
Abstract
Background
Until now there has been no way of distinguishing between physiological and epileptic hippocampal ripples in intracranial recordings. In the present study we addressed this by investigating the effect of cognitive stimulation on interictal high frequency oscillations in the ripple range (80–250 Hz) within epileptic (EH) and non-epileptic hippocampus (NH).
Methods
We analyzed depth EEG recordings in 10 patients with intractable epilepsy, in whom hippocampal activity was recorded initially during quiet wakefulness and subsequently during a simple cognitive task. Using automated detection of ripples based on amplitude of the power envelope, we analyzed ripple rate (RR) in the cognitive and resting period, within EH and NH.
Results
Compared to quiet wakefulness we observed a significant reduction of RR during cognitive stimulation in EH, while it remained statistically marginal in NH. Further, we investigated the direct impact of cognitive stimuli on ripples (i.e. immediately post-stimulus), which showed a transient statistically significant suppression of ripples in the first second after stimuli onset in NH only.
Conclusion
Our results point to a differential reactivity of ripples within EH and NH to cognitive stimulation.
Similar content being viewed by others


Background
Transient high-frequency oscillations (HFOs) in the frequency range 80–250 Hz have been recorded repeatedly in animal and human hippocampi. These field potentials, referred to as “ripples”, were observed initially with microrecordings in rats and later in human hippocampi of epileptic patients [1–3]. Ripples are believed to reflect short-term synchronization of neuronal activity and appear to play important roles in both normal and pathological brain functions [4, 5]. More recently, strong evidence has been presented for a link between memory consolidation and hippocampal ripples in both micro- and macroelectrode recording studies [6, 7]; ripples related to the strengthening and reorganization of memory traces are observed during slow wave sleep and quiet wakefulness [7–10].
Importantly, ripples have also been observed in the epileptic dentate gyrus, wherein they are absent in healthy animals [4, 11–13]. It seems feasible, therefore, that a proportion of ripples recorded within the epileptic hippocampus are linked to underlying pathological processes. Indeed, clinical macroelectrode recordings within mesial temporal structures in epileptic patients inconsistently report an increase of ripples on the side of epileptogenic tissue. This contrasts with observations from previous microrecordings in animals [14]. This has led some to suggest that very large electrodes may filter out physiological HFOs, such that only pathological HFOs remain visible [15]. Even in macroelectrode studies, however, different electrode types might lead to differing results concerning the contribution of hippocampal ripples to the lateralization of an epileptogenic region. Furthermore, their clinical/diagnostic utility is compromised by the inability to distinguish normal from pathological ripples in invasive electroencephalographic (EEG) recordings. Achieving such a differentiation is important not only to understand the fundamental mechanisms behind normal cognitive functions, but also for the utilization of ripples as a potential clinical biomarker for the identification of an epileptogenic region.
The aim of the present study was to investigate a potential means with which to differentiate between physiological and epileptic ripples in intracranial recordings. Specifically, we tested our hypothesis that cognitive stimulation would have a differing effect on ripples within epileptic and non-epileptic hippocampi in human subjects. To do so, we analyzed hippocampal activity in ten epileptic patients during quiet wakefulness and during a cognitive task.
Methods
Subjects
Our sample comprised 10 patients (three males) ranging in age from 20 to 47 years (mean age: 31 ± 7.5 years), all with medically intractable focal epilepsies (Table 1). All subjects were on chronic anticonvulsant medication, reduced slightly in most cases for the purpose of video-EEG monitoring. All but one of the patients (no. 3) were right handed. Written informed consent was obtained from each subject prior to the investigation and the study received approval from the Ethics Committee of Masaryk University, Brno.
EEG recordings
Depth electrodes were implanted to localize seizure origin prior to surgical treatment. Each patient received 5–11 orthogonal platinum electrodes in the temporal and/or frontal, parietal, and occipital lobes using the stereotaxic coordinate system of Talairach [16]. Standard depth electrodes (ALCIS) were used with a diameter of 0.8 mm, a contact length of 2 mm, and an inter-contact distance of 1.5 mm. The exact positions of the electrode contacts in the brain were verified using postplacement MRI with electrodes in situ. The EEG signal was recorded with sampling rate of 1,024 Hz (TrueScan EEG system, Deymed Diagnostic). All recordings were referenced to a linked earlobe. All impedances were less than 5 kΩ. In this study, we investigated EEG data from 61 contacts positioned in epileptic (31) and non-epileptic (30) hippocampi (Table 1). This categorization was performed according to the results of standard visual analysis of ictal stereo EEG recordings; the seizure onset zone (SOZ) was defined as the contacts showing the first EEG ictal activity.
