\right) \) and \(det(H(J_{1*}))=\frac{r_{x}\gamma _{y} P_{x1*}P_{y1*}}{k_{x}} -\frac{(-\alpha _{x} P_{x1}+\alpha _{y} P_{y1})^2}{4}\).
The eigenvalues are: \(\lambda =\frac{\sqrt{ \left( trac(H(J_{1*})) \right) ^2-4 Det(H(J_{1*}))} \pm trac(H(J_{1*}))}{2}\)
The largest part of the eigenvalues of the matrix \(H(J_{1*})\) is given by
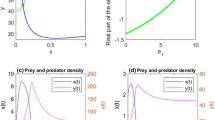
Now \(\lambda _{1*}(H(J_{1*}))=0\implies k_{x}\left( \alpha _{y} P_{y1*}-\alpha _{x} P_{x1*}\right) ^2=4 r_{x}\gamma _{y}P_{x1*}P_{y1*} \)
Clearly, \(k_{x}\left( \alpha _{y} P_{y1*}-\alpha _{x} P_{x1*}\right) ^2=4 r_{x}\gamma _{y}P_{x1*}P_{y1*}\) is a quadratic equation of prey harvesting effort \(e_{x}\).
Hence, the above equation has two roots of \(e_{x}\) say \(e_{x1*}\) and \(e_{x2*}\) with \(0<e_{x1*}<e_{x2*}\). Now \(P_{y1*}=0\implies e_{x1ext}=\frac{\left( \alpha _{y}k_{x}-m_{y} \right) r_{x} }{\alpha _{y}k_{x}q_{x}}\)
Therefore, predator species goes to extinction at \(e_{x}=e_{x1ext}\)
Clearly, \(\lambda _{1*}(H(J_{1*}))>0\) when \(e_{x}\in (0, e_{x1*})\) and \(e_{x}\in (e_{x2*}, e_{x1ext})\) and \(\lambda _{1*}(H(J_{1*}))<0\) when \(e_{x}\in (e_{x1*}, e_{x2*})\)
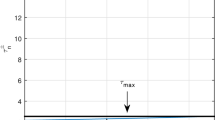
The yield \(Y_{x1}=\frac{q_{x}e_{x}k_{x} \left( \alpha _{x} m_{y}+\gamma _{y} r_{x}-\gamma _{y} q_{x} e_{x} \right) }{\alpha _{x} \alpha _{y} k_{x}+\gamma _{y} r_{x}}\) has maximum at \(e_{x1max}=\frac{\alpha _{x}m_{y}+\gamma _{y}r_{x}}{2q_{x}\gamma _{y}}\)
since \(\frac{\hbox {d}Y_{x1}}{\hbox {d}e_{x}}=0\implies e_{x1max}=\frac{\alpha _{x}m_{y}+\gamma _{y}r_{x}}{2q_{x}\gamma _{y}}\) and \(\frac{\hbox {d}^2Y_{x1}}{\hbox {d}e_{x}^2}<0\) at \(e_{x}=e_{x1max}\)
Therefore, the statements contained in Proposition 1* are proved and given below:
-
1.
If \(e_{x}\in (0, e_{x1*})\), then the stable equilibrium point \((P_{x1*},P_{y1*})\) remains reactive (\(\lambda _1*(H(J_{1*}))>0\)) under the disturbance made by prey harvesting and the strength of reactivity decreases with \(e_{x}\).
-
2.
If \(e_{x}\in (e_{x1*}, e_{x2*})\), then the stable equilibrium point \((P_{x1*},P_{y1*})\) remains non-reactive (\(\lambda _1*(H(J_{1*}))<0\)) under the disturbance made by prey harvesting and MSY rule holds good.
-
3.
If \(e_{x}\in (e_{x2*}, e_{x1ext})\), then the stable equilibrium point \((P_{x1*},P_{y1*})\) remains again reactive (\(\lambda _1*(H(J_{1*}))>0\)) under the disturbance made by prey harvesting (see Fig. 6).
Appendix B*. Proof of Proposition 2*.
The interior equilibrium of model (6) is given by \((P_{x2*}, P_{y2*})\):
Following the method which already applied in “Appendix A*”, we get the largest eigenvalue of the Hermitian matrix is given by at (\((P_{x2*}, P_{y2*})\)): The largest part of the eigenvalues of the matrix \(H(J_{2*})\) is given by
Now \(\lambda _{2*}(H(J_{2*}))=0\implies k_{x}\left( \alpha _{y} P_{y2*}-\alpha _{x} P_{x2*}\right) ^2=4 r_{x}\gamma _{y}P_{x2*}P_{y2*} \)
Clearly, \(k_{x}\left( \alpha _{y} P_{y2*}-\alpha _{x} P_{x2*}\right) ^2=4 r_{x}\gamma _{y}P_{x2*}P_{y2*}\) is a quadratic equation of prey harvesting effort \(e_{y}\).
