
Abstract
Accounts of causal explanation are standard in philosophy of science. Less common are accounts of experimentation to investigate causal relations: detailed discussions of the specific kinds of experiments scientists design and run. Silva, Landreth, and Bickle’s (SLB) (Engineering the next revolution in neuroscience: the new science of experiment planning, Oxford University Press, New York, 2014) account of “connection experiments” derives directly from landmark experiments in “molecular and cellular cognition.” We start with its key components, and then using a detailed case study from recent social neuroscience we emphasize and extend three features of SLB’s account: (1) a division of distinct types of connection experiments, each providing a different type of evidence for a hypothesized causal relationship; (2) the typically downward-looking nature (in the sense of componentry) of the experimental search for mediating causes in mainstream neurobiology; and most importantly, (3) the centrality of multiple-experiment research programs, with each experiment designed such that if successful its results can be integrated with the others, toward the goal of confirming multiple-phenomena causal pathways. Our extension of SLB’s account complements existing philosophical work on experimentation in neurobiology.

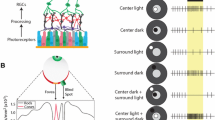
Reprinted with permission from Silva et al. (2014), figure 2.1, pg. 21

Reprinted with permission from Juruana et al. (2004), Figure 1, p. 191

Reprinted with permission from Meaney and Szyf (2005), Figure 2, pg. 109

Reprinted from Hurley (2013) with permission from Discover Magazine/Jay Smith

Similar content being viewed by others


Notes
Craver’s notion of “levels’ is the relatively innocuous one of componentry, which is the account we adopt throughout this paper. The kinds “one level down” from another are the ones of which the latter is composed. Chapter 5 of his (2007) develops this account into a “field guide” for levels in neuroscience.
In neuroscientific practice the philosophically vexing notion of a “kind” is specified by a publication-driven “ontology.” See Larson and Martone (2009).
However, citation numbers alone cannot be the entire basis for this judgment, because review articles tend to receive the highest citation numbers. SLB (2014) thus limit their metascientific considerations to a field’s experiment publications. See their chapter 7 for discussion.
An anonymous referee pointed out ways that a related field, molecular biology, has addressed the problem of an enormous published literature: by curating data from hundreds of thousands of published protein interaction studies into accessible databases. That reviewer asks how SLB’s “research maps” compare to such resources. An excellent question, but unfortunately one that even beginning to explore would take us far afield from our task in this paper.
We thank the same anonymous reviewer for pointing out this specific caveat to SLB-inspired metascience.
One of us (Bickle) has since begun a metascientific investigation of Tool Development experiments in neurobiology. See Bickle (2016) and (forthcoming).
We here specify the conditions for all three types of connection experiments for hypothesized excitatory causal connections only. For each type, the results on B will be exactly the opposite for hypothesized inhibitory causal connections, e.g., no (or diminished) A → maintained (or increased) B for a successful negative intervention into an inhibitory causal connection.
LG and ABN scores for individual mothers in the sample were based on numerous and regularly scheduled recording periods of mother–pup interactions, across both light and dark cycles. Frequency of LG episodes and of ABN postures could be calculated for each mother, and mean and standard deviation calculated for the group. High-LG-ABN mothers had both LG and ABN scores at least 1 standard deviation above group mean; low-LG-ABN mothers had both scores at least 1 standard deviation below. Incidentally, individual LG and ABN scores in mothers were highly positively correlated (Fish et al. 2004).
mRNAs result from DNA transcription, and carry the amino acid sequence for the protein product to the ribosomes, for translation and protein synthesis. A promoter is a region of DNA, upstream from the coding region of the gene, where mRNA transcription is initiated.
