
Abstract
The karyotypes of the toad Bufo marinus L. (2n=22) and the frogs Limnodynastes tasmaniensis Gthr. (2n=24), Rana temporaria L., R. esculenta L. (both 2n=26) and R. arvalis Nills. (2n=24) were analysed in colchicine treated leukocyte and spermatogonial metaphases and/or embryonic and larval mitoses. The DNA content of Feulgen stained erythrocyte nuclei was measured microspectrophotometrically. Heteromorphic sex chromosomes are absent in all species. L. tasmaniensis has the lowest DNA content among these species. The south American toad B. marinus shows a karyotype similar to the other known toad species and contains the same amount of DNA as the European species B. calamita with the lowest DNA amount among the European toads. In southern German populations of R. temporaria besides animals with the “standard”-karyotype (2n=26) individuals with 1 or 2, in rare cases with 3 or 4 supernumerary chromosomes have been found. The supernumeraries are heterochromatic and smaller than the smallest chromosome of the “standard”-karyotype. If only 1 or 2 supernumerary chromosomes are present, they seem to show normal mendelian inheritance as a rule. The observation of a few tadpoles with intraindividual different numbers of supernumeraries points to the occurrence of unequal distribution of these chromosomes in individuals containing a higher number of supernumerary chromosomes. The karyotype of R. esculenta is very similar to the “standard”-karyotype of R. temporaria, but the chromosomes of R. esculenta are somewhat longer than those of R. temporaria. R. esculenta contains about 54% more DNA than R. temporaria in the erythrocyte nuclei, so that it must be assumed that all chromosomes of R. esculenta contain more DNA than their homologues in R. temporaria. R. arvalis possesses about 28% more DNA than R. temporaria. It is supposed that these interspecific differences in DNA content of the Rana species — as observed earlier in Bufo species — are not a consequence of differential polyteny but are caused during evolutionary processes by local increase in DNA in the chromosomes of R. esculenta and R. arvalis.
Zusammenfassung
-
1.
An Colchicin-behandelten Mitosen von Leukocyten, Spermatogonien und Zellen embryonaler bzw. larvaler Gewebe wurden die Karyotypen der südamerikanischen Riesenkröte Bufo marinus, der australischen Froschart Limnodynastes tasmaniensis und der einheimischen Froschspecies Rana temporaria, R. esculenta und R. arvalis untersucht. Der DNS-Gehalt dieser Arten wurde cytophotometrisch an Feulgengefärbten Erythrocytenkernen bestimmt.
-
2.
Allen untersuchten Arten ist das Fehlen heteromorpher Geschlechtschromosomen gemeinsam.
-
3.
B. marinus (2n=22) besitzt einen ähnlichen Karyotyp wie die bisher bekannten Bufo-Arten und weist einen gleich hohen Gehalt an DNS auf wie die mitteleuropäische Art mit dem geringsten DNS-Gehalt, B. calamita.
-
4.
L. tasmaniensis besitzt 2n=24 größtenteils identifizierbare Chromosomen und ist vor den anderen Arten durch einen wesentlich geringeren DNS-Gehalt ausgezeichnet.
-
5.
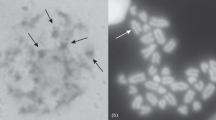
Der „Standard“-Karyotyp von R. temporaria weist 2n=26 fast vollzählig identifizierbare Chromosomen auf. Neben Tieren des „Standard“-Karyotyps treten bei R. temporaria in der gleichen Population jedoch in größerer Anzahl auch Individuen mit 1 oder 2, in seltenen Fällen auch mit 3 oder 4 überzähligen (B-)Chromosomen auf; diese zeichnen sich vor den anderen Chromosomen durch ihre geringe Größe aus, sind heterochromatisch und werden in der Regel normal mendelnd vererbt. In wenigen Fällen beobachtete „Mosaik“-Tiere weisen auf das Vorkommen ungleichmäßiger Verteilungen der Überzähligen in den Mitosen bei Tieren mit mehreren B-Chromosomen hin.
