

Abstract
Progressive phases of multiple sclerosis are associated with inhibited differentiation of the progenitor cell population that generates the mature oligodendrocytes required for remyelination and disease remission. To identify selective inducers of oligodendrocyte differentiation, we performed an image-based screen for myelin basic protein (MBP) expression using primary rat optic-nerve-derived progenitor cells. Here we show that among the most effective compounds identifed was benztropine, which significantly decreases clinical severity in the experimental autoimmune encephalomyelitis (EAE) model of relapsing-remitting multiple sclerosis when administered alone or in combination with approved immunosuppressive treatments for multiple sclerosis. Evidence from a cuprizone-induced model of demyelination, in vitro and in vivo T-cell assays and EAE adoptive transfer experiments indicated that the observed efficacy of this drug results directly from an enhancement of remyelination rather than immune suppression. Pharmacological studies indicate that benztropine functions by a mechanism that involves direct antagonism of M1 and/or M3 muscarinic receptors. These studies should facilitate the development of effective new therapies for the treatment of multiple sclerosis that complement established immunosuppressive approaches.




Similar content being viewed by others
References
Franklin, R. J. & Ffrench-Constant, C. Remyelination in the CNS: from biology to therapy. Nature Rev. Neurosci. 9, 839–855 (2008)
Franklin, R. J. Why does remyelination fail in multiple sclerosis? Nature Rev. Neurosci. 3, 705–714 (2002)
Nunes, M. C. et al. Identification and isolation of multipotential neural progenitor cells from the subcortical white matter of the adult human brain. Nature Med. 9, 439–447 (2003)
Belachew, S. et al. Postnatal NG2 proteoglycan-expressing progenitor cells are intrinsically multipotent and generate functional neurons. J. Cell Biol. 161, 169–186 (2003)
Huang, J. K. et al. Retinoid X receptor gamma signaling accelerates CNS remyelination. Nature Neurosci. 14, 45–53 (2011)
Gensert, J. M. & Goldman, J. E. Endogenous progenitors remyelinate demyelinated axons in the adult CNS. Neuron 19, 197–203 (1997)
Horner, P. J. et al. Proliferation and differentiation of progenitor cells throughout the intact adult rat spinal cord. J. Neurosci. 20, 2218–2228 (2000)
Kremer, D., Aktas, O., Hartung, H. P. & Kury, P. The complex world of oligodendroglial differentiation inhibitors. Ann. Neurol. 69, 602–618 (2011)
Patel, J. R. & Klein, R. S. Mediators of oligodendrocyte differentiation during remyelination. FEBS Lett. 585, 3730–3737 (2011)
Chang, A., Tourtellotte, W. W., Rudick, R. & Trapp, B. D. Premyelinating oligodendrocytes in chronic lesions of multiple sclerosis. N. Engl. J. Med. 346, 165–173 (2002)
Chari, D. M., Huang, W. L. & Blakemore, W. F. Dysfunctional oligodendrocyte progenitor cell (OPC) populations may inhibit repopulation of OPC depleted tissue. J. Neurosci. Res. 73, 787–793 (2003)
Wolswijk, G. Chronic stage multiple sclerosis lesions contain a relatively quiescent population of oligodendrocyte precursor cells. J. Neurosci. 18, 601–609 (1998)
Kuhlmann, T. et al. Differentiation block of oligodendroglial progenitor cells as a cause for remyelination failure in chronic multiple sclerosis. Brain 131, 1749–1758 (2008)
Hart, I. K., Richardson, W. D., Bolsover, S. R. & Raff, M. C. PDGF and intracellular signaling in the timing of oligodendrocyte differentiation. J. Cell Biol. 109, 3411–3417 (1989)
Billon, N., Tokumoto, Y., Forrest, D. & Raff, M. Role of thyroid hormone receptors in timing oligodendrocyte differentiation. Dev. Biol. 235, 110–120 (2001)
Tokumoto, Y. M., Tang, D. G. & Raff, M. C. Two molecularly distinct intracellular pathways to oligodendrocyte differentiation: role of a p53 family protein. EMBO J. 20, 5261–5268 (2001)
Fernandez, M. et al. Thyroid hormone administration enhances remyelination in chronic demyelinating inflammatory disease. Proc. Natl Acad. Sci. USA 101, 16363–16368 (2004)