Behavioral tasks
During the initial rest period, subjects were asked to relax for 30 min while sitting comfortably in a reclining position with eyes closed. For the subsequent cognitive phase they performed a visual oddball task. Stimuli consisted of capital letters presented randomly in the center of a monitor, with target (“X”) and frequent (“O”) trials intermixed with other distractor letters at a ratio of 1:4.6:1 (target:frequent:distractor). There were 50 target stimuli. The stimuli were presented for 500 ms, and the inter-stimulus interval varied between 4 and 6 s. Each subject was instructed to respond to the target stimulus as quickly as possible by pressing a button with their dominant hand.
Data analysis and statistics
Using an automated detection of ripples based on the signal power envelope, we analyzed potential differences in ripple rate (RR) during the cognitive compared with the resting period, within epileptic (EH) and non-epileptic hippocampi (NH). Further, we compared the direct impact of cognitive processes on ripples (i.e. immediately post-stimulus) in EH and NH.
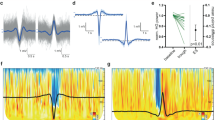
In the pre-detection stage the signal power envelope for 80 and 250 Hz band pass was calculated using the Hilbert transform. The HFOs were detected using normalized power envelope amplitude and duration thresholds (Figure 1). In order to stress the high power events and suppress the surrounding signal, the normalization of signal power envelopes (normPE) was performed by subtracting 2/3 percentile (p66) of the signal power envelope PE(f) and subsequently dividing it by half the value of the 2/3 and 1/3 percentile (p33) difference:

A demonstration of ripple detection. Top to bottom raw data from a single subject and contact; ripples within the signal filtered at 80–250 Hz; automated detection using amplitude of power envelope.
The amplitude threshold values of normPE were based on normalized characteristics of HFOs that were scored previously by expert reviewers in various empirical data sets (Figures 1, 2). The duration threshold was set to a minimum duration of ~30 ms which was ~5 oscillations at 150 Hz.

Block diagram of computational method for ripple detection.
To assess the effect of the cognitive stimuli, we examined the occurrence of ripples/RR for each contact in a moving window with the length of 0.5 s shifted in 0.05 s increments. The statistical significance of differences relative to baseline (−0.6 to −0.1 s pre-stimulus) was analyzed by Wilcoxon signed rank test.
We then investigated whether any differences existed in the duration of HFOs between EH and NH, during both resting-state and cognitive stimulation. As the durations of HFOs varied according to a normal distribution, we used unpaired t test to identify any significant differences.
To distinguish between ripples and high-frequency activities (HFAs), the latter of which appear to be related to the multiunit firing rate (e.g. task-induced gamma), we further generated time frequency maps (TFM) and power envelopes averaged to trigger stimuli in the frequency range 80–250 Hz. TFM provides an overview of the time–frequency increase in gamma activity associated with stimuli, and power envelopes enable comparisons between induced gamma power and power of ripples. These computations were performed on the signals from all hippocampal contacts and montages.
Results
Ripples were detected within hippocampal recordings from all subjects. Mean RR (across all EH or NH contacts) in resting-state periods was 16.4/min [SD = 12.2; range 1–58] within EH, and 19.4/min [SD = 17.4; range 0–73] within NH. Over cognitive-task periods, mean RR within EH and NH decreased to 11.2/min [SD = 7.7; range 1–35] and 17.2 [SD = 15.0; range 5–55], respectively (Figure 3). Median and quantile [0.1/0.9] values were as follows: resting-state EH 11.9 [2.7/32.6]; resting-state NH 12.8 [5.4/41.5]; cognitive task EH 8.4 [3.1/22.6]; cognitive task NH 10.9 [5.6/47.7]. The reduction of mean RR during the execution of cognitive task was significant in EH (p < 0.001) but only marginal in NH (p = 0.06; indicating a trend). There was no significant difference in RR within EH versus NH during the resting state, but a significant difference was revealed by statistical analysis of RR between EH and NH during cognitive task performance (p < 0.05).

Ripple rates during resting-state and cognitive-task periods within epileptic and non-epileptic hippocampi across all investigated subjects. Black asterisk means significant difference in epileptic hippocampus (p < 0.05).
