
Abstract
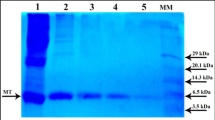
A cadmium-binding protein was purified from the digestive gland of the Antarctic scallop, Adamussium colbecki, and biochemically characterized. Purification procedures included gel permeation and anion exchange chromatography, followed by preparative polyacrylamide gel electrophoresis. Our results demonstrate that the A. colbecki cadmium-binding protein has the general properties of metallothioneins: low molecular weight of about 10 kDa, spectroscopic features typical of cadmium thiolate clusters and high metal (cadmium) content. Analysis of amino acid composition reveals the absence of aromatic amino acids, histidine, methionine and arginine. Asparagine and glutamine are also absent. The A. colbecki metallothionein shows high levels of glycine (14%), aspartic acid (14%), glutamic acid (11%) and a low lysine content (4%); the A. colbecki metallothionein shows a lower cysteine content (12%) compared to other metallothioneins (17–30%) purified from both vertebrate and invertebrate organisms. The presence of a metallothionein in the digestive gland of A. colbecki suggests that in cold-ocean-adapted molluscs the heavy metal homeostasis mechanisms may have evolved similarly to those of organisms living in temperate marine environments, although the A. colbecki cadmium-binding protein shows a typical amino acidic composition that might reflect a peculiar physiological role.
Similar content being viewed by others

Author information
Authors and Affiliations
Additional information
Accepted: 7 August 2000
Rights and permissions
About this article
Cite this article
Ponzano, E., Dondero, F., Bouquegneau, JM. et al. Purification and biochemical characterization of a cadmium metallothionein from the digestive gland of the Antarctic scallop Adamussium colbecki (Smith, 1902). Polar Biol 24, 147–153 (2001). https://doi.org/10.1007/s003000000186
Issue Date:
DOI: https://doi.org/10.1007/s003000000186