
Abstract
Over the past years, biodiversity has been reduced on an unprecedented scale, while new infectious diseases are emerging at an increasing rate. Greater overall biodiversity could lead to a greater diversity of hosts and thus of pathogens. Yet disease regulation – due to the buffering role of host diversity – is considered to be one of the services provided by biodiversity. In this chapter, we ask how biodiversity is linked to infectious disease. First, we investigate the influence of the biodiversity of pathogens. We highlight that the number of pathogen species is not well known but that new findings are facilitated by the rapid expansion of molecular techniques. We show that, although there is a trend to find higher pathogen richness toward the equator, identifying a global pattern between the richness of all pathogen species and their latitudinal distribution is challenging. We emphasize that pathogen intraspecific diversity is a crucial factor in disease emergence and allows pathogens to adapt to the selective pressures they face. In addition, the selective pressure acting on hosts due to parasite, and reinforced by parasite diversity within hosts seems to be a major evolutionary and ecological force shaping hosts biodiversity. Second, we investigate how the diversity of hosts influences infectious disease ecology. For multi-host diseases, a change in host species richness or abundance can modify the dynamics of local infectious diseases by either reducing (“dilution effect”) or increasing (“amplification effect”) the risk of transmission to the targeted host species. The underlying hypothesis is that, the competence of reservoirs varies according to the host species. The dilution effect has been demonstrated mainly through theoretical work and there have been only few case studies. Regarding the genetic diversity of host, an important issue is: to what extent does a reduction of this diversity impact the ability of the host population to response to infectious diseases? Third, we rapidly examine the role of biodiversity in the treatment of infectious diseases. To conclude, we consider that the consequences of the loss of species biodiversity on infectious diseases is still largely unknown, notably due to the lack of knowledge on the dynamics of host-pathogen relationships, especially at the population and at the community level.. We highlight that work on multi-host/ ulti-pathogen systems should be fostered and that new approaches, such as metagenomic investigations that does not require a priori assumptions, are promising to describe a community of pathogens and their interactions.
You have full access to this open access chapter, Download chapter PDF
Similar content being viewed by others


Keywords
- Major Histocompatibility Complex
- West Nile Virus
- Lyme Disease
- Pathogen Species
- Major Histocompatibility Complex Gene
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
1 Introduction
Over the past 50 years, human activity has altered habitats and reduced biodiversity on an unprecedented scale that comes close to mass extinction (MEA 2005). At the same time, new infectious diseases seem to be emerging at an increasing rate (Wilcox and Gubler 2005). During this period, there has been a dramatic spread of highly pathogenic diseases such as AIDS and multi-drug resistant bacterial infections, and in more recent years SARS, West Nile in North America, and highly pathogenic influenza viruses (Jones et al. 2008).
Habitat loss, largely a result of the conversion of forests and savannas into agricultural land, cities, and industrial sites, is the major cause of change in biodiversity. Biodiversity represents the diversity of life at all levels of biological organization, from the genes within populations, the species that compose a community, to the communities that compose ecosystems. Intuitively, one might assume that greater overall biodiversity would lead to a greater diversity of pathogens and hosts, and thereby increase the incidence of infectious diseases (Dunn et al. 2010). Yet disease regulation is said to be one of the services provided by biodiversity because a high level of species diversity creates a buffer that reduces the risk of transmission (MEA 2005; Walpole et al. 2009). Scientific evidence supporting both of these views is beginning to emerge, but the core question remains: how is biodiversity linked to infectious disease? This is the question addressed in this chapter.
Pathogens are organisms that have a negative impact on the fitness of their host(s), often, if now always, by producing visible symptoms (e.g. a disease). Such trophic interaction between two organisms, a host and a parasite, is just one of several interactions that take place within communities and ecosystems, others being those of prey-predator and plant-phytophagous for instance (Begon et al. 2006). To date, more attention has been paid to these other interactions, and their roles in ecosystem functioning (e.g. Steffan and Snyder 2010), than to pathogen-host interactions, and food web studies only recently have begun to take parasites into consideration (Arias-González and Morand 2007; Lafferty et al. 2006). Studies incorporating pathogens are scarce (Hudson et al. 2006), probably due to the difficulties of surveying pathogens (using intrusive or even destructive sampling methods…). Moreover, the systematics and even basic aspects of parasite biology often are unknown. However, although numerous species of pathogens still need to be described (Dobson et al. 2008), there is no doubt that pathogens represent a large part of biodiversity on earth. Given that each free living species is host to numerous pathogens, and that pathogens of pathogens also exist (consider, for example, phages that are virus affecting bacteria), several authors believe that pathogens may be the most diverse living group on earth (Windsor 1998).
The link between biodiversity and the ecology of infectious diseases is not simple. In this chapter, we investigate how biodiversity influences the ecology of infectious diseases at the intraspecific level (genetic variability of pathogens and hosts) and at the level of communities (species composition). Although we mainly provide examples from human and animal diseases, we also use some illustrations from plants. We describe patterns of biodiversity and the consequences of changes in biodiversity on the ecology of infectious diseases. Lastly, we rapidly examine the role of biodiversity in the treatment of infectious diseases.
2 How Does the Diversity of Pathogens Influence Infectious Disease Ecology?
2.1 How Many Pathogens Are There?
We shall consider infectious diseases caused by bacteria, virus, fungi, protozoa and endo-parasites, and exclude from our analysis ecto-parasites that are considered here as disease vectors. In the light of the discussion above, the pathogen status of a given living organism clearly is not a straightforward question (consider, for example, the case of some Rickettsia species that are considered to be not only blood vertebrate pathogens, but also tick symbionts, Perlman et al. 2006). This status is related to the host, and varies with individual hosts and species as well as in space and in time (different hosts, for example, can have different resistance or susceptibility). When pathogens have complex life-cycles, some stages may have a different biology (such as biotrophic or necrotrophic plant pathogens, Morris et al. 2009). Furthermore, horizontal gene transfer is so extensive in bacteria that many microbiologists question the existence of species in bacteria, preferring to consider bacteria as populations that exchange genes. However, the existence of core genes responsible for the maintenance of species-specific phenotypic clusters is an argument supporting the identification of bacterial species (Riley and Lizotte-Waniewski 2009).