Investigating the immediate impact of cognitive stimuli on ripple occurrence, we observed a statistically significant short-lasting suppression of ripples in NH within the first second after stimuli onset. This effect was virtually missing in EH (Table 2; Figures 4, 5). Interestingly, the short-lasting suppression in NH was followed by a robust and significant transient increase in RR approximately one second after the stimuli. A similar but somewhat later significant RR increase was also observed in EH (Figure 4a).

Immediate short-lasting impact of cognitive stimuli on ripple rate across subjects. a Transient suppression of relative ripple rate within epileptic (upper a) and non-epileptic (bottom a) hippocampus. Red vertical line defines visual stimulation onset (trigger). Full lines represent median, dotted lines 25 and 75 percentile across all subjects and all recording contacts. The figure clearly demonstrates task-induced HFOs reduction in non-epileptic hippocampus in time period approximately 0.3–1 s after the stimulation (arrow). White and gray horizontal bars indicate an area that corresponds to the box plots in the right b. b Box plots computed in baseline period before stimuli (−0.6 to −0.1 s) and after cognitive stimulation (0.4–0.9 s). Black asterisk means significant difference in non-epileptic hippocampus (p < 0.02).

Observations from a single contact within the non-epileptic hippocampus of subject (patient no. 6). Bottom to top the transient post-stimulus decrease in ripple occurrence within non-epileptic hippocampus coincides with event related 80–250 Hz power envelope reduction, the genesis of local-field cognitive potential (P3). S stimulus.
The mean duration of ripples in resting-state periods was 88.1 ms [SD = 7.4 ms; range 73.8–150.6 ms; median = 87.9 ms; 10% quantile = 77.8 ms] within EH, and 82.8 ms [SD = 9.8 ms; 61.0–109.2 ms; median = 81.2 ms, 10% quantile = 71.8 ms] within NH. In cognitive-task periods, mean ripple duration (RD) within EH and NH were 85.5 (SD = 5.2 ms; range 71.9–98.8 ms; median = 85.9 ms; 10% quantile = 80.1 ms] and 83 (SD = 10.1 ms; range 63.0–100.6 ms; median 85.9 ms, 10% quantile = 70.0 ms], respectively. There was a significant difference between EH and NH ripple durations during quiet wakefulness (p < 0.001), but we did not observe any significant EH/NH difference for RD under the cognitive load (p = 0.136). The changes in ripple duration during cognitive stimulation did not reach statistical significance for either EH (p = 0.052) nor NH (p = 0.902), although a trend was observed in case of EH.
Individual TFM analyses of signals from all the investigated non-epileptic hippocampal sites revealed unequivocal task-induced gamma (HFA) in 6 of 30 channels (3 subjects in total), and only after targets. The frequency range of this gamma activity overlapped partially with ripple activity. However, averaged TFM and power envelopes reveal that the power of induced gamma on single trials was significantly lower than the threshold for ripple detection. This significant difference in power excludes the false detection of induced gamma activity as ripples.
Discussion
In the present study we have used simple cognitive task to investigate whether the effect of cognitive stimulation on hippocampal ripples can be used as a new approach for distinguishing presumably normal HFOs in NH from presumably pathological HFOs in EH. We observed significantly different, and in some aspects opposite, behavior of ripples within EH and NH. We must stress that we have not explored the nature of ripples in both NH or EH in the present experiment; our finding of different HFO content/dynamics in the response of the epileptic and non-epileptic hippocampus to cognitive stimulation offers little insight into the difference between physiological and pathological ripples. On the other hand there is a consensus to support that ripples are not only normal activity in hippocampus. Many reports from humans and non-primates showed an increase in ripple frequency HFO at, or prior to, the onset of seizures [17]. It seems unlikely that these “pathological” ripples reflect just an “exaggerated” version of physiological activity as suggested recently by [6]. Rather, while physiological hippocampal ripples (and underlying sharp wave-ripple/SPWR/complexes) appear to reflect summated synchronous inhibitory postsynaptic potentials generated by subsets of interneurons regulating the discharges of principal cells [1], epileptic HFOs represent field potentials of population spikes from clusters of abnormal synchronously bursting neurons [4, 18, 19]. Recent cellular evidence suggests that physiological ripple oscillations reflect phasic perisomatic inhibitory synaptic potentials in pyramidal cells, together with sparse phase-locked firing and rhythmic depolarizing potentials [20–22]. Inhibitory interneurons could then secure an orderly recruitment of pyramidal cells [23, 24]. Pathological ripple-like high-frequency oscillations might represent heterogeneous cellular and synaptic phenomena. The recent study of Alvarado-Rojas et al. revealed an involvement of distinct synaptic processes and different mechanisms of synchrony in the origin of ripple-like HFOs (150–250 Hz) during interictal (IID) and preictal epileptic discharges (PID). IID ripples were associated with rhythmic inhibitory postsynaptic potentials and weak phase-locked firing, whilst PID were associated with depolarizing potentials that usually triggered rhythmic burst firing [21]. Both types of pathological ripples must be distinguished from fast ripples in the 250–800 Hz range, which reflect population spikes of partially synchronous, massively bursting, uninhibited pyramidal cells [22]. Regardless, the present findings of different ripple dynamics to external stimulation seem to suggest diverse mechanisms behind their generation.
