
Summary
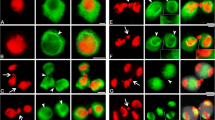
Photomicrographic evidence is presented of the difference in behavior of nuclear membranes during mitosis in amoebae, zygotes and plasmodia of Myxomycetes. One of the species was cultured on bacteria and possessed a normal cycle of plasmogamy and karyogamy between the amoebal and plasmodial phases. The second species was axenically grown in liquid media and had become highly heteroploid and lacked the ability to develop into plasmodia, existing only in the amoeboid form. The significance of the amoeboid form of mitosis in the heteroploid axenically cultured strain is discussed in relation to the difference in nuclear membrane behavior and the possible significance of such behavior.
Similar content being viewed by others


References
Aldrich, H., 1967: The ultrastructure of meiosis in three species ofPhysarum. Mycologia59, 127–148.
Alexopoulos, C. J., 1966: Morphogenesis in the Myxomycetes. In: The Fungi, Vol. II, 211–234, eds.G. C. Ainsworth andA. S. Sussman. New York: Academic Press.
Cadman, E., 1932: The life history and cytology ofDidymium nigripes Fr. Trans. Roy. Soc. Edinburgh57, 93–143.
Elliot, A. M., 1963: The fine structure ofTetrahymena pyriformis during mitosis. In: The Cell in Mitosis, ed.L. Levine. New York: Academic Press.
Godwood, M. B. C., 1966: The chromosomes of algae. New York: St. Martin Press.
Kerr, N. S., 1967: Plasmodium formation by a minute mutant of the true slime moldDidymium nigripes. Exp. Cell Res.45, 646–655.
Kerr, S. J., 1967: A comparative study of mitosis in amoebae and plasmodia of the true slime moldDidymium nigripes. J. Protozool.14, 439–445.
Koevenig, J. L., 1964: Studies on the life cycle ofPhysarum gyrosum and other Myxomycetes. Mycologia56, 170–184.
Mazia, D., 1964: Mitosis and the physiology of cell division. In: The Cell, Vol. III, eds.J. Brachet andA. E. Mirsky. New York: Academic Press.
Motta, J. J., 1967: A note on the mitotic apparatus in the rhizomorph meristem ofArmillaria mellea. Mycologia59, 370–375.
Robinow, C. F., andA. Bakerspigel, 1965: Somatic nuclei and the form of mitosis in fungi. In: The Fungi, Vol. I, eds.G. C. Ainsworth andA. S. Sussman, New York: Academic Press.
Ross, I. K., 1966: Chromosome numbers in pure and gross cultures of Myxomycetes. Amer. J. Bot.53, 712–718.
—, 1967 a: Abnormal cell behavior in the heterothallic myxomyceteDidymium iridis. Mycologia59, 235–245.
—, 1967 b: Growth and development of the MyxomycetePerichaena vermicularis. I. Cultivation and vegetative nuclear divisions. Amer. J. Bot.54, 617–625.
—, 1967 c: Syngamy and plasmodium formation in the MyxomyceteDidymium iridis. Protoplasma64, 104–119.
Schuster, F. L., 1965: Ultrastructure and morphogenesis of solitary stages of true slime molds. Protistologica1, 49–57.
Author information
Authors and Affiliations
Rights and permissions
About this article
Cite this article
Ross, I.K. Nuclear membrane behavior during mitosis in normal and heteroploid Myxomycetes. Protoplasma 66, 173–184 (1968). https://doi.org/10.1007/BF01252531
Received:
Issue Date:
DOI: https://doi.org/10.1007/BF01252531