For these reasons, combined with the limited knowledge available of the systematics of many pathogens (Brooks and Hoberg 2001), it is difficult to accurately estimate the number of pathogen species. Estimations of pathogen species richness vary from 10% to 50% of living beings (de Meeûs et al. 1998; Poulin and Morand 2004). In estuaries, the biomass of macro and micro-parasites has been estimated as exceeding that of top predators (Kuris et al. 2008). Although the existence of pathogens has been known for a long time, lists of species only were compiled recently for human and animals (Ashford and Crewe 1998; Cleaveland et al. 2001; Taylor et al. 2001), with an update on human pathogens completed in 2007 (Woolhouse and Gaunt 2007). Approximately 1,400 human pathogens were reported, 616 livestock pathogens (cattle, sheep, goats, pigs and horses), and 374 domestic carnivore pathogens (dogs and cats). No clear figure was given for wildlife (but see the Global Mammal Parasite Database at http://www.mammalparasites.org/). On average, over two new species of human viruses also are discovered each year (Woolhouse et al. 2008). Pathogens affecting humans have received more attention than those affecting other species. If one assumes that other animal species are affected in a proportional manner, huge numbers of pathogens remain to be discovered. Altogether, 70,000, 5,000 and 4,000 of fungi, viruses and bacteria respectively have been described, which represent only 5%, 1%, and 0–1% of the total estimated number of species of fungal, viral and bacterial species. It is difficult to know the number of plant pathogens, but a significantly proportion of the fungal, viral and bacterial species are likely to be plant pathogens (Ingram 1999).
Until recently, many new pathogen discoveries relied on the investigation of atypical symptoms. Today, new findings are facilitated by molecular techniques that render it possible to detect and characterise unculturable pathogens and to investigate the presence of genes and genomes independently of individuals (metagenomics). Although multi-host pathogens are more numerous than single hosts, interactions between pathogens and hosts can evolve towards the specialisation of pathogens on a given host species (Cleaveland et al. 2001; Huyse et al. 2005; Pedersen et al. 2005). Such a specialisation can lead to speciation, id est the birth of a new pathogen species. Co-cladogenesis, a process of parallel diversification in hosts and pathogens, also can give birth to numerous pathogen species (Page 2003). Until the development of molecular tools, these species were very difficult to distinguish (cryptic species). Systematic investigations using molecular tools have made it possible, however, to reveal a high diversity of pathogens. For instance, in a systematic inventory of viruses in various vertebrate hosts conducted over a 20 year period in the Central African Republic, 919 different viruses were isolated, including 39 new ones (Saluzzo et al. 2004). Two species of Plasmodium, P. falciparum, infecting humans, and P. reichenowi, infecting chimpanzees, were long considered to be within the clade that includes humans and the great apes. However, recent studies of apes in their natural habitat have revealed a much higher diversity of species infecting great apes; in addition, it has been found that P. falciparum also infect gorillas (Liu et al. 2010; Prugnolle et al. 2010) and are at the origin of human malaria. Metagenomic studies in ecosystems such as human faeces (Zhang et al. 2006) and marine sediments (Breitbart et al. 2004) also have revealed that the majority of viral sequences found did not match in the databanks. Finally, new investigations have been launched to monitor people, animals and animal die-offs in areas where people have a high exposure to wildlife. Generic, broad screening tools will be used to detect pathogen species (Wolfe et al. 2007). To our knowledge, a similar approach has not yet been implemented for pathogens of animals or plants.
2.2 Does the Worldwide Distribution of Pathogen Species Diversity Mirror That of Other Organisms?
In addition to the inventories of pathogen biodiversity, scientists have investigated which part of the world holds the highest diversity of pathogen species. Many studies on plants and animals have shown that species richness decreases the further one moves away from the equator. The reasons most likely are linked to the area, energy, time and habitat heterogeneity, and geometric constraints (Gaston and Blackburn 2000). Comparative studies exploring pathogen species richness in the tropics compared to temperate zones are scarce and have produced discrepant results. Guernier et al (2004) studied the worldwide distribution of 229 human pathogens (bacteria, virus, fungi, protozoa, and helminths) according to environmental, demographic and economical factors. They found that parasite species richness decreased with latitude and had a spatially nested organization; i.e. some species are widely distributed and occur in many communities while others have more restricted distributions and occur only in a subset of locations. Such findings were confirmed by the analysis of Dunn et al. (2010), who showed that human pathogen diversity was strongly related to both mammal and bird species richness. Diseases that occur in temperate zones also tend to occur in the tropics, while some tropical diseases are restricted only to the tropics. In primates, the number of protozoa species, which primarily are vector-borne transmitted, increase as one approaches the equator, however, the same trend was not found for viruses and helminths (Nunn et al. 2005). Lindenfors et al (2007), who examined the parasite richness of 980 parasite species and 146 terrestrial carnivore species, found that helminth parasite species richness increased the further away one moved from the equator. The reason for this finding is unknown and may be related to a bias in sampling because carnivores inhabiting areas of industrialized countries in the Northern Hemisphere may have been sampled more intensely. Poulin (1995) and Bordes et al (2010) did not find any correlation between helminth species richness at intra or interspecific levels and latitude. Some studies have shown higher tick species richness at lower latitudes (Cumming 2000). However, this is not the case for flea species, which have been found to have low richness at lower latitudes (Krasnov et al. 2004). A final example is Ichneumonid parasitoid hymenoptera. Although a higher specific host diversity is found in the tropics, the number of species of this parasitoid group is similar in both tropical and temperate regions. It has been hypothesised that this is due to habitat fragmentation (leading to a lower density of hosts); lack of seasonality (and thus of a host population dynamics with peaks and high density of hosts), or the higher content in toxic compounds of tropical plants and thus in phytophagous insects (the “nasty hypothesis”) (Gauld et al. 1992). A meta-analysis of parasite-associated host mortality (Robar et al. 2010) revealed that host mortality risk declined as one moves away from the equator, indicating that, in addition to parasitic load, the effect of parasites on host mortality might be determined by some abiotic factors. Thus, although there is a trend to find higher pathogen richness as we move toward the equator, it is thus challenging to identify a global pattern between the richness of all pathogen species and their latitudinal distribution. However, it should be noted that of the 87 pathogens that have been discovered since 1980, most have a global distribution (Woolhouse and Gaunt 2007).
2.3 Pathogen Intraspecific Diversity Is One Factor Favouring Disease Emergence
Pathogens generally are characterised as having higher mutation rates and generation times than those of their hosts (Hamilton et al. 1990). Genetic variability also results from recombination during sexual reproduction of eukaryotic pathogens, and any other genetic exchange mechanisms such as bacterial conjugation or viral recombination. In addition, many animal and plant pathogens use a vector to increase gene flow among populations and to reach a new individual host. This genetic diversity is a crucial factor in disease emergence (Cleaveland et al. 2001) and allows pathogens to adapt to the main selective pressures they face: hosts’ immune systems, the need to be transmitted, and treatments or vaccines used to counter infections.
