
Abstract
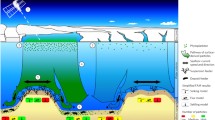
The sedimentation pattern of organic material in the Greenland-Norwegian Sea is reflected in the surface sediments, although less than 0.5% of the organic matter is buried in the sediment. Maximum fluxes and benthic responses are observed during June and/or August/September, following the pattern of export production in the pelagial zone. The annual remineralization rate on the Vøring Plateau is 3.0 g C m−2 a −1 Freshly settled phytodetritus, as detected by chlorophyll measurements, is rapidly mixed into the sediment and decomposed. It stimulates the activity of benthic organisms, especially foraminifera. The mixing coefficient for this material is D b=0.2 cm2 d−1, which is two to three orders of magnitude higher than that estimated from radiotracer methods. The effect on the geological record, however, is likely to be small. Chlorophyll-containing particles are at first very evenly distributed on the seafloor. After partial decomposition and resuspension, a secondary redistribution of particles occurs which can result in the formation of a high accumulation area, with an up to 80-fold increase in the sedimentation rate by lateral advection. This is mainly due to physical processes, because biodeposition mediated by benthic animals increases sedimentation by only a factor of two or three.
Similar content being viewed by others


References
Abele-Oeschger D (1991) Potential of some carotenoids in two recent sediments of Kiel Bight as biogenic indicators of phytodetritus. Mar Ecol Prog Ser 70:83–92
Altenbach AV (1992) Short term processes and patterns in the foraminiferal response to organic flux rates. Mar Micropalaeontol 19:119–129
Bacon MP, Loeff MMRvd (1989) Removal of thorium-234 by scavenging in the bottom nepheloid layer of the ocean. Earth Planet Sci Lett 92:157–164
Barnett PRO, Watson J, Conelly D (1984) A multiple corer for taking virtually undisturbed samples from shelf, bathyal, and abyssal sediments. Oceanol Acta 7:399–408
Billett DSM, Lampitt RS, Rice ALM, Mantoura RFC (1983) Seasonal sedimentation of phytoplankton to the deep-sea benthos. Nature 302:520–522
Blaume F (1992) Hochakkumulationsgebiete am norwegischen Kontinentalhang: Sedimentologische Abbilder Topographiegeführter Strömungsmuster. Ber SFB 313, Univ Kiel No 36:1–150
Bodungen B von, Theilen F, Werner F (1988) Poseidon Reise 141–2/142 vom 17. 10. 1987–18. 11. 1987, Poseidon-Reise 146–1 vom 25. 4.-11. 5. 1988, Poseidon-Reise 146–3 vom 29. 5.-19. 6. 1988, Berichte der Fahrtleiter. Ber SFB 313 Univ Kiel No 11:1–66
Bodungen B von, Voss M, Wunsch M (1991) Vertical paerticle flux in the Norwegian Sea. In: Wassmann P, Heiskanen AS, Lindahl O (eds) Sediment Trap Studies in the Nordic Countries 2. Nurmi Print, Nurmijärvi, pp 116–136
Bodungen B von, Antia A, Bauerfeind E, Haupt O, Koeve W, Peeken I, Peinert R, Reitmeier S, Thomsen C, Voss M, Wunsch M, Zeller U, Zeitzschel B. Pelagic processes and vertical flux of particles: an overview over a long-term comparative study in the Norwegian Sea and Greenland Sea. Geol Rundsch 84: 11–27
Boudreau BP (1986a) Mathematics of tracer mixing in sediments: I. spatially-dependant, diffusive mixing. Am J Sci 286:161–198
Boudreau BP (1986b) Mathematics of tracer mixing in sediments: II. nonlocal mixing and biological conveyor-belt phenomena. Am J Sci 286:199–238
Boudreau BP (1987) Mathematics of tracer mixing in sediments: III. The theory of nonlocal mixing within sediments. Am J Sci 287:693–719
Brandt A (1993) Composition, abundance, and diversity of peracarid crustaceans on a transect of the Kolbeinsey Ridge, north of Iceland. Polar Biol 13:565–576
Cochran JK (1985) Particle mixing rates in sediments of the eastern equatorial Pacific: evidence from 210Pb, 239,240Pu and 137Cs distributions at MANOP sites. Geochim Cosmochim Acta 49:1195–1210
Erlenkeuser H, Balzer W (1988) Rapid appearance of Chernobyl radiocesium in the deep Norwegian Sea sediments. Oceanol Acta 11:101–106