-
6.
Der Karyotyp von R. esculenta (2n=26) ist dem „Standard“-Karyotyp von R. temporaria sehr ähnlich, so daß sich alle Chromosomen beider Arten miteinander homologisieren lassen. R. esculenta besitzt jedoch einen um etwa 54% höheren DNS-Gehalt als R. temporaria. Da die Chromosomen von R. esculenta allgemein etwas größer als die ihnen homologen Chromosomen bei R. temporaria erscheinen, ist anzunehmen, daß alle oder fast alle Chromosomen von R. esculenta mehr DNS enthalten als ihre Homologen bei R. temporaria.
-
7.
R. arvalis besitzt mit 2n=24 Chromosomen einen offenbar durch Translokationsvorgänge während der Artdifferenzierung der Rana-Species abgewandelten Karyotyp. Hinsichtlich des DNS-Gehalts nimmt R. arvalis mit einem gegenüber R. temporaria um 28% höheren Gehalt an DNS eine Mittelstellung zwischen R. temporaria und R. esculenta ein.
-
8.
Die bestehenden interspezifischen Unterschiede im DNS-Gehalt der Rana-Arten werden wie bei den kürzlich analysierten Bufo-Arten (Ullerich, 1965, 1966) auf der Grundlage lokaler DNS-Zunahme in den Chromosomen während der Artdifferenzierung diskutiert.
Similar content being viewed by others


Literatur
Bianchi, N. O., and R. Laguens: Somatic chromosomes of Bufo arenarum. Cytologia (Tokyo) 29, 151–154 (1964).
Di Berardino, M. A.: The karyotype of Rana pipiens and investigation of its stability during embryonic differentiation. Develop. Biol. 5, 101–126 (1962).
Gall, J. G.: Kinetics of deoxyribonuclease action on chromosomes. Nature (Lond.) 198, 36–38 (1963); - Chromosomes and cytodifferentiation. In: Cytodifferential and macromolecular synthesis. New York: Academic Press 1963.
Hennen, S.: The karyotype of Rana sylvatica and its comparison with the karyotype of Rana pipiens. J. Heredity 55, 124–128 (1964).
Hertwig, G.: Kern- und Zellgrößenunterschiede der Eltern als Ursache des verschiedenen Ausfalls reziproker Krötenbastarde. Z. Anat. 92, 718–739 (1930); - Hochgradige Polynemie der Chromosomen als Ursache der vergleichsweise erheblichen Zell- und Kerngrößen der Amphibien. Verh. anat. Ges. (Jena) 53, 104–112 (1956).
Keyl, H.-G.: Verdopplung des DNS-Gehalts kleiner Chromosomenabschnitte als Faktor der Evolution. Naturwissenschaften 51, 46–47 (1964); - A demonstrable local and geometric increase in the chromosomal DNA of Chironomus. Experientia (Basel) 21, 191–193 (1965a); - Duplikationen von Untereinheiten der chromosomalen DNS während der Evolution von Chironomus thummi. Chromosoma (Berl.) 17, 139–180 (1965b).
La Cour, L. F., and S. R. Pelc: Effect of colchicine on the utilization of labelled thymidine during chromosomal reproduction. Nature (Lond.) 182, 506–508 (1958).
Makino, S.: Notes on the chromosomes of Rana temporaria L. and Bufo sachalinensis (Nikolski). Proc. Imp. Acad. 8, 23–28 (1932).
Melander, Y.: Accessory chromosomes in animals, specially in Polycelis tennis. Hereditas (Lund) 36, 19–37 (1950).
Mikamo, K., and E. Witschi: The mitotic chromosomes in Xenopus laevis (Daudin): normal, sex reversed and female WW. Cytogenetics 5, 1–19 (1966).
Miller, O. L.: Fine structure of lampbrush chromosomes. Nat. Cancer Inst. Monogr. 18, 79–99 (1965).
Mirsky, A. E., and H. Ris: The deoxyribonucleic acid content of animal cells and its evolutionary significance. J. gen. Physiol. 34, 451–462 (1951).