The capacity of some pathogens to genetically diversify facilitates their ability to evade host immune systems. One of the best examples is the Human Immunodeficiency Virus (HIV), which is able to change its appearance faster than the time its takes for the immune system to reply (Drosopoulos et al. 1998). Another example is P. falciparum, which generates high levels of variability in genes involved in antigenic variability and virulence (var genes) by producing frequent recombination events between heterologous chromosomes (Freitas-Junior et al. 2000). High genetic variation of pathogens also is involved in the interspecies infection process as it facilitates the infection of a broader range of host species, which is another characteristic of emerging pathogens (Cleaveland et al. 2001; Woolhouse and Gowtage-Sequeria 2005). The evolutionary potential of pathogens allows them to respond quickly to the directional selective pressure provided by the massive use of drugs (Palumbi 2001). In areas where selective pressure is important, such as in hospitals, multi-resistant bacteria are very frequent (Levy and Marshall 2004). For bacteria, resistant genes probably originated from environmental organisms with which they shared their ecological niche (Aminov and Mackie 2007). These genes can be transferred between different species of bacteria and even between species that colonize different hosts (Nikolich et al. 1994).
Although vaccination is a major advance of modern medicine, it thus far has contributed to the eradication of only one infectious disease in humans (small pox, www.who.int/mediacentre/factsheets/smallpox/en) and one in cattle, buffalo and wildebeest (rinderpest, Normile 2008). As many vaccines do not totally block transmission, vaccination modifies the selective pressure on pathogens. Depending on how vaccines act on the pathogen, the epidemiology consequences can differ (Gandon and Day 2007). For instance, theoretical work has shown that vaccines that reduce the growth rate or toxicity of pathogens also reduce selection pressure against virulent pathogens, leading to higher intrinsic virulence (Gandon et al. 2001). In the poultry industry, an increase in virulence of avian tumour viruses has followed the use of vaccines that reduce virus growth rates (Witter 1997).
2.4 Pathogen Diversity to Which Hosts Are Exposed Influences Host Susceptibility to Disease
Although plants lack an adaptive immune system, through evolution they have developed various strategies to stop plant pathogen infections. An induced or acquired systemic resistance occurs following host recognition of a pathogen, which triggers the production of a hypersensitive reaction (Jones and Dangl 2006). Through this mechanism, the plant provides itself protection from secondary infection in distal tissues, even if the plant faces a pathogen for which it does not have the resistance gene (Durrant and Dong 2004).
The immune system of vertebrates acquires its efficiency by being exposed to a diverse array of pathogens. The striking increase in hygiene standards that began in the early twentieth century has considerably lowered humans’ exposure to pathogens, at least in developed countries. The immune response triggered by a pathogen can provide some cross protection against other pathogens (e.g. Huang et al. 2008). A low exposure to a diversity of pathogens has had immediate consequences in decreasing the risk of disease. But could this low exposure also induce evolutionary change in the ability of a host to respond to infection? Due to a trade off between investment in disease resistance and other traits linked to fitness, low exposure could decrease the frequency of resistance down through the generations, setting the stage for a potentially devastating outbreak (Altizer et al. 2003; Graham et al. 2010). Domestic species that are bred to be protected from pathogens might be more susceptible to infectious diseases (Lyles and Dobson 1993). Furthermore, it has been suggested that on islands, where some pathogens may be absent, hosts may have lower immune response abilities (island syndrome) (Lee and Klasing 2004). However, studies that have tested this hypothesis, both using experimental and theoretical approaches, have had contrasting results (Beadell et al. 2007; Hochberg and Møller 2001; Matson 2006).
Infections by different species of pathogens or by different strains of the same species within the same individual host or vector are common (Abbot et al. 2007; Cox 2001). In fact, parasite diversity in hosts seems to be a major evolutionary and ecological force for hosts (Bordes and Morand 2009). These concomitant infections can trigger cross-effective immune responses between pathogens that are antigenically similar, having thus an impact on the issue of the infection (Lee et al. 2010). An infection also can enhance susceptibility to subsequent infection (Cattadori et al. 2007). In particular, individuals with already are in poor physical condition may be more susceptible to multiple infections (Beldomenico and Begon 2009; Telfer et al. 2008). Furthermore, concomitant infection may allow the exchange of genetic material between strains of a given pathogen species or even between species through horizontal gene transfer (see Sect. 13.2.2 above), allowing the emergence of new virulent strains. An extreme case is one in which a pathogen drives the extinction of a population or species. Such scenarios are rare but do occur, generally due to a conjunction of pathogens and other causes. For instance, the decline of amphibian populations around the world is thought to be linked to a fungal pathogen Batrachochytrium dendrobatidis causing Chytridiomycosis (Crawford et al. 2010). Amphibians could have an increased susceptibility to the fungus due to changes in temperature variability (Rohr and Raffel 2010). Another example is the dramatic decline of the native red squirrel in the UK that has been attributed to a combination of direct competition with the grey squirrel and disease-mediated competition because the grey squirrel is a reservoir host of the squirrelpox virus that causes disease in the red squirrel (Tompkins et al. 2002). The local extinction of a host also may have tremendous consequences on an entire ecosystem (see for example the case of the wildebeest /rinderpest interactions in the Serengeti, Holdo et al. 2009).
3 How Does the Diversity of Hosts Influence Infectious Disease Ecology?
3.1 Change in Host Species Richness Modifies Infectious Disease Risk
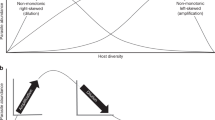
A change in species richness or abundance can modify the dynamics of local infectious diseases by either reducing or increasing the risk of transmission to the targeted species. The first outcome has been named, the “dilution effect”, the second, the “amplification effect”. The term “dilution effect” has conveyed different meanings since its first use in disease ecology literature (see Box 2 in the paper Keesing et al. 2006). The broad definition of the dilution effect refers to “the phenomenon – the net effect – when increased species diversity reduces disease risk” that is produced by a variety of mechanisms (“amplification effect” refers to the opposite phenomenon) (Keesing et al. 2006). This applies to vector-borne and directly transmitted diseases, although the concept of dilution has been developed most with regards to the tick-borne Lyme disease (Allan et al. 2003; LoGiudice et al. 2003, 2008).
The hypothesis underlying the amplification and dilution effect is that for many diseases, the competence of reservoirs, i.e. the ability to become infected and retransmit the pathogen, varies according to the host species (Haydon et al. 2002). The composition of the host community thus can influence the transmission dynamic of the disease. Similarly, since vectors have different competence to transmit pathogens, the composition of the vector community likely influences transmission dynamics. Different mechanisms are thought to be involved, but they are difficult to differentiate (Begon 2008; Keesing et al. 2006). One is the modification of the encounter rate (when reduced, this corresponds to the “dilution effect” sensu stricto). In the presence of species that are poorly competent, the transmission event that should link an infectious individual to a susceptible individual instead links infectious individuals to non-competent individuals. For vector-borne diseases, the increased diversity of a poorly competent host species on which the vector feeds increases the proportion of vector bites that are wasted. For direct transmission, the addition of non competent hosts can decrease transmission if these hosts remove infectious particles (Begon 2008). A second mechanism at work is that a high diversity of host species regulates the abundance of the competent host population. This regulation can be mediated by interspecific competition for limiting resources or by predation upon competent hosts. This typically is the idea that underlies biological controls, where organisms prey upon reservoir hosts, vectors or intermediate hosts (Straub and Snyder 2006). A third mechanism is based on the link between species richness and host mortality. This is the case when predators modify the mortality rate of a host and lower pathogen transmission by feeding on a heavily diseased individual (Packer et al. 2003). Two other mechanisms are cited by Keesing et al (2006), but they are difficult to demonstrate: (i) the modification of recovery when species added to a community facilitate the recovery of infected individuals by, for instance, providing resources, and (ii) the modification of transmission once the contact has occurred, for instance, when adding a species modifies the pathogen load within the host.
