
Abstract
Classical evolutionary game theory (EGT) focuses on competition among phenotypes while assuming asexual transmission of these phenotypes to the next generation. However, phenotypic selection and sexual recombination are not necessarily mutually reinforcing in populations with sexual reproduction. In particular, it has been long known that some of the evolutionarily stable strategies derived by EGT methods cannot be achieved by sexually reproducing, real-world, populations. Thus, the recently formulated polymorphic evolutionary game theory (PEGT), which adds underlying genetics and sexual reproduction to evolutionary games, has the potential to revolutionize game theoretical modeling of coevolutionary processes. To illustrate the advantages of PEGT over classical EGT, I analyze two of the best known EGT models: Hawk/Retaliator/Dove and Defector/Tit-for-Tat/Altruist, by PEGT methods. I show that if one admits non-Mendelian genetics—common in heritable behavior, both of these games exhibit the properties of moderated aggression and conditional cooperation as components of population-level polymorphisms.


Similar content being viewed by others
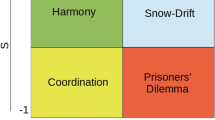

Notes
I prefer to defer modeling non-random mating to asymmetric PEGT games.
References
Alcock J (1993) Animal behavior: an evolutionary approach. Sinauer Associates, Sunderland
Allchin D (2000) Mending mendelism. Am Biol Teach 62(9):633–640
Arnott G, Elwood RW (2008) Information gathering and decision making about resource value in animal contests. Anim Behav 76(3):529–542
Begon M, Harper JL, Townsend CR (1990) Ecology: individuals, populations and communities. Blackwell, Cambridge
Catling DC, Claire MW (2005) How Earth’s atmosphere evolved to an oxic state: a status report. Earth Planet Sci Lett 237(1):1–20
Davis KL, Karpanty SM, Spendelow JA, Cohen JB, Althouse MA, Parsons KC, Luttazi CF (2019) Begging behavior as an honest signal of need and parent–offspring association during the postfledging dependency period. Ecol Evol 9:7497–7508
de Boer SF (2016) Animal models: implications for human aggression and violence. In: Aggression and violence. Routledge, pp 32–54
de Boer SF, van der Vegt BJ, Koolhaas JM (2003) Individual variation in aggression of feral rodent strains: a standard for the genetics of aggression and violence? Behav Gen 33(5):485–501
Fijarczyk A, Babik W (2015) Detecting balancing selection in genomes: limits and prospects. Mol Ecol 24(14):3529–3545
Fishman MA (2006) Involuntary defection and the evolutionary origins of empathy. J Theor Biol 242:873–879
Fishman MA (2008) Asymmetric evolutionary games with non-linear pure strategy payoffs. Games Econ Behav 63:77–90
Fishman MA (2016) Polymorphic evolutionary games. J Theor Biol 398:130–135
Fishman MA (2019) Animal conflicts in diploid populations with sexual reproduction. J Theor Biol 462:475–478
Gilbert OM (2018) Altruism or association? Proc Nat Acad Sci 115(14):E3069–E3070
Hofbauer J, Sigmund K (1998) Evolutionary games and population dynamics. Cambridge University Press, Cambridge, p 65
Karlin S (1975) General two-locus selection models: some objectives, results and interpretations. Theor Popul Biol 7:364–398
Karlin S, Lessard S (1986) Theoretical studies on sex ratio evolution, vol 22. Princeton University Press, Princeton
Krebs JR, Davies NB (1993) An introduction to behavioural ecology. Blackwell, Oxford
Mesterton-Gibbons M, Adams ES (1998) Animal contests as evolutionary games: paradoxical behavior can be understood in the context of evolutionary stable strategies. The trick is to discover which game the animal is playing. Am Sci 86(4):334–341
Ohta R, Kojima K (2019) Hatano rats selectively bred for high-and low-avoidance learning: an overview. Exp Anim 68(2):127–136
Sellis D, Callahan BJ, Petrov DA, Messer PW (2011) Heterozygote advantage as a natural consequence of adaptation in diploids. Proc Nat Acad Sci 108(51):20666–20671
Selten R, Hammerstein P (1984) Gaps in Harley’s argument on evolutionary stable learning rules and in the logic of TfT. Behav Brain Sci 7:115–116
Smith JM (1982) Evolution and the theory of games, vol 2, 4. Cambridge University Press, Cambridge
Smith JM, Price GR (1973) The logic of animal conflict. Nature 246(5427):15
Staes N, Koski SE, Helsen P, Fransen E, Eens M, Stevens JM (2015) Chimpanzee sociability is associated with vasopressin (Avpr1a) but not oxytocin receptor gene (OXTR) variation. Horm Behav 75:84–90
Teixeira JC, de Filippo C, Weihmann A, Meneu JR, Racimo F, Dannemann M, Atencia R (2015) Long-term balancing selection in LAD1 maintains a missense trans-species polymorphism in humans, chimpanzees, and bonobos. Mol Biol Evol 32(5):1186–1196
Van Segbroeck M, Knoll AT, Levitt P, Narayanan S (2017) MUPET—mouse ultrasonic profile extraction: a signal processing tool for rapid and unsupervised analysis of ultrasonic vocalizations. Neuron 94(3):465–485
Wang C, Lu X (2018) Hamilton’s inclusive fitness maintains heritable altruism polymorphism through rb = c. Proc Nat Acad Sci 115(8):1860–1864
Acknowledgements
Special thanks are due to Dr. Reuben Hiller for his invaluable technical assistance. I thank the unknown reviewers for suggestions that make this a better paper.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Fishman, M.A. Polymorphic Evolutionary Games and Non-Mendelian Genetics. Bull Math Biol 82, 31 (2020). https://doi.org/10.1007/s11538-020-00705-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11538-020-00705-2