The question remains whether the mechanisms of generation, as well as the functional significance of waking- and sleep-related ripples (both normal and pathological), are similar or different. Unfortunately, sleep related HFOs have been studied much more extensively in the past, with less evidence coming from ripple studies performed during quiet wakefulness. A growing number of papers have, however, focused on awake SPWRs, suggesting their relationship to behavioral performance and complementary role in memory consolidation with sleep SPWRs [7, 9, 25–28]. It was shown that physiological as well as pathological hippocampal HFOs can be recorded reliably with standard macroelectrodes in awake periods, and it is interestingly that the occurrence of ripples changes as a function of the state of vigilance. However, the results of previous micro- and macroelectrode studies on memory are contradictory; using microelectrodes recordings in rodents, monkeys, and humans, ripples appear typically to be more present during immobility and slow-wave sleep [1, 29, 30]. On the other hand investigating HFOs from human non-epileptic hippocampus during a memory consolidation task and using macroelectrodes recordings, [7] report that the majority of all hippocampal ripples occurred during waking state, with only a minority occurring during stages of deep sleep. In contrast, epileptic ripples are significantly more frequent during non-REM sleep compared to epochs of wakefulness in both micro- and macrorecordings [31].
Evaluating ripple occurrence in resting and task periods, our study revealed a significant decrease of HFO rate in epileptic tissue during event discrimination. This finding might reflect an increased involvement of hippocampal neurons in physiological cognitive processing, and consequently decreased synchronization within the network driven by synchronously bursting epileptic neurons. This hypothesis seems to be congruent with the well-known prevalence of epileptic ripples during non-REM sleep (see above), which results very likely from the sleep-dependent enhancement of network synchronization within the mesial aspect of the temporal lobe [32–34]. Our observation that cognitive stimulation only marginally impact upon general ripple rate within the non-epileptic hippocampi can be explained by anticipated involvement of normal hippocampal neurons in both memory consolidation/awake neuronal replay (rest period) and complex event discrimination processing (task period). Finally, the oddball task used in our study is linked intimately with an information-processing cascade, during which attentional and memory mechanisms are engaged preferentially [35].
In contrast to the significant effect of event processing on long-standing RR within epileptic hippocampi and only marginal effect in presumably healthy structures, we observed an immediate short-lasting impact of cognitive stimuli on ripple occurrence in non-epileptic hippocampi only. Specifically, we reveal that single cognitive stimuli appear to suppress NH ripples briefly. Despite difference in experimental paradigms, this finding might be analogous to the selective suppression of SPWRs by timed electrical stimulation [36, 37]. Noteworthy, in a study published recently [38], no disruption of SPWRs was observed during the presentation of simple non-cognitive light stimuli in adult rabbits. The question thus remains whether or not simple sensory stimulation is sufficiently potent to disrupt physiological hippocampal ripples in human macrorecordings. In our study this immediate effect took approximately 500 ms and culminated in the peaking of an averaged event-related potential known as P3, which consists largely of contributions from theta and delta oscillations (Figure 5). We can therefore assume a compromising of the activity of normal hippocampal neurons between distinct physiological processes, resulting in either high frequency or slow oscillations. Finally, the subsequent transient increase in ripple occurrence approximately 1 s after stimulus onset might represent a simple rebound of HFOs or possibly true increase of ripples, that was recently observed in human hippocampi approximately 800 ms after the presentation of typical visual memory stimuli [41]. Theoretically it may also reflect some task-induced high-frequency neural activity (HFA), which seems to be related largely to multi-unit activity (MUA) and could be linked to memory consolidation and ongoing retrieval of stored memories [8, 10, 39, 40]. In our study, however, the methodology used for ripple identification prevented the false detection of task-induced gamma; averaged power envelopes revealed very different powers in ripples and gamma activities, the latter of which was significantly lower than the threshold level for ripple detection. Particularly noteworthy is our observation of task-induced gamma in only a minority of hippocampal contacts and subjects in this study. This might be somewhat surprising given that the hippocampus is likely to be engaged in some memory processes during the oddball task [35]. On the other hand, this limited occurrence of task-induced gamma in hippocampal regions after target stimuli might explain the small increase in the blood oxygen level-dependent (BOLD) signal (in which gamma is reflected) within these regions in functional MRI studies using the oddball task [42]. Finally, task-induced gamma occurs usually between 100 and 400 ms after stimuli presentation, and is therefore likely to precede ripple suppression [43]. Regardless, the more limited direct impact of external stimuli on ripple rate within epileptic hippocampi likely reveals the low reactivity of epileptic neurons to physiological events.