The dilution effect has been demonstrated mainly through theoretical work; there have been few case studies. One of the main examples is Lyme disease in the USA that is caused by pathogenic bacteria transmitted by ticks. These ticks feed readily on many species of vertebrates and these species vary in their degrees of reservoir competence. The white-footed mouse (Peromyscus leucopus) is thought to be the most competent host and dominates in fragmented forests. In native forests, which harbour a higher diversity of species than fragmented forests, ticks have a higher probability to dilute their bite by feeding on a less competent host (Allan et al. 2003; LoGiudice et al. 2003, 2008). However, such a dilution effect has not been demonstrated in Europe, probably because of the complexity of the disease ecology which involves numerous reservoir host and bacteria species (Halos et al. 2010). Another example is the West Nile virus, where an increased diversity of non passerine birds, which are less competent reservoir hosts compared to passerines, was associated with decreased West Nile virus infection in mosquitoes and humans (Ezenwa et al. 2006; Swaddle and Calos 2008). To date, there have been few examples of directly transmitted diseases, but studies on hantaviruses have shown that higher diversity of small mammals appears to regulate reservoir host populations through competition or predation. High small-mammal diversity also might inhibit intraspecific aggressive encounters between reservoir hosts that result in hantavirus transmission (Suzán et al. 2009).
In plants, crop diversity can reduce the total burden of disease in agricultural systems. This results from the combined effects of (i) the limitation of pathogen dispersal thanks to the physical barriers provided by the presence of non-host plants (Burdon and Chilvers 1982), (ii) induced systemic resistance, and (iii) competition among pathogens. The efficiency of crop mixtures is linked to the size of the area on which this method is used: a high level of success has been observed in a field trial with susceptible and resistant varieties of rice conducted on a large scale (3,342 ha) in China (Zhu et al. 2000).
Illustrations of amplification effects are typically the consequences of species introduction that radically modifies encounter rates. The added species can introduce new pathogens that infect native hosts (spillover) (Bruemmer et al. 2010) or amplify the circulation of local pathogens (spillback) (Kelly et al. 2009). The introduction of additional species also can provide sources of vector meals and increase vector numbers or activity (Saul 2003). For instance, the introduction of Siberian chipmunks (Tamias sibiricus) in suburban forests could increase the risk of Lyme disease because this host seems to be more competent than native hosts (Vourc’h et al. 2007). The introduction of the mosquito Aedes albopictus in many parts of the world has facilitated the transmission of the chikungunya virus (Benedict et al. 2007; Charrel et al. 2007).
Theoretical works based on deterministic modelling have looked at the conditions in disease transmission dynamics that are needed for the amplification or the dilution effect to occur (Begon 2008; Dobson 2004). When there is a relationship between the risk of a disease, the abundance of the reservoir host, and the abundance of an additional host, the addition of a species does not necessarily decrease the risk. In the case of Lyme disease, for example, tick abundance mainly is determined by the abundance of deer, which are in fact a non competent reservoir. An increased abundance of deer may reduce infection prevalence when immature ticks are feeding on the deer. At the same time, however, the overall number of adult ticks increase proportionally with the number of deer (Begon 2008). Further research in this field are relying on the modelling of the global community competence of hosts and vectors (Roche 2008).
Scientists and societies are increasingly interested in the dilution effect (MEA 2005) due to the link between habitat disturbance, generalist host characteristics, and their competence in disease transmission. Disturbance seems to favour generalist hosts (hosts that use different types of habitats) (Devictor et al. 2008; Marvier et al. 2004), and these hosts often have a broad geographical distribution (McKinney and Lockwood 1999; Smart et al. 2006). Crucially, these species also seem to have a higher competence reservoir or vector reservoir than species that are not favoured by disturbance (Mills 2006; Molyneux et al. 2008; Vittor et al. 2006). For example, many murid rodents that are recognized hosts of hemorrhagic fever viruses are opportunistic species that seem to be favoured in disturbed environments. The question is whether there is a causal link between a species’ generalist and opportunist character and its disease competence. Why are murid species associated with hemorrhagic fever more generalist than those which are not? Could it be possible that specialist species also carry hemorrhagic fever viruses, only these viruses have not yet been identified? Or is there something intrinsic in opportunistic species that makes them more likely to evolve and maintain hemorrhagic fever viruses (Mills 2006)?
3.2 Does a Loss in a Host’s Genetic Diversity Weaken Its Ability to Respond to Infectious Diseases?
Only a very small subset of plant and animal species have been domesticated (Diamond 2002). Many species of that small subset, for example, cattle (in animals) and maize (in plants), have seen their genetic diversity considerably reduced for the purpose of intensive production (The Bovine Hapmap Consortium, Matsuoka et al. 2002). In the wild, small populations of endangered species often have a very reduced genetic diversity (Keller and Waller 2002). This then raises the following question: to what extent does a reduction of the genetic diversity in a host species impact the ability of the host population to response to infectious diseases (May 1995)?
Genetic loci associated with the major histocompatibility complex (MHC) plays a key role in the acquired immune response of vertebrates (Altizer et al. 2003). MHC genes code for molecules that recognize foreign peptides (antigens) and display them on the cell surface. When the MHC-protein is displayed, it can be presented to immune cells, such as T lymphocytes or Natural Killer cells, which subsequently can trigger an adaptive immune response. Because MHC genes are faced with an important diversity of antigens, they must themselves be diverse. The measure of the genetic diversity of MHC based on an analysis of polymorphism sequences of MHC among individuals in populations has been widely used in conservation biology as a proxy to estimate the fitness of populations confronted by pathogens (Alcaide et al. 2010; Bernatchez and Landry 2003; Sommer 2005). However, the level of genetic variation at MHC loci results from different evolutionary forces (selection, gene flow, mutation) varying both in space and time in co-evolutionary systems involving both hosts and pathogens, making conservation genetics of non-model organism a challenging task (Stockwell et al. 2003). We already have many examples where low genetic diversity of species has favoured the diffusion of, and/or susceptibility to, pathogens. For example, the low genetic diversity of the Tasmanian devil could be involved in its susceptibility to facial tumor disease (McCallum 2008). The low genetic diversity found in commercially traded bee queens has been hypothesised as being one of the factors explaining colony collapse disorder (Le Conte and Navajas 2008). The problem is even more critical in intensive crops in which disease resistance has relied on the use of a very small number of genes. This selection strategy has proven to be ineffective as pathogens manage to overcome the resistance. For example, the resistance of Brassica napus (canola, oilseed rape and colza) to Leptosphaeria maculans (causing the blackleg disease) due to a major resistance gene was overcome in an area covering approximately 50,000 ha in South Australia in a period of 3 years (Sprague et al. 2006).