Gardner WD, Southard JB, Hollister CD (1985) Sedimentation, resuspension and chemistry of particles in the Northwest Atlantic. Mar Geol 65:199–242
Gerlach SA, Graf G (1991) Europaisches Nordmeer, Reise Nr. 13, 6. Juli-24. August 1990. Meteor-Ber Univ Hamburg No 91–2:1–217
Gerlach SA, Thiede J, Graf G, Werner F (1986) Forschungsschiff Meteor, Reise 2 vom 19. Juni bis 16. Juli 1986, Forschungsschiff Poseidon, Reise 128 vom 7. Mai bis 8. Juni 1986. Ber SFB 313 Univ Kiel No 4:1–140
Gerlach SA, Theilen F, Werner F (1987) Forschungsschiff Poseidon, Reise 119 vom 16. Juli bis 1. August 1985. Forschungsschiff Poseidon, Reise 120 vom 4. August bis 20. August 1985. Forschungsschiff Valdivia, Reise 201–48A vom 17. Juli bis 31. Juli 1986. Forschungsschiff Poseidon, Reise 137 vom 3. Februar bis 20. Februar 1987. Ber SFB 313, Univ Kiel No 5:1–94
Graf G (1989a) Benthic-pelagic coupling in a deep-sea benthic community. Nature 341:437–439
Graf G (1989b) Die Reaktionen des Benthals auf den saisonalen Partikelfluß und die laterale Advektion, Bowie deren Bedeutung für Sauerstoff- und Kohlenstoffbilanzen. Ber SFB 313 Univ Kiel: 1–77
Graf G (1992) Benthic-pelagic coupling: a benthic view. Oceanogr Mar Biol Annu Rev 30:149–190
Graf G, Linke P (1992) Adenosine nucleotides as indicators of deep-sea benthic metabolism. In: Rowe GT, Pariente V (eds) Deep-Sea Food Chains and the Global Carbon Cycle. Kluwer Academic, Dordrecht, Boston, London, pp 237–243
Grasshoff K (1976) Methods of Sea Water Analysis. Verlag Chemie, Weinheim, pp 1–317
Grebmeier JM, Barry JP (1991) The influence of oceanographic processes on pelagic-benthic coupling in polar regions: a benthic perspective. J Mar Syst 2:495–518
Hecker B (1990) Photographic evidence for the rapid flux of particles to the seafloor and their transport down the continental slope. Deep-Sea Res 37:1773–1782
Heeger T (1990) Elektronenmikroskopische Untersuchungen zur Ernährungsbiologie benthischer Foraminiferen. Ber SFB 313 Univ Kiel No 21:1–139
Hirschleber H, Theilen F, Balzer W, Bodungen B von, Thiede J (1988) Forschungsschiff Meteor, Reise 7 vom 1. Juni bis 28. September 1988. Ber SFB 313 Univ Kiel No 10:1–358
Jeffrey SW, Humphrey GF (1975) New spectrophotometric equations for determining chlorophylls a, b, c1, and c2 in higher plants, algae and natural phytoplankton. Biochem Physiol Pflanzen (BPP) p 167
Jensen P (1992a) ‘An Enteropneust's Nest’: results of the burrowing traits by the deep-sea acorn worm Stereobalanus canadensis (Spengel). Sarsia 77:125–129
Jensen P (1992b) Cerianthus vogti Danielssen, 1890 (Anthozoa: Ceriantharia). A species inhabiting an extended tube system deeply buried in deep-sea sediments off Norway. Sarsia 77:75–80
Jensen P, Rumohr J, Graf G (1992) Sedimentological and biological differences across a deep-sea ridge exposed to advection and accumulation of fine-grained particles. Oceanol Acta 15:287–296
Köster M, Charfreitag O, Meyer-Reil L-A (1991) Availability of nutrients to a deep-sea benthic microbial community: results from a ship-board experiment. Kiel Meeresforsch Sonderh 8:127–133
Lampitt RS (1985) Evidence for seasonal deposition of detritus to the deep-seafloor and its subsequent resuspension. Deep-Sea Res 32:885–897
Linke P (1992) Metabolic adaptations of deep-sea benthic foraminifera to seasonally varying food input. Mar Ecol Prog Ser 81:51–63
Linke P, Lutze GF (1993) Microhabitat preferences of benthic foraminifera — a static concept or a dynamic adaptation to optimize food acquisition? Mar Micropaleontol 20:215–234
Loo L-O, Rosenberg R (1989) Bivalve suspension-feeding dynamics and benthic-pelagic coupling in an eutrophicated marine bay. J Exp Mar Biol Ecol 130:253–276
Lutze GF, Altenbach A (1988) Rupertina stabilis (Wallich), a highly adapted, suspension feeding foraminifer. Meyniana 40:55–69
Meyer-Reil L-A, Koster M (1992) Microbial life in pelagic sediments: the impact of environmental parameters on enzymatic degradation of organic material. Mar Ecol Prog Ser 81:65–72
Muschenheim DK (1987) The role of hydrodynamic sorting of seston in the nutrition of a benthic suspension feeder, Spio setosa (Polychaeta: Spiobnidae). Biol Oceanogr 4:265–288