Morescalchi, A.: Il corredo cromosomico del Bufonidi Italiani. Bull. Zool. 31, 827–836 (1964).
Peacock, W. J.: Chromosome duplication and structure as determined by autoradiography. Proc. nat. Acad. Sci. (Wash.) 49, 793–801 (1963).
Prescott, D. M., and M. A. Bender: Autoradiographic study of chromatid distribution of labelled DNA in two types of mammalian cells in vitro. Exp. Cell Res. 29, 430–442 (1963).
Rothfels, K., E. Sexsmith, M. Heimburger, and M. O. Krause: Chromosome size and DNA content of species of Anemone L. and related genera (Ranunculaceae). Chromosoma (Berl.) 20, 54–74 (1966).
Seto, T.: Cytogenetic studies in lower vertebrates. II. Karyological studies of several species of frogs (Ranidae). Cytologia (Tokyo) 30, 437–446 (1965).
Taylor, J. H.: Sister chromatid exchange in tritium labelled chromosomes. Genetics 43, 515–529 (1958); - Chromosome reproduction. Int. Rev. Cytol. 13, 39–73 (1962); - The replication and organization of DNA in chromosomes. In: Molecular genetics, p. 65–111, ed. by J. H. Taylor. New York: Academic Press 1963.
—, P. S. Woods, and W. L. Hughes: The organization and duplication of chromosomes as revealed by autoradiographic studies using tritium labelled thymidine. Proc. nat. Acad. Sci. (Wash.) 43, 122–128 (1957).
Trosko, J. E., and S. Wolff: Strandedness of Vicia faba chromosomes as revealed by enzyme digestion studies. J. Cell Biol. 26, 125–135 (1965).
Uhl, C. H.: Chromosome structure and crossing over. Genetics 51, 191–207 (1965).
Ullerich, F.-H.: Unterschiede im DNS-Gehalt der Genome von Bufo bufo und Bufo viridis. Z. Naturforsch. 20b, 720–722 (1965); - Karyotyp und DNS-Gehalt von Bufo bufo, B. viridis, B. bufo x B. viridis und B. calamita (Amphibia, Anura). Chromosoma (Berl.) 18, 316–342 (1966).
Weiler, C., and S. Ohno: Cytological confirmation of female heterogamety in the african water frog (Xenopus laevis). Cytogenetics 1, 217–223 (1962).
White, M. J. D.: Animal cytology and evolution, 2nd ed. Cambridge: Cambridge University Press 1954.
Wickbom, T.: Cytological studies on Dipnoi, Urodela, Anura, and Emys. Hereditas (Lund) 31, 241–346 (1945).
Witschi, E.: Die Entwicklung der Keimzellen der Rana temporaria L. I. Urkeimzellen und Spermatogenese. Z. Zellen- und Gewebelehre 1, 523–561 (1924).
—, M. Kodani, and K. Mikamo: Comparative study of the chromosomes of European, American, and Japanese frogs. Anat. Rec. 131, 610 (1958).
Wolf, K., and M. C. Quimby: Amphibian cell culture: permanent cell line from the bullfrog (Rana catesbeiana). Science 144, 1578–1580 (1964).
Woods, P. S., and M. U. Schairer: Distribution of newly synthesized deoxyribonucleic acid in dividing chromosomes. Nature (Lond.) 183, 303–305 (1959).
Yosida, T. H.: Sex chromosomes of the tree frog, Hyla arborea japonica. J. Fac. Sci. (Ser. 6), Hokkaido Univ. 13, 352–358 (1957).
Author information
Authors and Affiliations
Rights and permissions
About this article
Cite this article
Ullerich, FH. Weitere Untersuchungen über Chromosomenverhältnisse und DNS-Gehalt bei Anuren (Amphibia). Chromosoma 21, 345–368 (1967). https://doi.org/10.1007/BF00336947
Received:
Issue Date:
DOI: https://doi.org/10.1007/BF00336947