Calzà, L., Fernandez, M. & Giardino, L. Cellular approaches to central nervous system remyelination stimulation: thyroid hormone to promote myelin repair via endogenous stem and precursor cells. J. Mol. Endocrinol. 44, 13–23 (2010)
Barres, B. A., Lazar, M. A. & Raff, M. C. A novel role for thyroid hormone, glucocorticoids and retinoic acid in timing oligodendrocyte development. Development 120, 1097–1108 (1994)
Buckley, C. E. et al. Drug reprofiling using zebrafish identifies novel compounds with potential pro-myelination effects. Neuropharmacology 59, 149–159 (2010)
Ibanez, C. et al. Steroids and the reversal of age-associated changes in myelination and remyelination. Prog. Neurobiol. 71, 49–56 (2003)
Baer, A. S. et al. Myelin-mediated inhibition of oligodendrocyte precursor differentiation can be overcome by pharmacological modulation of Fyn-RhoA and protein kinase C signalling. Brain 132, 465–481 (2009)
Joubert, L. et al. Chemical inducers and transcriptional markers of oligodendrocyte differentiation. J. Neurosci. Res. 88, 2546–2557 (2010)
Gard, A. L. & Pfeiffer, S. E. Two proliferative stages of the oligodendrocyte lineage (A2B5+O4−and O4+GalC−) under different mitogenic control. Neuron 5, 615–625 (1990)
Pfeiffer, S. E., Warrington, A. E. & Bansal, R. The oligodendrocyte and its many cellular processes. Trends Cell Biol. 3, 191–197 (1993)
Gaspard, N. et al. Generation of cortical neurons from mouse embryonic stem cells. Nature Protocols 4, 1454–1463 (2009)
Barres, B. A. et al. Cell death and control of cell survival in the oligodendrocyte lineage. Cell 70, 31–46 (1992)
Eshleman, A. J., Henningsen, R. A., Neve, K. A. & Janowsky, A. Release of dopamine via the human transporter. Mol. Pharmacol. 45, 312–316 (1994)
McKearney, J. W. Stimulant actions of histamine H1 antagonists on operant behavior in the squirrel monkey. Psychopharmacol. 77, 156–158 (1982)
Agoston, G. E. et al. Novel N-substituted 3α-[bis(4'-fluorophenyl)methoxy]tropane analogues: selective ligands for the dopamine transporter. J. Med. Chem. 40, 4329–4339 (1997)
De Angelis, F., Bernardo, A., Magnaghi, V., Minghetti, L. & Tata, A. M. Muscarinic receptor subtypes as potential targets to modulate oligodendrocyte progenitor survival, proliferation and differentiation. Dev. Neurobiol. 72, 713–728 (2012)
Stidworthy, M. F. et al. Notch1 and Jagged1 are expressed after CNS demyelination, but are not a major rate-determining factor during remyelination. Brain 127, 1928–1941 (2004)
Taveggia, C., Feltri, M. L. & Wrabetz, L. Signals to promote myelin formation and repair. Nature Rev. Neurol. 6, 276–287 (2010)
Ragheb, F. et al. Pharmacological and functional characterization of muscarinic receptor subtypes in developing oligodendrocytes. J. Neurochem. 77, 1396–1406 (2001)
Felder, C. C. Muscarinic acetylcholine receptors: signal transduction through multiple effectors. FASEB J. 9, 619–625 (1995)
Owens, T. & Sriram, S. The immunology of multiple sclerosis and its animal model, experimental allergic encephalomyelitis. Neurol. Clin. 13, 51–73 (1995)
Lawson, B. R. et al. Inhibition of transmethylation down-regulates CD4 T cell activation and curtails development of autoimmunity in a model system. J. Immunol. 178, 5366–5374 (2007)
Mix, E., Meyer-Rienecker, H. & Zettl, U. K. Animal models of multiple sclerosis for the development and validation of novel therapies—potential and limitations. J. Neurol. 255 (Suppl 6). 7–14 (2008)
Steinman, L. & Zamvil, S. S. Virtues and pitfalls of EAE for the development of therapies for multiple sclerosis. Trends Immunol. 26, 565–571 (2005)
Aharoni, R. et al. Distinct pathological patterns in relapsing-remitting and chronic models of experimental autoimmune enchephalomyelitis and the neuroprotective effect of glatiramer acetate. J. Autoimmun. 37, 228–241 (2011)
Liu, L. et al. Myelin repair is accelerated by inactivating CXCR2 on nonhematopoietic cells. J. Neurosci. 30, 9074–9083 (2010)
Mi, S. et al. LINGO-1 antagonist promotes spinal cord remyelination and axonal integrity in MOG-induced experimental autoimmune encephalomyelitis. Nature Med. 13, 1228–1233 (2007)