Peinert R, Bathmann U, Bodungen Bv, Noji T (1987) The impact of grazing on spring phytoplankton growth and sedimentation in the Norwegian current. Mitt Geol-Paläontol Inst Univ Hamburg Sb 62:149–164
Pfannkuche O (1993a) Benthic response to the sedimentation of particulate organic matter at the BIOTRANS station, 47°N 20°W. Deep-Sea Res 40:135–149
Pfannkuche O (1993b) Standing stock, Aktivität und Respirationsraten des Tiefseebenthos im NO-Atlantik. In: Pfannkuche O, Rheinheimer G, Thiel H (eds) BIO-C-FLUX Biologischer Kohlenstofffluß in der bodennahen Wasserschicht des küsten-fernen Ozeans. Ber IFM Kiel Nr 242:51–79
Pfannkuche O, Duinker JC, Graf G, Henrich R, Thiel H, Zeitschel B (1993) Nordatlantik 92, Reise Nr. 21, 16. März-31. August 1992. Meteor-Ber Univ Hamburg No 93–4:1–281
Romero-Wetzel MB (1987) Sipunculans as inhabitants of very deep, narrow burrows in deep-sea sediments. Mar Biol 96:87–91
Romero-Wetzel MB (1989) Branched burrow-systems of the enteropneust Stereobalanus canadensis (Spengel) in deep-sea sediments of the Vøring Plateau, Norwegian Sea. Sarsia 74:85–89
Romero-Wetzel MB, Gerlach SA (1991) Abundance, biomass, size-distribution and bioturbation potential of deep-sea macrozoobenthos on the Vøring Plateau (1200–1500 m, Norwegian Sea). Meeresforsch 33:247–265
Sarnthein M, Altenbach A. Late quaternary changes in surface water and deepwater masses of the nordic seas and northeastern North-Atlantic — a review. Geol Rundsch 84: 89–107
Smith CR, Jumars PA, DeMaster DJ (1986) In situ studies of megafaunal mounds indicate rapid sediment turnover and community response at the deep-seafloor. Nature 323:251–253
Smith CR, Pope RH, Demaster DJ, Magaard L (1993) Age-dependent mixing of deep-sea sediments. Geochim Cosmochim Acta 57:1473–1488
Struck U (1992) Zur Paläo-Ökologie benthischer Foraminiferen im Europäischen Nordmeer während der letzten 600000 Jahre. Ber SFB 313 Univ Kiel No 38:1–89
Suess E (1980) Particulate organic carbon flux in the oceans-surface productivity and oxygen utilization. Nature 288:260–263
Suess E, Altenbach AV (1992) Europaisches Nordmeer, Reise Nr. 17, 15. Juli-29. August 1991. Meteor-Ber Univ Hamburg No 92–3:1–164
Sun M, Aller RC, Lee C (1991) Early diagenesis of chlorophyll-a in Long Island Sound sediments: a measure of carbon flux and particle reworking. J Mar Res 49:379–401
Thiel H, Pfannkuche O, Schriever G, Lochte K, Gooday AJ, Hemleben C, Mantoura RFG, Turley CM, Patching JW, Riemann F (1988/1989) Phytodetritus on the deep-sea floor in a central oceanic region of the Northeast Atlantic. Biol Oceanogr 6:203–239
Thomsen L (1992) Untersuchungen zur Bodennepheloidschicht am westlichen Barents See Kontinentalhang. Ber SFB 313 Univ Kiel No 42:1–93
Thomsen L, Graf G. Characteristics of suspended particulate matter in the benthic boundary layer of the continental margin of the western Barents Sea. Oceanol Acta, submitted
Thomsen L, Graf G, Martens V, Steen E (1994) An instrument for sampling water from the benthic boundary layer. Cont Shelf Res 14:871–882
Thomsen L, Graf G, Juterzenka K von, Witte U. An in situ experiment to investigate the depletion of seston above an interface feeder field on the continental slope of the western Barents Sea. Mar Ecol Prog Ser, submitted
Wagner T (1993) Organisches Material in pelagischen Sedimenten: Glaziale/Interglaziale Variationen im Europäischen Nordmeer. Ber SFB 313 Univ Kiel No 42:1–138
Zeitzschel B, Lenz J, Thiel H, Boje R, Passow U, Stuhr A (1990) Expedition Plankton 89 — Benthos 89, Reise Nr. 10, 19. März-31. August 1989. Meteor-Ber Univ Hamburg No 90–1:1–216
Author information
Authors and Affiliations
Rights and permissions
About this article
Cite this article
Graf, G., Gerlach, S.A., Linke, P. et al. Benthic-pelagic coupling in the Greenland-Norwegian Sea and its effect on the geological record. Geol Rundsch 84, 49–58 (1995). https://doi.org/10.1007/BF00192241
Received:
Accepted:
Issue Date:
DOI: https://doi.org/10.1007/BF00192241