
Abstract
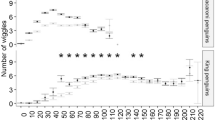
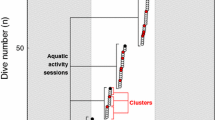
Diving behavior of 2 breeding Chinstrap penguins (Pygoscelis antarctica) was studied focusing first and primarily on dive bouts rather than dives themselves. Analysis of dive bout organization revealed (1) though there are differences between solitary dives and dive bouts in dive duration and dive depth, the first dives of dive bouts do not differ from solitary dives in the dive parameters, (2) mean dive duration during bout correlates positively to both mean dive depth during bout and mean surface interval during bout, while number of dives during bout negatively correlates to both cost (consumed energy) and duration of a dive cycle during bout. These findings suggest the following possibilities on foraging behavior of penguins: (1) their decision to repeat diving depends on the result of the first dive at a site, and the first dives of bouts would tend to be searching or evaluating dives though they would be also successful foraging dives, (2) they repeat diving at a foraging patch until foraging efficiency decrease to a threshold of diminishing returns.
Similar content being viewed by others

References
Bengtson, J.L., Croll, D.A., & Goebel, M.E. 1993 Diving behavior of Chinstrap penguins on Seal Island, Antarctica.Antarct. Sci. 5: 9–15.
Chappell, M.A., Shoemaker, V.H., Janes, D.N., Bucher, T.L. & Maloney, S.K. 1993a Diving behavior during foraging in breeding adelie penguins.Ecology 74: 1204–1215.
Chappell, M.A., Shoemaker, V.H., Janes, D.N., Maloney, S.K. & Bucher, T.L. 1993b Energetics of foraging in breeding adelie penguins.Ecology 74: 2450–2461.
Croll, D.A., Gaston, A.J., Burger, A.E. & Konnoff, D. 1992 Foraging behavior and physiological adaptation for diving in thick-billed murres.Ecology 73: 344–356.
Croxall, J.P., Davis, R.W., & O’Connel, M.J. 1988 Diving patterns in relation to diet of gentoo and macaroni penguins at South Georgia.Condor 90: 157–167.
Croxall, J.P., Everson, I., Kooyman, G.L., Ricketts, C., & Davis, R.W. 1985 Fur seal diving behavior in relation to krill vertical distribution.J. Anim. Ecol. 54: 1–8.
Culik, B. & Wilson, R.P. 1991 Energetics of under-water swimming in Adelie penguins (Pygoscelis adeliae).J. Comp. Physiol. B 161: 285–291.
Culik, B., Wilson, R.P., Dannfeld, R., Adelung, D., Spairani, H.J., & Coria, N.R. 1991 Pygoscelid penguins in a swim canal.Polar Biol. 11: 277–282.
Houston, A.I. & Carbone, C. 1992 The optimal allocation of time during the diving cycle.Behav. Ecol. 3: 255–265.
Japanese Fishery Agency 1993 The Sixth Antarctic Ocean Survey Cruise of JFA R.V. Kaiyo-maru. (in Japanese)
Kooyman, G.L. & Davis, R.W. 1987 Diving behavior and performance, with special reference to penguins. InSeabirds: feeding ecology and role in marine ecosystems (ed. J.P. Croxall), p.63–75 Cambridge University Press, Cambridge.
Kooyman, G.L., Cherel, Y., Le Maho, Y., Croxall, J.P., Thorson, P.H., Ridoux, V. and Kooyman, C.A. 1992 Diving behavior and energetics during foraging cycles in king penguins.Ecol. Monogr. 62: 143–163.
Kooyman, G.L. & Croll, D.A. 1987 Feeding patterns of emperor penguins.Antarct. J. 22: 221.
Kooyman, G.L. 1989 Diverse Divers: Physiology and behavior. Springer-Verlag, Berlin.
Kooyman, G.L., Davis, R.W., Croxall, J.P., & Costa, D.P. 1982 Diving depth and energy requirements of king penguins.Science 217: 726–727.
Krammer, D.L. 1987 The behavioral ecology of air breathing by aquatic animals.Can. J. Zool. 66: 89–94.
Lishman, G.S. & Croxall, J.P. 1983 Diving depth of the Chinstrap penguinPygoscelis antarctica.Bull. Br. Antarct. Surv. 61: 21–25.
Martin, P. & Bateson, P. 1993 Measuring Behaviour. 2nd ed. Cambridge University Press. Cambridge.
Naito, Y., Asaga, T., & Ohyama, Y. 1990 Diving behavior of Adélie penguins determined by time-depth recorders.Condor 92: 582–586.
Sibly, R.M., Nott, H.M.R. & Fletcher, D.J. 1990 Splitting behavior into bouts.Anim. Behav. 39: 63–69.
Stephens, D.W. & Krebs, J.B. 1986 Foraging Theory. Princeton University Press, Princeton.
Watanuki, Y., Kato, A., Mori, Y., & Naito, Y. 1993 Diving performance of Adélie penguins in relation to food availability in fast sea-ice area: comparison between years.J. Anim. Ecol. 62: 634–646.
Williams, T.D., Briggs, D.R., Croxall, J.P., Naito, Y., & Kato, A. 1992 Diving pattern and performance in relation to foraging ecology in the gentoo penguins (Pygoscelis papua).J. Zool., London 227: 211–230.
Wilson, R.P. 1984 An improved stomach pump for penguins and other sea birds.J. Field Ornithol. 55: 109–112.
Author information
Authors and Affiliations
About this article
Cite this article
Mori, Y. Dive bout organization in the Chinstrap penguin at seal island, antarctica. J Ethol 15, 9–15 (1997). https://doi.org/10.1007/BF02767321
Received:
Accepted:
Issue Date:
DOI: https://doi.org/10.1007/BF02767321