Methylation is the process by which methyl groups (one carbon atom bonded to three hydrogen atoms, CH3) are added to specific binding sites on the DNA molecule. Its functional significance is to suppress gene expression, discussed in the text which follows.
Weaver et al. (2004) followed a standard four-square fostering protocol. There were two control groups: offspring of low-LG-ABN mothers fostered to other low-LG-ABN mothers, and offspring of high-LG-ABN mothers fostered to other high-LG-ABN mothers.
See SLB (2014), chapter 6, where they show how integrative practices generate causal graphs with multiple, even interlevel components (e,g., activated NMDA receptors, intracellular signaling cascades in post-synaptic neurons, hippocampus CA1-region LTP, and hippocampus-based learning and memory); and chapter 7, where they show how such causal paths are generated from graphs of “Networks of EXperiments” (NEX graphs).
Histone acetylation is the process by which acetyl groups, that is, a methyl group CH3 single-bonded to a carboxyl, CO, (COCH3), are attached to lysine residues of the histone core of the nucleosome. The nucleosome is the basic unit of DNA in eukaryote cells (cells containing a nucleus), consisting of a DNA segment wound around a protein core consisting of 8 histones. Its functional significance is discussed in the text which follows.
Histone deacetylases remove acetyl groups from lysine residues in chromatin to silence transcription activity by blocking transcriptional factor binding. So histone deacetylase inhibitors like TSA block the actions of histone deacetylases, to maintain active chromatin and gene expression (with maintained promoter binding).
DNA microarray analyses are capable of measuring expression patterns simultaneously of upwards to ten thousand genes in a single tissue sample. Weaver et al. report microarray analyses of tissue from across the hippocampus, in both l-methionine-treated and vehicle-treated offspring, with results showing that “neither maternal care nor methionine treatment affected global DNA methylation levels” (2005, 11050).
More recently Meaney’s lab has found related molecular mediating causes between childhood experiences and stress responses in human brains. Work on measuring differing hippocampus GR gene expression in human brain cadaver tissue drawn from groups of age- and sex-matched controls at time of death but with different causes of death and childhood abuse backgrounds has demonstrated effects on the hippocampus NR3C1/GR 1F promoter region, the human homologue of the exon 17 promoter region in the rat (McGowan et al. 2009). These studies are not limited to strictly non-intervention experiments. Patch methylation techniques applied to hippocampus NR3C1 promoter region tissue revealed a significantly higher percentage of methylated sites in suicide completers with a history of child abuse, compared to both suicide completers without a history of child abuse and sudden non-suicide death control subjects without a history of child abuse. This percentage was most significantly higher at a site on the human GR promoter region containing an NGF1-A binding site. This experimental procedure induces methylation in DNA cloned from the cadaver samples in the presence of NGF1-A, so indicate intervention evidence for a causal relation (McGowan et al. 2009). The authors note the consistency of these studies with the rodent work discussed above, and with previous human studies on activation of HPA stress responses in victims of child abuse and the role of hippocampus GR activation to dampen HPA activity. Using these same techniques, the Meaney lab has also shown that other, nearby exon regions on the human hippocampus GR gene show similar patterns of expression and methylation across groups as the GR1F promoter region (Labonte et al. 2012). The locations of these other regions on the genome, along with the locations of numerous transcriptional binding sites in their vicinity, likewise “render them susceptible to epigenetic regulation” (Labonte et al. 2012, p. 41).
For some landmark experiments and results in this quarter-century track record, drawn directly from among the most highly-cited MCC publications, see SLB (2014), chapters 3 through 7.