4 The Importance of Biodiversity as a Source of Treatment for Infectious Diseases
Even with advances in synthetic chemistry, which provides many biologically active molecules, pharmaceuticals derived from nature remain an important part of pharmaceutical practice today (Newman et al. 2008). All organisms have developed compounds to protect themselves against infectious diseases and to interact with individuals of their own species or other species (e.g. Rogerio et al. 2010). These molecules, coming from all organisms (bacteria, fungus, animals, plants) in terrestrial, marine and extreme ecosystems, represent an amazing diversity that has been tested in the field for millions of years by involving millions of individuals. However, only a very small subset of plants and marine organisms has been investigated for novel bioactive compounds. Furthermore, it is estimated that less that 1% of bacterial species and only 5% of fungal species are known. Those which have not yet been identified could be sources of novel molecules (Cragg and Newman 2005).
Observations of natural medicine practices used by indigenous people have led to the discovery of many drugs. The most well known and widely used pharmaceuticals are quinine, used as a model to synthesize anti-malarial drugs (chloroquine and mefloquine), and artemisinin, indentified as a potent anti-malarial drug by Chinese scientists (Newman et al. 2008). Animals also are a source of inspiration for drugs against infectious diseases. For instance, compounds of the sponge Cryptotethya crypta inspired the elaboration of antiviral medication such as AZT used in the treatment of HIV/AIDS (Cragg and Newman 2005). Observing great apes medicate themselves through the plants they eat also could help to reveal new active compounds (Krief et al. 2004).
5 Conclusion
Pathogens constitute a large part of biodiversity on earth and are present in all ecosystems and at all trophic levels, where they have a large impact on ecosystem functioning and on the population dynamics and evolution of their hosts. The recent acceleration of biodiversity loss due to human activities deeply impacts host-pathogen dynamics. Pathogens and hosts form co-evolving systems exercising major selective pressures on each other. Furthermore, the virulence or pathogenicity of a given species depends on its environment – which includes the hosts – that is highly variable in space and time. In such a context, human beings will never be able to completely control or eradicate every pathogen species; rather, we should accept that we must coexist with pathogens. To better understand and predict the evolution of pathogens and the impact of human activities on them, more in-depth studies are needed on how pathogens interact with host communities within different ecosystems.
To understand human, animal, and plant epidemics, two-species systems involving only a single host and a single pathogen species are no longer appropriate. The approach considering multi-host/multi-pathogen systems in their environment is the framework that now needs to be used to deepen our understanding of disease dynamics (Holt et al. 2003; Woolhouse et al. 2001) (Fig. 13.1). However, these dynamics are very complex and difficult to study because precise knowledge regarding the diversity of pathogens and of interactions taking place on several scales is lacking (Lloyd-Smith et al. 2009). In addition to intensive fieldwork to collect adequate data and modeling to understand the main processes, the use of molecular tools in a multi-host/multi-pathogen framework will facilitate investigations into pathogen-host community interactions. In particular, new generation sequencing techniques render it easier to characterize the genetic diversity of pathogens and hosts. For instance, metagenomic investigations allow an approach that does not require a priori assumptions that is useful to describe a community of pathogens and their interactions. Molecular techniques also may be used to clarify the taxonomic status of pathogens, revealing cryptic species or host races. With suitable molecular markers (producing a high level of polymorphism), the analysis of genetic variability within a spatially explicit framework renders it possible to identify the routes followed by a given pathogen. Moreover, molecular techniques can be used to identify genes involved in important life history traits of a pathogen such as virulence and transmission. Better knowledge of the mechanisms involved in host-pathogen interactions, and the extent of their variability, will significantly advance our understanding of outbreaks.

Schematic representation of the link between biodiversity and the ecology of infectious diseases. Diseases results from the complex interactions between the three compartments corresponding to pathogens, hosts and vectors (in the case of vector-borne diseases). The biodiversity of these three compartments can be considered at the community level (each circle corresponding to a species) or at the intraspecific level (each circle corresponding to a population or an individual within a population). The gray shading off of each unit considered (species, population or individual) illustrates its genetic or phenotypic variability in space and time, while variations in size illustrate frequency or density differences within the ecosystem. Interactions within each compartment can be direct (competition) or indirect (apparent competition…), synergic or antagonistic as illustrated by the different arrows
Although our knowledge of the number and variety of pathogens is not complete, it appears that their diversity, like that of their hosts and vectors, is greater in tropical areas than in temperate ones, and in undisturbed habitats than disturbed ones (Chaisiri et al. 2010; Friggens and Beier 2010). The reason we are so concerned by the loss of species biodiversity is because a reduction of biodiversity seems to favour opportunistic species that are highly competent as pathogen reservoirs and vectors. However, this observation was derived from only a few studies and theoretical works, mainly undertaken in the temperate zones. Further investigations should be launched to verify the link between, and understand the process involved in, biodiversity loss and disease dynamics. This especially should be done in the tropics to understand whether high levels of biodiversity create a buffer reducing the risk of disease transmission, and to understand the consequences of biodiversity loss in high biodiversity regions. With global changes, there is a high risk that diseases currently circulating in the tropics will reach temperate zones where species diversity is reduced and the availability of alternate hosts is limited. What could be the consequences of such a shift (Dobson et al. 2006)? What may happen in a world where increased movements of hosts and pathogens, high population densities, and rapidly changing environments increase contact rates, spread, and selective pressures on pathogens and hosts while at the same time a combination of low exposure to pathogen biodiversity and decreased genetic variability in some animals increases susceptibility to new diseases? The investigation of such questions requires collaboration across disciplines and between countries.