Kawashima, K. & Fujii, T. Basic and clinical aspects of non-neuronal acetylcholine: overview of non-neuronal cholinergic systems and their biological significance. J. Pharmacol. Sci. 106, 167–173 (2008)
Stern, J. N. et al. Promoting tolerance to proteolipid protein-induced experimental autoimmune encephalomyelitis through targeting dendritic cells. Proc. Natl Acad. Sci. USA 107, 17280–17285 (2010)
Steelman, A. J., Thompson, J. P. & Li, J. Demyelination and remyelination in anatomically distinct regions of the corpus callosum following cuprizone intoxication. Neurosci. Res. 72, 32–42 (2012)
Matsushima, G. K. & Morell, P. The neurotoxicant, cuprizone, as a model to study demyelination and remyelination in the central nervous system. Brain Pathol. 11, 107–116 (2001)
Kappos, L. et al. Oral fingolimod (FTY720) for relapsing multiple sclerosis. N. Engl. J. Med. 355, 1124–1140 (2006)
Durelli, L. et al. Every-other-day interferon beta-1b versus once-weekly interferon beta-1a for multiple sclerosis: results of a 2-year prospective randomised multicentre study (INCOMIN). Lancet 359, 1453–1460 (2002)
Noronha, A., Toscas, A. & Jensen, M. A. Interferon β decreases T cell activation and interferon γ production in multiple sclerosis. J. Neuroimmunol. 46, 145–153 (1993)
Brinkmann, V. et al. The immune modulator FTY720 targets sphingosine 1-phosphate receptors. J. Biol. Chem. 277, 21453–21457 (2002)
Kondo, T. & Raff, M. The Id4 HLH protein and the timing of oligodendrocyte differentiation. EMBO J. 19, 1998–2007 (2000)
Wang, S., Sdrulla, A., Johnson, J. E., Yokota, Y. & Barres, B. A. A role for the helix-loop-helix protein Id2 in the control of oligodendrocyte development. Neuron 29, 603–614 (2001)
Pang, Y. et al. Neuron-oligodendrocyte myelination co-culture derived from embryonic rat spinal cord and cerebral cortex. Brain Behav. 2, 53–67 (2012)
Izrael, M. et al. Human oligodendrocytes derived from embryonic stem cells: Effect of noggin on phenotypic differentiation in vitro and on myelination in vivo. Mol. Cell. Neurosci. 34, 310–323 (2007)
Acknowledgements
This work was supported by the Skaggs Institute for Chemical Biology and the California Institute for Regenerative Medicine grant TR3-05617 (to P.G.S), the California Institute for Regenerative Medicine (TG2-01165) and National Science Foundation pre-doctoral fellowships (to V.A.D and C.A.L, respectively). We are grateful to T. Hasnat, M. Chadwell, W. Kiosses and M. Wood for technical support. This is manuscript number 21786 of The Scripps Research Institute.
Author information
Authors and Affiliations
Contributions
L.L.L., P.G.S., C.A.L. and V.A.D. initiated the project and developed strategy. V.A.D., V.T., C.C.G., B.K., H.J.K., K.P., J.G.S. and I.A. performed the experiments. L.L.L., P.G.S. and V.A.D.wrote the manuscript. B.R.L., C.A.L., A.N.T., F.H.G. and T.K. contributed essential ideas and comments.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Extended data figures and tables
Extended Data Figure 1 High-throughput screen to identify inducers of OPC differentiation.
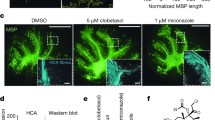
a, Rat primary OPCs in basal differentiation media treated with DMSO (<0.1%) or thyroid hormone (T3; 1 μM) for 6 days in culture, fixed and stained using antibodies for myelin basic protein (MBP), 2′,3′-cyclic-nucleotide 3′-phosphodiesterase (CNP) and oligodendrocyte marker O4. A2B5+ OPCs differentiated into immature oligodendrocytes that express CNP and O4, but not MBP, upon reduction of PDGF-AA. T3 added as a positive control induced differentiation to mature cells that express MBP. Scale bars, 100 μm. b, Schematic representation of the high-throughput screening platform used to identify inducers of OPC differentiation. c, Inducers of OPC differentiation identified as hits from a screen of known biologically active compounds. Scale bars, 100 μm; inset, 40 μm.
Extended Data Figure 2 Benztropine induces dose-dependent OPC differentiation in vitro to mature oligodendrocytes.
