
Abstract
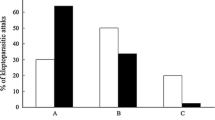
Kleptoparasitism is a well-known foraging tactic used opportunistically by many seabird species. However, little is known about this behaviour during the early stages of the breeding cycle and its effects on breeding performance. Here, we investigated the relationship between kleptoparasitism during the courtship period and female reproductive performance in Common Terns (Sterna hirundo). All identified kleptoparasites were males, and none of their mates performed such behaviour. We compared two groups of tern pairs, one where the males performed kleptoparasitism (kleptoparasitic group, n = 10), the other one where both mates were non-kleptoparasitic (honest group, n = 22). The body mass of kleptoparasitic females was between 8 and 15% higher than that of honest females. In kleptoparasitic females, the third egg was significantly bigger than in honest birds, and the egg-volume was not significantly different between the three eggs of the clutch in contrast to honest birds. We found no differences in the comparison among hatching success between both groups. The reproductive output, however, was significantly higher in the kleptoparasitic than in the honest group. Hence, we are providing the first evidence that kleptoparasitism during early stages of the breeding cycle has a strong link with egg size and reproductive output in Common Terns.


Similar content being viewed by others

References
Becker PH, Wendeln H (1997) A new application for transponders in population ecology of the common tern. Condor 99:534–538
Belisle M (1998) Foraging group size: models and a test with jaegers kleptoparasitizing terns. Ecology 79:1922–1938
Bize P, Roulin A, Richner H (2002) Covariation between egg size and rearing condition determines offspring quality: an experiment with the alpine swift. Oecologia 132:231–234
Bonato M, Evans MR, Cherry MI (2009) Investment in eggs is influenced by male coloration in the ostrich (Struthio camelus). Anim Behav 77:1027–1032
Brockmann HJ, Barnard CJ (1979) Kleptoparasitism in birds. Anim Behav 27:487–514
Brown GR, Almond REA, van Bergen Y (2004) Begging, stealing, and offering: food transfer in nonhuman primates. Adv Stud Behav 34:265–295
Cam E, Monnat JY (2000) Stratification based on reproductive state reveals contrasting patterns of age-related variation in demographic parameters in the kittiwake. Oikos 90:560–574
Cam E, Hines JE, Monnat JY, Nichols JD, Danchin E (1998) Are adult nonbreeders prudent parents? The kittiwake model. Ecology 79:2917–2930
Catry P, Ratcliffe N, Furness RW (1998) The influence of hatching date on different life-history stages of great skuas Catharacta skua. J Avian Biol 29:299–300
Christians JK (2002) Avian egg size: variation within species and inflexibility within individuals. Biol Rev 77:1–26
Cooper WE, Pérez-Mellado V (2003) Kleptoparasitism in the Balearic lizard, Podarcis lilfordi. Amphibia-Reptilia 24:219–224
Coulson JC (1968) Differences in the quality of birds nesting in the center and on the edge of a colony. Nature 217:478–479
Coulson JC, Porter JM (1985) Reproductive success of the kittiwake Rissa tridactyla: the roles of clutch size, chick growth rates and parental quality. Ibis 127:450–466
Crawley MJ (2007) The R book. Wiley, West Sussex, UK
Furness RW (1987) Kleptoparasitism in seabirds. In: Croxal JP (ed) Seabirds, feeding biology and role in marine ecosystem. Cambridge University Press, Cambridge, pp 77–99
García GO, Favero M, Vassallo AI (2010) Factors affecting kleptoparasitism by Gulls in a multi-species seabird colony. Condor (in press)