References
Abbot P, Aviles AE, Eller L et al (2007) Mixed infections, cryptic diversity, and vector-borne pathogens: evidence from Polygenis fleas and Bartonella species. Appl Environ Microbiol 73:6045–6052
Alcaide M, Lemus JA, Blanco G et al (2010) MHC diversity and differential exposure to pathogens in kestrels (Aves: Falconidae). Mol Ecol 19:691–705
Allan BF, Keesing F, Ostfeld RS (2003) Effect of forest fragmentation on Lyme disease risk. Conserv Biol 17:267–272
Altizer S, Harvell D, Friedle E (2003) Rapid evolutionary dynamics and disease threats to biodiversity. Trends Ecol Evol 18:589–596
Aminov RI, Mackie RI (2007) Evolution and ecology of antibiotic resistance genes. FEMS Microbiol Lett 271:147–161
Arias-González JE, Morand S (2007) Trophic functioning with parasites: a new insight for ecosystem analysis. Mar Ecol Prog Ser 320:43–53
Ashford RW, Crewe W (1998) The parasites of Homo sapiens: an annotated checklist of the protozoa, helminths and arthropods for which we are home. Liverpool School of Tropical Medicine, Liverpool, 728 p
Beadell JS, Atkins C, Cashion E et al (2007) Immunological change in a parasite-impoverished environment: divergent signals from four island taxa. PLoS One 2(9):e896
Begon M (2008) Effect of host diversity on disease dynamics. In: Ostfeld RS, Keesing F, Eviner VT (eds) Effects of ecosystems on disease and of disease on ecosystems. Princeton University Press, Princeton, pp 12–29
Begon M, Townsend CR, Harper JL (2006) Ecology: individuals, populations and communities. Blackwell, Oxford
Beldomenico PM, Begon M (2009) Disease spread, susceptibility and infection intensity: vicious circles? Trends Ecol Evol 25:21–27
Benedict MQ, Levine RS, Hawley WA et al (2007) Spread of the tiger: global risk of invasion by the mosquito Aedes albopictus. Vector Borne Zoonotic Dis 7:76–85
Bernatchez L, Landry C (2003) MHC studies in nonmodel vertebrates: what have we learned about natural selection in 15 years? J Evol Biol 16:363–377
Bordes F, Morand S (2009) Parasite diversity: an overlooked metric of parasite pressures? Oikos 118:801–806
Bordes F, Morand S, Krasnov BR et al (2010) Parasite diversity and latitudinal gradients in terrestrial mammals. In: Morand S, Krasnov BR (eds) The biogeography of host-parasite interactions. Oxford University Press, Oxford, pp 89–98
Breitbart M, Felts B, Kelley S et al (2004) Diversity and population structure of a near-shore marine-sediment viral community. Proc Biol Sci 271:565–574
Brooks DR, Hoberg EP (2001) Parasite systematics in the 21st century: opportunities 303 and obstacles. Trends Parasitol 17:273–275
Bruemmer CM, Rushton SP, Gurnell J et al (2010) Epidemiology of squirrelpox virus in grey squirrels in the UK. Epidemiol Infect 138:941–950
Burdon JJ, Chilvers GA (1982) Host density as a factor in plant-disease ecology. Annu Rev Phytopathol 20:143–166
Cattadori IM, Albert R, Boag B (2007) Variation in host susceptibility and infectiousness generated by co-infection: the myxoma-Trichostrongylus retortaeformis case in wild rabbits. J R Soc Interface 4:831–840
Chaisiri K, Chaeychomsri W, Siruntawineti J et al (2010) Human-dominated habitats and helminth parasitism in Southeast Asian murids. Parasitol Res 107:931–937
Charrel RN, de Lamballerie X, Raoult D (2007) Chikungunya outbreaks – the globalization of vectorborne diseases. New Engl J Med 356:769–771
Cleaveland S, Laurenson MK, Taylor LH (2001) Diseases of humans and their domestic mammals: pathogen characteristics, host range and the risk of emergence. Phil Trans R Soc Lond B 356:991–999
Cox FEG (2001) Concomitant infections, parasites and immune responses. Parasitology 122:S23–S38
Cragg GM, Newman DJ (2005) Biodiversity: a continuing source of novel drug leads. Pure Appl Chem 77:7–24
Crawford AJ, Lips KR, Bermingham E (2010) Epidemic disease decimates amphibian abundance, species diversity, and evolutionary history in the highlands of central Panama. Proc Natl Acad Sci USA 107:13777–13782
Cumming GS (2000) Using habitat models to map diversity: pan-African species richness of ticks (Acari: Ixodida). J Biogeogr 27:425–440
de Meeûs T, Michalakis Y, Renaud F (1998) Santa Rosalia revisited: or why are there so many kinds of parasites in ‘the garden of earthly delights’? Parasitol Today 14:10–13
Devictor V, Julliard R, Jiguet F (2008) Distribution of specialist and generalist species along spatial gradients of habitat disturbance and fragmentation. Oikos 117:507–514
Diamond J (2002) Evolution, consequences and future of plant and animal domestication. Nature 418:700–707
Dobson A (2004) Population dynamics of pathogens with multiple host species. Am Nat 164:S64–S78
Dobson A, Cattadori I, Holt RD et al (2006) Sacred cows and sympathetic squirrels: the importance of biological diversity to human health. PLoS Med 3:714–718
Dobson A, Lafferty KD, Kuris AM et al (2008) Homage to Linnaeus: how many parasites? How many hosts? Proc Natl Acad Sci USA 105:11482–11489
Drosopoulos WC, Rezende LF, Wainberg MA et al (1998) Virtues of being faithful: can we limit the genetic variation in human immunodeficiency virus? J Mol Med 76:604–612
Dunn RR, Davies TJ, Harris NC et al (2010) Global drivers of human pathogen richness and prevalence. Proc R Soc Lond B Biol Sci 277:2587–2595
Durrant WE, Dong X (2004) Systemic acquired resistance. Annu Rev Phytopathol 42:185–209
Ezenwa VO, Godsey MS, King RJ et al (2006) Avian diversity and West Nile virus: testing associations between biodiversity and infectious disease risk. Proc R Soc Lond B Biol Sci 273:109–117
Freitas-Junior LH, Bottius E, Pirrit LA et al (2000) Frequent ectopic recombination of virulence factor genes in telomeric chromosome clusters of P. falciparum. Nature 407:1018–1022
Friggens MM, Beier P (2010) Anthropogenic disturbance and the risk of flea-borne disease transmission. Oecologia 164:809–820
Gandon S, Day T (2007) The evolutionary epidemiology of vaccination. J R Soc Interface 4:803–817
Gandon S, Mackinnon MJ, Nee S et al (2001) Imperfect vaccines and the evolution of pathogen virulence. Nature 414:751–756
Gaston KJ, Blackburn TM (2000) Pattern and process in macroecology. Blackwell Science Publications, Oxford, 377 p
Gauld ID, Gaston KJ, Janzen DH (1992) Plant allelochemicals, tritrophic interactions and the anomalous diversity of tropical parasitoids: the ‘nasty’ host hypothesis. Oikos 65:352–357