Distinguishing normal and pathological HFOs represents currently one of the most challenging tasks for basic and clinical neuroscience. Pathological HFOs may have important diagnostic and prognostic value, serving potentially as biomarkers for the epileptogenic zone that is a crucial target for highly effective epilepsy surgery. Research on normal HFOs can then give us a better understanding of memory encoding and consolidation, as well as better insight into the distortion of these processes in neurocognitive disorders. To the best of our knowledge this is the first study providing evidence of significantly different effects of external sensory stimulation on ripples within epileptic and non-epileptic hippocampi. Until now only one published paper has addressed specifically the differentiation of normal and pathological HFOs. Using visual or motor task in five epileptic patients, distinct parameters of two types HFOs have been suggested [43]. The identification of normal and pathological HFOs according to frequency content is currently a matter of intense experimental scrutiny, and certainly cannot be used within the ripple range [44]. Despite previous suggestions of the distinct filtering of normal HFOs by very large electrodes, this effect is highly equivocal [14]. Even in our present study we found slightly (but not significantly) higher RR in non-epileptic compared to epileptic hippocampi, a result that clearly contradicts the proposed “filtering” effect. No other approaches to reach this “holy grail” for hippocampal HFOs are available today.
An obvious limitation of our study is the analyses of data from chronic epileptic patients only. Even if we carefully differentiated epileptic and non-epileptic hippocampi with presumed epileptic and normal ripples, the impact of epileptic activity on normal neurons even within non-epileptic hippocampi cannot be excluded completely. This risk is higher when treating contralateral hippocampus in patients suffering from unilateral temporal lobe epilepsy. In our study, this risk was reduced because the majority of non-epileptic hippocampal recordings were taken from extratemporal epilepsy patients. Still, translating our “normal” results into normal hippocampus behavior must be done with great caution. On the other hand, intracerebral EEG recordings are possible only in patients suffering from major brain disorders. As such, this limitation must be kept in mind.
Conclusions
In conclusion, the results of our study point to a differential reactivity of ripples recorded from within EH and NH to cognitive stimulation. This discovery could present a possible means with which to identify hippocampal epileptic ripples in laboratories that record intracerebral EEG data with the standard sampling frequency of 1 kHz.