González-Solís J, Becker PH, Wendeln H (1999) Divorce and asynchronous arrival in common terns, Sterna hirundo. Anim Behav 58:1123–1129
González-Solís J, Sokolov E, Becker PH (2001) Courtship feedings, copulations and paternity in common terns Sterna hirundo. Anim Behav 61:1125–1132
González-Solís J, Becker PH, Jover L, Ruiz X (2004) Individual changes underlies age-specific patterns of laying date and egg-size in female common terns. J Ornithol 145:129–136
Grimm MP, Klinge M (1996) Pike and some aspects of its dependence on vegetation. In: Craig JF (ed) Pike: biology and exploitation. Chapman and Hall, New York, pp 125–126
Harris MP (1964) Aspects of the breeding biology of the gulls Larus argentatus, L. fuscus and L. marinus. Ibis 106:432–456
Hays H (1970) Common terns pirating fish on Great Gull Island. Wilson Bull 12:99–100
Hudson AV, Furness RW (1988) Utilisation of discarded fish by scavenging seabirds behind whitefish trawlers in Shetland. J Zool 215:151–166
Hunt GL (1980) Mate selection and mating systems in seabirds. In: Burger J, Olla B, Winn H (eds) Behavior of marine animals, vol. 4, Marine birds. Plenum, New York, pp 113–144
Iyengar EV (2004) Host-specific performance and host use in the kleptoparasitic marine snail Trichotropis cancellata. Oecologia 138:628–639
Järvinen AR, Vaisanen A (1983) Egg size and related reproductive traits in a southern passerine Ficedula hypoleuca breeding in an extreme northern environment. Ornis Scand 14:253–262
Kahn NW, John JS, Quinn W (1998) Chromosome-specific intron size differences in the avian method for sex identification in birds. Auk 115:1074–1078
Kruuk H (1972) The spotted hyena: a study of predation and social behaviour. University of Chicago Press, Chicago
Lack D (1940) Courtship feeding in birds. Auk 57:169–178
Lindström J (1999) Early development and fitness in birds and mammals. Trends Ecol Evol 14:343–348
Ludwigs JD (1998) Kleptoparasitismus bei der Flußseeschwalbe Sterna hirundo als Anzeiger für. Nahrungsmangel. Vogelwelt 119:193–203
Mills JA (1989) Red-billed Gull. In: Newton I (ed) Lifetime reproduction in birds. Academic, London, pp 387–404
Morand-Ferron J, Sol D, Lefebvre L (2007) Food-stealing in birds: brain or brawn? Anim Behav 74:1725–1734
Nager RG, Zandt HS (1994) Variation in egg size in great tits. Ardea 82:315–328
Nisbet ICT (1973) Courtship-feeding, egg-size and breeding success in common terns. Nature 241:141–142
Nisbet ICT (1977) Courtship-feeding and clutch size in common terns Sterna hirundo. In: Stonehouse B, Perrins C (eds) Evolutionary ecology. Macmillan, London, pp 101–109
Nisbet ICT, Spendelow JA, Hatfield JS, Zingo JM, Gough GA (1998) Variations in growth of roseate tern chicks. II. Early growth as an index of parental quality. Condor 100:305–315
Ojanen M, Orell M, Vasisanen RA (1979) Role of heredity in egg size variation in the great tit Parus major and pied flycatcher Ficedula hypoleuca. Ornis Scand 10:22–28
Ojanen M, Orell M, Vasisanen RA (1981) Egg size variation within passerine clutches: effects of ambient temperature and laying sequence. Ornis Fenn 58:93–108
Parsons J (1976) Factors determining the number and size of eggs laid by the herring gull. Condor 78:481–492
Pinheiro JC, Bates DM (2000) Mixed-effects models in S and S-PLUS. Springer, Berlin
Reid WV (1987) Constraints on clutch size in the Glaucous-winged Gull. In: Hand JL, Southern WE, Vermeer K (eds) Ecology and behavior of Gulls: Proceedings of an International Symposium of the Colonial Waterbird Group and the Pacific Seabird Group, 6 December 1985, San Francisco, California: Stud Avian Biol, no. 10. pp 8–25
Reid WV, Boersma PD (1990) Parental quality and selection on egg size in the magellanic penguin. Evolution 44:1780–1786