Graham AL, Hayward AD, Watt KA et al (2010) Fitness correlates of heritable variation in antibody responsiveness in a wild mammal. Science 330:662–665
Guernier V, Hochberg ME, Guégan J-F (2004) Ecology drives the worldwide distribution of human diseases. PLoS Biol 2:e141
Halos L, Bord S, Cotté V et al (2010) Ecological factors characterizing the prevalence of bacterial tick-borne pathogens in Ixodes ricinus ticks in pastures and woodlands. Appl Environ Microbiol 76:4413–4420
Hamilton WD, Axelrod R, Tanese R (1990) Sexual reproduction as an adaptation to resist to parasites (a review). Proc Natl Acad Sci USA 87:3566–3573
Haydon DT, Cleaveland S, Taylor LH et al (2002) Identifying reservoirs of infection: a conceptual and practical challenge. Emerg Infect Dis 8:1468–1473
Hochberg ME, Møller AP (2001) Insularity and adaptation in coupled victim-enemy associations. J Evol Biol 14:539–551
Holdo RM, Sinclair ARE, Dobson AP et al (2009) A disease-mediated trophic cascade in the Serengeti and its implications for ecosystem C. PLoS Biol 7:e1000210
Holt RD, Dobson AP, Begon M et al (2003) Parasite establishment in host communities. Ecol Lett 6:837–842
Huang WJ, Zhang HY, Harrison TJ et al (2008) Cross-protection of hepatitis E virus genotypes 1 and 4 in rhesus macaques. J Med Virol 80:824–832
Hudson PJ, Dobson AP, Lafferty KD (2006) Is a healthy ecosystem one that is rich in parasites? Trends Ecol Evol 21:381–385
Huyse T, Poulin R, Theron A (2005) Speciation in parasites: a population genetic approach. Trends Parasitol 21:469–475
Ingram DS (1999) Biodiversity, plant pathogens and conservation. Plant Pathology 48:433–442
Jones JD, Dangl JL (2006) The plant immune system. Nature 444:323–329
Jones KE, Patel NG, Levy MA et al (2008) Global trends in emerging infectious diseases. Nature 451:990–993
Keesing F, Holt RD, Ostfeld RS (2006) Effects of species diversity on disease risk. Ecol Lett 9:485–498
Keller LF, Waller DM (2002) Inbreeding effects in wild populations. Trends Ecol Evol 17:230–241
Kelly DW, Paterson RA, Townsend CR et al (2009) Parasite spillback: a neglected concept in invasion ecology? Ecology 90:2047–2056
Krasnov BR, Shenbrot GI, Khokhlova IS et al (2004) Flea species richness and parameters of host body, host geography and host ‘milieu’. J Anim Ecol 73:1121–1128
Krief S, Martin MT, Grellier P et al (2004) Novel antimalarial compounds isolated in a survey of self-medicative behavior of wild chimpanzees in Uganda. Antimicrob Agents Chemother 48:3196–3199
Kuris AM, Hechinger RF, Shaw JC et al (2008) Ecosystem energetic implications of parasite and free-living biomass in three estuaries. Nature 454:515–518
Lafferty KD, Dobson AP, Kuris AM (2006) Parasites dominate food web links. Proc Natl Acad Sci USA 103:11211–11216
Le Conte Y, Navajas M (2008) Climate change: impact on honey bee populations and diseases. Revue Scientifique et Technique-Office International des Epizooties 27:499–510
Lee KA, Klasing KC (2004) A role for immunology in invasion biology. Trends Ecol Evol 19:523–529
Lee BH, Kim WH, Jeong J et al (2010) Prevalence and cross-immunity of Eimeria species on Korean chicken farms. J Vet Med Sci 72:985–989
Levy SB, Marshall B (2004) Antibacterial resistance worldwide: causes, challenges and responses. Nat Med 10:S122–S129
Lindenfors P, Nunn CL, Jones KE et al (2007) Parasite species richness in carnivores: effects of host body mass, latitude, geographical range and population density. Glob Ecol Biogeogr 16:496–509
Liu W, Li Y, Learn GH et al (2010) Origin of the human malaria parasite Plasmodium falciparum in gorillas. Nature 467:420–425
Lloyd-Smith JO, George D, Pepin KM et al (2009) Epidemic dynamics at the human-animal interface. Science 326:1362–1367
LoGiudice K, Ostfeld RS, Schmidt KA et al (2003) The ecology of infectious disease: effects of host diversity and community composition on Lyme disease risk. Proc Natl Acad Sci USA 100:567–571
LoGiudice K, Duerr STK, Newhouse MJ et al (2008) Impact of host community composition on Lyme disease risk. Ecology 89:2841–2849
Lyles AM, Dobson AP (1993) Infectious disease and intensive management: population dynamics, threatened hosts, and their parasites. J Zoo Wildlife Med 24:315–326
Marvier M, Kareiva P, Neubert MG (2004) Habitat destruction, fragmentation, and disturbance promote invasion by habitat generalists in a multispecies metapopulation. Risk Anal 24:869–878
Matson KD (2006) Are there differences in immune function between continental and insular birds? Proc R Soc Lond B Biol Sci 273:2267–2274
Matsuoka Y, Vigouroux Y, Goodman MM et al (2002) A single domestication for maize shown by multilocus microsatellite genotyping. Proc Natl Acad Sci USA 99:6080–6084
May RM (1995) Population genetics – the cheetah controversy. Nature 374:309–310
McCallum H (2008) Tasmanian devil facial tumour disease: lessons for conservation biology. Trends Ecol Evol 23:631–637
McKinney ML, Lockwood JL (1999) Biotic homogenization: a few winners replacing many losers in the next mass extinction. Trends Ecol Evol 14:450–453
MEA (2005) Millenium ecosystem assessment. In: Ecosystems and human well–being: biodiversity synthesis. Island Press, Washington, DC
Mills JN (2006) Biodiversity loss and emerging infectious disease: an example from the rodent-borne hemorrhagic fevers. Biodiversity 7:9–17
Molyneux DH, Ostfeld RS, Bernstein A et al (2008) Ecosystem disturbance, biodiversity loss, and human infectious disease. In: Chivian E, Bernstein A (eds) Sustaining life: how human health depends on biodiversity. Oxford University Press, Oxford, pp 287–323
Morris CE, Bardin M, Kinkel LL et al (2009) Expanding the paradigms of plant pathogen life history and evolution of parasitic fitness beyond agricultural boundaries. PLoS Pathog 5:e1000693
Newman DJ, Kilama J, Bernstein A et al (2008) Medicines from nature. In: Chivian E, Bernstein A (eds) Sustaining life: how human health depends on biodiversity. Oxford University Press, New York, pp 117–161
Nikolich MP, Hong G, Shoemaker NB et al (1994) Evidence for natural horizontal transfer of tetQ between bacteria that normally colonize humans and bacteria that normally colonize livestock. Appl Environ Microbiol 60:3255–3260
Normile D (2008) Driven to extinction. Science 319:1606–1609