References
Buzsaki G, Horvath Z, Urioste R, Hetke J, Wise K (1992) High-frequency network oscillation in the hippocampus. Science (80) 256:1025–1027
Bragin A, Engel J, Wilson CL, Fried I, Buzsaki G (1999) High-frequency oscillations in human brain. Hippocampus 9:137–142
Staba RJ, Wilson CL, Bragin A, Fried I, Engel J (2002) Quantitative analysis of high-frequency oscillations (80–500 Hz) recorded in human epileptic hippocampus and entorhinal cortex. J Neurophysiol 88:1743–1752
Engel J, Bragin A, Staba R, Mody I (2009) High-frequency oscillations: what is normal and what is not? Epilepsia 50:598–604
Engel J, da Silva FL (2012) High-frequency oscillations—where we are and where we need to go. Prog Neurobiol 98:316–318
Buzsaki G, da Silva FL (2012) High frequency oscillations in the intact brain. Prog Neurobiol 98:241–249
Axmacher N, Elger CE, Fell J (2008) Ripples in the medial temporal lobe are relevant for human memory consolidation. Brain 131:1806–1817
Carr MF, Jadhav SP, Frank LM (2011) Hippocampal replay in the awake state: a potential substrate for memory consolidation and retrieval. Nat Neurosci 14:147–153
Girardeau G, Zugaro M (2011) Hippocampal ripples and memory consolidation. Curr Opin Neurobiol 21:452–459
Lachaux J-P, Axmacher N, Mormann F, Halgren E, Crone NE (2012) High-frequency neural activity and human cognition: {Past}, present and possible future of intracranial {EEG research}. Prog Neurobiol 98:279–301
Buzsaki G (1986) Hippocampal sharp waves—their origin and significance. Brain Res 398:242–252
Bragin A, Jando G, Nadasdy Z, Hetke J, Wise K, Buzsaki G (1995) Gamma (40–100-Hz) oscillation in the hippocampus of the behaving rat. J Neurosci 15:47–60
Bragin A, Wilson CL, Almajano J, Mody I, Engel T (2004) High-frequency oscillations after status epilepticus: epileptogenesis and seizure genesis. Epilepsia 45:1017–1023
Jacobs J, Staba R, Asano E, Otsubo H, Wu JY, Zijlmans M et al (2012) High-frequency oscillations (HFOs) in clinical epilepsy. Prog Neurobiol 98:302–315
Crepon B, Navarro V, Hasboun D, Clemenceau S, Martinerie J, Baulac M et al (2010) Mapping interictal oscillations greater than 200 Hz recorded with intracranial macroelectrodes in human epilepsy. Brain 133:33–45
Talairach J (1967) Atlas D’anatomie Stéréotaxique Du Télencéphale; Études Anatomo-Radiologiques, Par. J. Talairach [et Al.]. Masson, Paris
Worrell GA, Gardner AB, Stead SM, Hu S, Goerss S, Cascino GJ et al (2008) High-frequency oscillations in human temporal lobe: simultaneous microwire and clinical macroelectrode recordings. Brain 131:928–937
Ibarz JM, Foffani G, Cid E, Inostroza M, de la Prida L (2010) Emergent dynamics of fast ripples in the epileptic hippocampus. J Neurosci 30:16249–16261
Bragin A, Benassi SK, Kheiri F, Engel J (2011) Further evidence that pathologic high-frequency oscillations are bursts of population spikes derived from recordings of identified cells in dentate gyrus. Epilepsia 52:45–52
Maier N, Tejero-Cantero A, Dorrn AL, Winterer J, Beed PS, Morris G et al (2011) Coherent phasic excitation during hippocampal ripples. Neuron 72:137–152
Alvarado-Rojas C, Huberfeld G, Baulac M, Clemenceau S, Charpier S, Miles R et al (2015) Different mechanisms of ripple-like oscillations in the human epileptic subiculum. Ann Neurol 77:281–290
Gulyás AI, Freund TT (2015) Generation of physiological and pathological high frequency oscillations: the role of perisomatic inhibition in sharp-wave ripple and interictal spike generation. Curr Opin Neurobiol 31:26–32. [{SI}: {Brain} Rhythms and Dynamic Coordination]
Csicsvari J, Hirase H, Mamiya A, Buzsaki G (2000) Ensemble patterns of hippocampal CA3–CA1 neurons during sharp wave-associated population events. Neuron 28:585–594
Klausberger T, Somogyi P (2008) Neuronal diversity and temporal dynamics: the unity of hippocampal circuit operations. Science (80) 321:53–57
Kudrimoti HS, Barnes CA, McNaughton BL (1999) Reactivation of hippocampal cell assemblies: effects of behavioral state, experience, and EEG dynamics. J Neurosci 19:4090–4101
O’Neill J, Senior T, Csicsvari J (2006) Place-selective firing of CA1 pyramidal cells during sharp wave/ripple network patterns in exploratory behavior. Neuron 49:143–155
Davidson TJ, Kloosterman F, Wilson MA (2009) Hippocampal replay of extended experience. Neuron 63:497–507
Gupta AS, van der Meer MAA, Touretzky DS, Redish AD (2010) Hippocampal replay is not a simple function of experience. Neuron 65:695–705