Salzer D, Larkin G (1990) Impact of courtship feeding on clutch and third-egg size in glaucous-winged gulls. Anim Behav 39:1149–1162
Shealer DA, Spendelow JA (2002) Individual foraging strategies of kleptoparasitic roseate terns. Waterbirds 25:436–441
Shealer DA, Spendelow JA, Harfield JS, Nisbet ICT (2005) The adaptive significance of stealing in a marine bird and its relationship to parental quality. Behav Ecol 16:371–376
Silva MC, Boersma PD, Mackay S, Strange I (2007) Egg size and parental quality in thin-billed prions, Pachyptila belcheri: effects on offspring fitness. Anim Behav 74:1403–1412
Slagsvold T, Sandvikg R, Lorentsen MH (1984) On the adaptive value of intra-clutch egg-size variation in birds. Auk 101:685–697
Steele WK, Hockey PAR (1995) Factors influencing the rate and success of intraspecific kleptoparasitism among kelp gulls (Larus dominicanus). Auk 112:847–859
Stienen EWM (2006) Living with gulls: trading off food and predation in the Sandwich Tern Sterna sandvicensis. PhD dissertation, University of Groningen, Netherlands
Sudmann S, von Rienen F (2000) Fernausgelöste Farbspritzanlagen zur individuellen Kennzeichnung von Vögeln. Vogelwarte 40:319–322
Tasker CR, Mills JA (1981) A functional analysis of courtship feeding in the red-billed gull, Larus novaehollandiae scopulinus. Behaviour 77:222–241
R Development Core Team (2008) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. ISBN 3-900051-07-0. http://www.R-project.org
Vollrath F (1984) Kleptobiotic interactions in invertebrates. In: Barnard CJ (ed) Producers and scroungers: strategies of exploitation and parasitism. Croom Helm, London, pp 61–94
Wagener M (1998) Praktische Hinweise für brutbiologische Untersuchungen an der Flussseeschwalbe Sterna hirundo. Vogelwelt 119:279–286
Wendeln H (1997) Body mass of female common terns (Sterna hirundo) during courtship: relationship to male quality, egg mass, diet, laying date and age. Col Waterbirds 20:235–243
Wendeln H, Becker PH (1996) Body mass change in breeding common terns Sterna hirundo. Bird Study 43:85–95
Wendeln H, Becker PH (1999) Effects of parental quality and effort on the reproduction of common terns. J Anim Ecol 68:205–214
Williams TD (1994) Intraspecific variation in egg size and egg composition in birds: effects on offspring fitness. Biol Rev 68:35–59
Winkler DW (1985) Factors determining a clutch size reduction in California gulls (Larus californicus): a multi-hypothesis approach. Evolution 39:667–677
Wooller RD, Dunlop JN (1981) Annual variation in clutch and egg sizes in the silver gull Larus novaehollandiae. Aust Wildl Res 8:431–433
Acknowledgments
We would like to thank Götz Wagenknecht for logistic assistance. Thanks to Christina Bauch, Juliane Riechert and Alexander Braasch for providing helpful comments at different stages of the study. Thanks to Juliane Riechert for compiling data and to Walter Tito Svagelj for providing statistical help. Germán García was supported by ALEARR Program (Ministerio de Educación de la República Argentina, Argentina; DAAD-Deutscher Akademischer Austausch Dienst, Germany), and doctoral scholarships from CONICET (Argentina). This work was supported by the Deutsche Forschungsgemeinschaft (Be 916/8-3).
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by T. Friedl.
Rights and permissions
About this article
Cite this article
García, G.O., Becker, P.H. & Favero, M. Kleptoparasitism during courtship in Sterna hirundo and its relationship with female reproductive performance. J Ornithol 152, 103–110 (2011). https://doi.org/10.1007/s10336-010-0554-4
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10336-010-0554-4