Nunn CL, Altizer SM, Sechrest W et al (2005) Latitudinal gradients of parasite species richness in primates. Divers Distrib 11:249–256
Packer C, Holt RD, Hudson PJ et al (2003) Keeping the herds healthy and alert: implications of predator control for infectious disease. Ecol Lett 6:797–802
Page RDM (2003) Tangled trees: phylogeny, cospeciation, and coevolution. The University of Chicago Press, Chicago
Palumbi SR (2001) Evolution – humans as the world’s greatest evolutionary force. Science 293:1786–1790
Pedersen AB, Altizer S, Poss M et al (2005) Patterns of host specificity and transmission among parasites of wild primates. Int J Parasitol 35:647–657
Perlman SJ, Hunter MS, Zchori-Fein E (2006) The emerging diversity of Rickettsia. Proc R Soc Lond B Biol Sci 273:2097–2106
Poulin R (1995) Phylogeny, ecology and the richness of parasite communities in vertebrates. Ecol Monogr 65:283–302
Poulin R, Morand S (2004) Parasite biodiversity. Smithsonian Institution Press, Washington, DC
Prugnolle F, Durand P, Neel C et al (2010) African great apes are natural hosts of multiple related malaria species, including Plasmodium falciparum. Proc Natl Acad Sci USA 107:1458–1463
Riley MA, Lizotte-Waniewski M (2009) Population genomics and the bacterial species concept. Methods Mol Biol 532:367–377
Robar N, Burness G, Murray DL (2010) Tropics, trophics and taxonomy: the determinants of parasite-associated host mortality. Oikos 119:1273–1280
Roche B (2008) Complexité des écosystèmes, dynamique de la diversité biologique et maladies infectieuses. Une introduction à l’épidémiologie des communautés. University Montpellier II
Rogerio AP, Sa-Nunes A, Faccioli LH (2010) The activity of medicinal plants and secondary metabolites on eosinophilic inflammation. Pharmacol Res 62:298–307
Rohr JR, Raffel TR (2010) Linking global climate and temperature variability to widespread amphibian declines putatively caused by disease. Proc Natl Acad Sci USA 107:8269–8274
Saluzzo J-F, Vidal P, Gonzalez J-P (2004) Les virus émergents. IRD Editions, Paris, 188 p
Saul A (2003) Zooprophylaxis or zoopotentiation: the outcome of introducing animals on vector transmission is highly dependent on the mosquito mortality while searching. Malar J 2:32–50
Smart SM, Thompson K, Marrs RH et al (2006) Biotic homogenization and changes in species diversity across human-modified ecosystems. Proc R Soc Lond B Biol Sci 273:2659–2665
Sommer S (2005) The importance of immune gene variability (MHC) in evolutionary ecology and conservation. Front Zool 2:1–18
Sprague SJ, Marcroft SJ, Hayden HL et al (2006) Major gene resistance to blackleg in Brassica napus overcome within three years of commercial production in southeastern Australia. Plant Dis 90:190–198
Steffan SA, Snyder WE (2010) Cascading diversity effects transmitted exclusively by behavioral interactions. Ecology 91:2242–2252
Stockwell CA, Hendry AP, Kinnison MT (2003) Contemporary evolution meets conservation biology. Trends Ecol Evol 18:94–101
Straub CA, Snyder WE (2006) Experimental approaches to understanding the relationship between predator biodiversity and biological control. In: Brodeur J, Boivin G (eds) Trophic and guild in biological interactions control. Springer, Dordrecht, pp 221–239
Suzán G, Marcé E, Giermakowski JT et al (2009) Experimental evidence for reduced rodent diversity causing increased Hantavirus prevalence. PLoS One 4(5):e5461
Swaddle JP, Calos SE (2008) Increased avian diversity is associated with lower incidence of human West Nile infection: observation of the dilution effect. PLoS One 3:e2488
Taylor LH, Latham SM, Woolhouse MEJ (2001) Risk factors for human disease emergence. Phil Trans R Soc Lond B 356:983–989
Telfer S, Birtles R, Bennett M et al (2008) Parasite interactions in natural populations: insights from longitudinal data. Parasitology 135:767–781
Tompkins DM, Sainsbury AW, Nettleton P et al (2002) Parapoxvirus causes a deleterious disease in red squirrels associated with UK population declines. Proc R Soc Lond B Biol Sci 269:529–533
Vittor AY, Gilman RH, Tielsch J et al (2006) The effect of deforestation on the human-biting rate of Anopheles darlingi, the primary vector of Falciparum malaria in the Peruvian Amazon. Am J Trop Med Hyg 74:3–11
Vourc’h G, Marmet J, Chassagne M et al (2007) Borrelia burgdorferi sensu lato in Siberian chipmunks (Tamias sibiricus) introduced in suburban forests in France. Vector Borne Zoonotic Dis 7:637–641
Walpole M, Almond REA, Besancon C et al (2009) Tracking progress toward the 2010 biodiversity target and beyond. Science 325:1503–1504
Wilcox BA, Gubler DJ (2005) Disease ecology and the global emergence of zoonotic pathogens. Environ Health Prev Med 10:263–272
Windsor DA (1998) Most of the species on Earth are parasites. Int J Parasitol 28:1939–1941
Witter RL (1997) Avian tumor viruses: persistent and evolving pathogens. Acta Vet Hung 45:251–266
Wolfe ND, Dunavan CP, Diamond J (2007) Origins of major human infectious diseases. Nature 447:279–283
Woolhouse M, Gaunt E (2007) Ecological origins of novel human pathogens. Crit Rev Microbiol 33:231–242
Woolhouse MEJ, Gowtage-Sequeria S (2005) Host range and emerging and reemerging pathogens. Emerg Infect Dis 11:1842–1847
Woolhouse MEJ, Taylor LH, Haydon DT (2001) Population biology of multihost pathogens. Science 292:1109–1112
Woolhouse MEJ, Howey R, Gaunt E et al (2008) Temporal trends in the discovery of human viruses. Proc R Soc Lond B Biol Sci 275:2111–2115
Zhang T, Breitbart M, Lee WH et al (2006) RNA viral community in human feces: prevalence of plant pathogenic viruses. PLoS Biol 4:108–118
Zhu YY, Chen HR, Fan JH et al (2000) Genetic diversity and disease control in rice. Nature 406:718–722
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2012 Springer Science+Business Media B.V.
About this chapter
Cite this chapter
Vourc’h, G., Plantard, O., Morand, S. (2012). How Does Biodiversity Influence the Ecology of Infectious Disease?. In: Morand, S., Beaudeau, F., Cabaret, J. (eds) New Frontiers of Molecular Epidemiology of Infectious Diseases. Springer, Dordrecht. https://doi.org/10.1007/978-94-007-2114-2_13
Download citation
DOI: https://doi.org/10.1007/978-94-007-2114-2_13
Published:
Publisher Name: Springer, Dordrecht
Print ISBN: 978-94-007-2113-5
Online ISBN: 978-94-007-2114-2
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)