Skaggs WE, McNaughton BL, Permenter M, Archibeque M, Vogt J, Amaral DG et al (2007) EEG sharp waves and sparse ensemble unit activity in the Macaque hippocampus. J Neurophysiol 98:898–910
Van Quyen M, Le Bragin A, Staba R, Crepon B, Wilson CL, Engel J (2008) Cell type-specific firing during ripple oscillations in the hippocampal formation of humans. J Neurosci 28:6104–6110
Bagshaw AP, Jacobs J, LeVan P, Dubeau F, Gotman J (2009) Effect of sleep stage on interictal high-frequency oscillations recorded from depth macroelectrodes in patients with focal epilepsy. Epilepsia 50:617–628
Steriade M, Contreras D, Amzica F (1994) Synchronized sleep oscillations and their paroxysmal developments. Trends Neurosci 17:199–208
Dinner DS (2002) Effect of sleep on epilepsy. J Clin Neurophysiol 19:504–513
De Guzman PH, Nazer F, Dickson CT (2010) Short-duration epileptic discharges show a distinct phase preference during ongoing hippocampal slow oscillations. J Neurophysiol 104:2194–2202
Polich J (2007) Updating p300: an integrative theory of P3a and P3b. Clin Neurophysiol 118:2128–2148
Girardeau G, Benchenane K, Wiener SI, Buzsaki G, Zugaro MB (2009) Selective suppression of hippocampal ripples impairs spatial memory. Nat Neurosci 12:1222–1223
Ego-Stengel V, Wilson MA (2010) Disruption of ripple-associated hippocampal activity during rest impairs spatial learning in the rat. Hippocampus 20:1–10
Nokia MS, Mikkonen JE, Penttonen M, Wikgren J (2012) Disrupting neural activity related to awake-state sharp wave-ripple complexes prevents hippocampal learning. Front Behav Neurosci 6:84
Miller KJ, Zanos S, Fetz EE, den Nijs M, Ojemann JG (2009) Decoupling the cortical power spectrum reveals real-time representation of individual finger movements in humans. J Neurosci 29:3132–3137
Manning JR, Jacobs J, Fried I, Kahana MJ (2009) Broadband shifts in local field potential power spectra are correlated with single-neuron spiking in humans. J Neurosci 29:13613–13620
Kucewicz MT, Cimbalnik J, Matsumoto JY, Brinkmann BH, Bower MR, Vasoli V et al (2014) High frequency oscillations are associated with cognitive processing in human recognition memory. Brain 137(Pt 8):2231–2244
Brazdil M, Dobsik M, Mikl M, Hlustik P, Daniel P, Pazourkova M et al (2005) Combined event-related fMRI and intracerebral ERP study of an auditory oddball task. Neuroimage 26:285–293
Matsumoto A, Brinkmann BH, Stead SM, Matsumoto J, Kucewicz MT, Marsh WR et al (2013) Pathological and physiological high-frequency oscillations in focal human epilepsy. J Neurophysiol 110:1958–1964
Jefferys JGR, de la Prida L, Wendling F, Bragin A, Avoli M, Timofeev I et al (2012) Mechanisms of physiological and epileptic HFO generation. Prog Neurobiol 98:250–264
Authors’ contributions
MB conceived of the study, designed the experiment, drafted the manuscript and gave final approval of the version to be published. JC participated in the design of the study and performed the data analysis. RR participated in the design of the study and carried out the experiment with the acquisition of data. PD participated in data acquisition and was involved in drafting the manuscript. PJ has been involved in data interpretation. JH participated in data analysis and interpretation, and revised the final manuscript version for important intellectual content. MMS developed analysis tool and participated in data interpretation. DJS participated in data interpretation and drafting the manuscript. All authors read and approved the final manuscript.
Acknowledgements
We thank the patients for participating in this study, Jan Chladek for his contribution in data analysis (TFM). The study was supported by the project “CEITEC-Central European Institute of Technology” (CZ.1.05/1.1.00/02.0068) from European Regional Development Fund and Grant GACR P103/11/0933. The technical part of the study was supported by the ‘Application laboratories of advanced microtechnologies and nanotechnologies’ project, CZ.1.05/2.1.00/01.0017, co-funded by the ‛Research and Development for Innovations’ Operational Programme, the European regional development fund, and the state budget.
Compliance with ethical guidelines
Competing interests The authors declare that they have no competing interests.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Brázdil, M., Cimbálník, J., Roman, R. et al. Impact of cognitive stimulation on ripples within human epileptic and non-epileptic hippocampus. BMC Neurosci 16, 47 (2015). https://doi.org/10.1186/s12868-015-0184-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12868-015-0184-0