The extension of the Meaney’s lab rodent work to human brain tissue and gene homologues, discussed in footnote 16 above, appear to be an example of this kind of integration within a research program. The rodent experiments emphasize the norm of reliability, while the human brain tissue experiments are more concerned with establishing the external validity of the derailed causal path “outside the laboratory,” in real-world human brains. The consistency of the expected results across species homologues seems sufficient for these experimenters to integrate their results across experiments despite vastly different subprotocols.
References
Bickle, J. (2016). Revolutions in neuroscience: Tool development. Frontiers in Systems Neuroscience, 10, 24. https://doi.org/10.3389/fnsys.2016.00024.
Bickle, J. (forthcoming). From microscopes to optogenetics: Ian Hacking vindicated. Philosophy of Science, 85(5).
Craver, C. F. (2007). Explaining the brain. New York: Oxford University Press.
Craver, C. F., & Darden, L. (2013). In search of mechanisms: Discoveries across the life sciences. Chicago: University of Chicago Press.
Fish, E. W., Shakrokh, D., Bagot, R., Caldji, C., Bredy, T., Szyf, M., et al. (2004). Epigenetic programming of stress responses through variations in maternal care. Annals of the New York Academy of Sciences, 1036, 167–180.
Francis, D., Diorio, J., Liu, D., & Meaney, M. J. (1999). Nongenomic transmission across generations of maternal behavior and stress responses in the rat. Science, 286, 1155–1158.
Gillett, C. (2017). Reduction and emergence in science and philosophy. Cambridge: Cambridge University Press.
Hurley, D. (2013). Trait versus fate. Discover Magazine, pp. 48–55.
Juruena, M. K., Cleare, A. J., & Pariante, C. M. (2004). The Hypothalamic Pituitary Adrenal axis glucocorticoid receptor function and relevance to depression. Revista Brasiliera de Psiquiatria (Brazilian Journal of Psychiatry), 26(3), 189–201.
Kandel, E. R., Schwartz, J., & Jessell, T. (Eds.). (2000). Principles of neural science (4th ed.). New York: McGraw-Hill.
Labonte, B., Yerko, V., Gross, J., Mechawar, M., Meaney, M. J., Szyf, M., et al. (2012). Differential glucocorticoid receptor exon 1B, 1C, and 1H expression and methylation in suicide completers with a history of childhood abuse. Biological Psychiatry, 72, 41–48.
Larson, S. D., & Martone, M. E. (2009). Ontologies for neuroscience: What are they and what are they good for? Frontiers in Neuroscience. https://doi.org/10.3389/neuro.01.007.2009.
McGowan, P. O., Sasaki, A., D’Alessio, A. C., Dymov, S., Labonte, B., Szyf, M., et al. (2009). Epigenetic regulation of the glucocorticoid receptor in human brain associates with childhood abuse. Nature Neuroscience, 12(3), 342–348.
Meaney, M. J., & Szyf, M. (2005). Environmental programming of stress responses through DNA methylation: Life at the interface between a dynamic environment and a fixed genome. Dialogues in Clinical Neuroscience, 7, 103–123.
Silva, A. J., Landreth, A., & Bickle, J. (2014). Engineering the next revolution in neuroscience: The new science of experiment planning. New York: Oxford University Press.
Sullivan, J. (2007). Reliability and validity of experiment in the neurobiology of learning and memory. Doctoral dissertation, Department of History and Philosophy of Science, University of Pittsburgh.
Sullivan, J. (2009). The multiplicity of experimental protocols: A challenge for reductionist and non-reductionist models of the unity of neuroscience. Synthese, 167, 511–539.
Sullivan, J. (2015). Experimentation in cognitive neuroscience and cognitive neurobiology. In J. Clausen & N. Levy (Eds.), Handbook of neuroethics (pp. 31–47). Dordreccht: Springer.
Sweatt, J. D. (2009). Mechanisms of memory (2nd ed.). San Diego: Academic Press.
Weaver, I. C. G., Cervoni, N., Champagne, F. A., Alessio, A. C. D., Sharma, S., Seckl, J. R., et al. (2004). Epigenetic programming by maternal behavior. Nature Neuroscience, 7/8, 847–854.
Weaver, I. C. G., Champagne, F. A., Brown, S. E., Dymov, S., Sharma, S., Meaney, M. J., et al. (2005). Reversal of maternal programming of stress responses in adult offspring through methyl supplementation: Altering epigenetic marking later in life. Journal of Neuroscience, 25(47), 11045–11054.
Woodward, J. (2003). Making things happen: A theory of causal explanation. New York: Oxford University Press.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Bickle, J., Kostko, A. Connection experiments in neurobiology. Synthese 195, 5271–5295 (2018). https://doi.org/10.1007/s11229-018-1838-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11229-018-1838-0