Hence, the above equation has two roots of \(e_{y}\) say \(e_{y1*}\) and \(e_{y2*}\) with \(0<e_{y1*}<e_{y2*}\). Now \(P_{y2*}=0\implies e_{y1ext}=\frac{\left( \alpha _{y}k_{x}-m_{y}\right) r_{x}}{r_{x}q_{y}}\)
Therefore, predator species goes to extinction at \(e_{y}=e_{y1ext}\)
Clearly, \(\lambda _{2*}(H(J_{2*}))>0\) when \(e_{y}\in (0, e_{y1*})\) and \(e_{y}\in (e_{y2*}, e_{y1ext})\) and \(\lambda _{2*}(H(J_{2*}))<0\) when \(e_{y}\in (e_{y1*}, e_{y2*})\)
The yield \(Y_{y1}=\frac{q_{y}e_{y}\left( \alpha _{y}k_{x}r_{x}-r_{x}m_{y}-r_{x}q_{y}e_{y}\right) }{\alpha _{x}\alpha _{y}k_{x}+\gamma _{y}r_{x}}\) has maximum at \(e_{y1max}=\frac{\alpha _{x}m_{y}+\gamma _{y}r_{x}}{2q_{x}\gamma _{y}}\)
since \(\frac{\hbox {d}Y_{x1}}{\hbox {d}e_{x}}=0\implies e_{x1max}=\frac{\left( \alpha _{y}k_{x}-m_{y}\right) r_{x}}{2r_{x}q_{y}}\) and \(\frac{\hbox {d}^2Y_{y1}}{\hbox {d}e_{y}^2}<0\) at \(e_{y}=e_{y1max}\)
Therefore, the statements contained in Proposition 2* are proved and given below:
-
1.
If \(e_{y}\in (0, e_{y1*})\), then the stable equilibrium point \((P_{x2*},P_{y2*})\) remains reactive (\(\lambda _2*(H(J_{2*}))>0\)) under the disturbance made by predator harvesting and the strength of reactivity decreases with \(e_{y}\).
-
2.
If \(e_{y}\in (e_{y1*}, e_{y2*})\), then the stable equilibrium point \((P_{x2*},P_{y2*})\) remains non-reactive (\(\lambda _2*(H(J_{2*}))<0\)) under the disturbance made by predator harvesting.
-
3.
If \(e_{y}\in (e_{y2*}, e_{y1ext})\), then the stable equilibrium point \((P_{x2*},P_{y2*})\) remains again reactive (\(\lambda _2*(H(J_{2*}))>0\)) under the disturbance made by predator harvesting and MSY rule holds good. (see Fig. 8)
Appendix C*. Proof of Proposition 3*.
The interior equilibrium of model (7) is given by \((P_{x3*}, P_{y3*})\):
where \(\gamma _{y}k_{x}q_{x}e_{x}-\alpha _{x}k_{x}q_{y}e_{y}<\alpha _{x}k_{x}m_{y}+\gamma _{y}k_{x}r_{x}\) and \(\alpha _{y} k_{x}q_{x}e_{x}+r_{x}q_{y}e_{y}<\left( \alpha _{y}k_{x}-m_{y}\right) r_{x}\).
Following the method which already applied in “Appendix A*”, we get the largest eigenvalue of the Hermitian matrix given by at \((P_{x3*}, P_{y3*})\):
Therefore, the stable equilibrium point \((P_{x3*}, P_{y3*})\) is reactive under the disturbance made by selective harvesting on both (prey and predator) if \(\lambda _{3*}(H(J_{3*}))>0\), i.e., either \(det(H(J_{3*}))<0\) or, \(det(H(J_{3*}))>0\)
i.e., \(k_{x}\left( \alpha _{y} P_{y3*}-\alpha _{x} P_{x3*}\right) ^2=4 r_{x}\gamma _{y}P_{x3*}P_{y3*}\).
Clearly, \(P_{y3*}=0\implies \alpha _{y} k_{x}q_{x}e_{x}+r_{x}q_{y}e_{y}=\left( \alpha _{y}k_{x}-m_{y}\right) r_{x}\)
Hence, predator species goes to extinction when \(\alpha _{y} k_{x}q_{x}e_{x}+r_{x}q_{y}e_{y}=\left( \alpha _{y}k_{x}-m_{y}\right) r_{x}\)
The yield \(Y_{xy1}=q_{x}e_{x}\frac{\alpha _{x}k_{x}\left( e_{y}q_{y}+m_{y}\right) +\gamma _{y}k_{y} \left( r_{x}-q_{x}e_{x}\right) }{\alpha _{x} \alpha _{y}k_{x}+\gamma _{y}r_{y}} +q_{y}e_{y}\frac{\alpha _{y}k_{x}r_{x}-m_{y}r_{x}-\alpha _{y}k_{x}q_{x}e_{x}-r_{x}q_{y}e_{y}}{\alpha _{x}\alpha _{y} k_{x}+\gamma _{y}r_{x}}\) has maximum, since the Hessian matrix corresponding to \(Y_{xy1}\) is negative definite.
Let
Clearly, \(\frac{\partial ^2Y_{xy}}{\partial e_{x}^{2}}<0\) and \(det(H_{xy1})>0\) which implies the Hessian matrix corresponding to \(Y_{xy1}\) is negative definite. Consequently, \(Y_{xy1}\) has maximum value and MSTY exists (see Fig. 10).
Therefore, the statements contained in Proposition 3* are proved and given below:
-
1.
If \((e_{x}, e_{y})\in R_{xy1}\), the stable equilibrium point \((P_{x3*},P_{y3*})\) remains reactive (\(\lambda _3*(H(J_{3*}))>0\)) under the multiple disturbances made by selective harvesting on both and MSTY rule holds good (see Fig. 10).
Rights and permissions
About this article

Cite this article
Paul, P., Kar, T.K. & Das, E. Reactivity in prey–predator models at equilibrium under selective harvesting efforts. Eur. Phys. J. Plus 136, 510 (2021). https://doi.org/10.1140/epjp/s13360-021-01525-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1140/epjp/s13360-021-01525-6