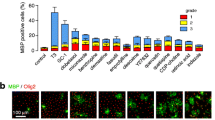
a, Dose response assay used to confirm primary screening activity of benztropine and determine potency (EC50). OPCs were treated with benztropine and immunostained using antibodies for MBP (n = 3, mean and s.d.). b, Images showing dose-dependent induction of OPC differentiation after treatment with benztropine. (Scale bars, 100 μm; inset, 40 μm). OPCs in basal differentiation media treated with DMSO (<0.1%), T3 (1 μM) or benztropine (1.5 μM) for 6 days and analysed for MBP and MOG expression by western blot (c) and by qRT–PCR (d) (n = 3, mean and s.d.). e, OPCs were plated in differentiation medium and treated with DMSO (<0.1%), benztropine (1.5 μM) or T3 (1 μM) for 6 days. Cells were fixed and immunostained for myelin basic protein (MBP), myelin oligodendroglial glycoprotein (MOG), CNP, oligodendrocyte marker O1, oligodendrocyte marker O4, glial marker SOX10, proteolipid peptide (PLP), OLIG1 and OLIG2. Representative images showing expression of mature oligodendrocyte markers in benztropine- and T3-treated cells, but not DMSO-treated cells. Scale bars, 100 μm; inset, 40 μm. f, Expression of cell cycle genes by qRT–PCR. (n = 3, mean and s.d., *P < 0.05, t-test). g, OPCs plated in basal differentiation medium and treated with benztropine (1.5 μM) on various days (0, 1, 2, 3, 4 and 5), fixed on day 6 and immunostained for MBP (n = 3, mean and s.d.). h, OPCs plated in basal differentiation medium, treated with benztropine (1.5 μM) on the same day, fixed on various days (3, 4, 5 and 6) following compound treatment and immunostained for MBP (n = 3, mean and s.d.).
Extended Data Figure 3 Benztropine induces OPC differentiation and in vitro myelination through M1/M3 muscarinic receptor antagonism and has no effect on histamine or nicotinic signalling.
a, Mouse OPCs co-cultured with mouse cortex-derived cells in the presence of DMSO or benztropine and immunostained for MBP (red) and nuclei with Hoechst 33342 (blue). Scale bars, 100 μm. b, Quantification of MBP staining of mouse OPCs treated with DMSO or benztropine. c, Analysis of myelination in OPCs with neurons co-culture. Arrowheads point to regions of myelination. Scale bars, 20 μm. d, Quantification of fraction of myelinating oligodendrocytes in OPCs with neurons co-cultures (n = 10, mean and s.e.m., **P < 0.01, ANOVA with Bonferroni correction). OPCs co-treated with benztropine (1.5 μM) and carbachol (2.3 μM) for 6 days and stained for MBP (green) (Scale bars, 100 μm; inset, 40 μm). e, Antagonism of benztropine-induced OPC differentiation by muscarinic agonist carbachol. f, Quantification of MBP staining of OPCs co-treated with benztropine (1.5 μM) and muscarinic receptor agonist carbachol for 6 days under basal differentiation conditions (n = 3, mean and s.d.). g–k, OPCs plated co-treated with benztropine (0.8 μM) and either nicotine (g), histamine (h), histamine receptor agonist histamine trifluoromethyl toluidide (HTMT) (i), dopamine receptor agonist quinpirole (j) or dopamine receptor antagonist haloperidol (k) (n = 3, mean and s.d., ns = not significant). l, Various nicotinic receptor antagonists have no effect on OPC differentiation. m, Benztropine blocks carbachol- and muscarine-induced activation of Notch signalling measured by western blot for intracellular domain of Notch1. (a.u., arbitrary unit, n = 3, mean and s.d., *P < 0.05, t-test). n, Naive whole rat brain and rat primary OPCs treated with DMSO (<0.1%) or T3 (1 μM) for 6 days tested for expression of muscarinic receptors and choline acetyl transferase (ChAT) by PCR using gene-specific primers. o, Quantification of M1, M2, M3, M4 and ChAT expression by qRT-PCR. (n = 3, mean and s.d., expression fold change normalized to OPCs). p, OPCs treated with benztropine (25 μM) and pelleted for western blot analysis of total protein. q, Carbachol induced a dose-dependent increase in intracellular Ca2+ levels, whereas benztropine and atropine (a muscarinic antagonist) dose-dependently blocked carbachol (50 μM) induced calcium influx through antagonism of M1/M3 muscarinic receptors. r, Benztropine (13 μM) had no effect on the levels of cAMP. Forskolin is a positive control for increasing intracellular cAMP (n = 3, mean and s.d., *P < 0.05, t-test).
Extended Data Figure 4 Benztropine dose-dependently reduces clinical severity and induces remyelination in the PLP-induced EAE model.
a, Clinical severity scores of EAE mice treated with various doses of benztropine in the prophylactic mode (n = 8, mean and s.e.m.). b, EAE mice treated with benztropine (10 mg per kg) or vehicle in the therapeutic mode and spinal cord sections from mice representative of the average group scores during the relapse phase of EAE stained with Luxol fast blue and H&E, or Luxol fast blue only. Arrows point to regions of lymphocyte infiltration (LFB + H&E) or demyelination (LFB). Scale bars represent 100 μm. EAE mice treated with benztropine (10 mg per kg) or vehicle in prophylactic mode. c, Spinal cord sections from mice representative of the average group scores on day 8, 11 and 14 immunostained with antibodies specific to CD45 and GSTπ. d, Mean clinical scores of mice at the time of spinal cord isolation and quantification of the infiltrated areas (CD45+) and number of GSTπ+ cells (n = 8, mean and s.e.m., **P < 0.01, t-test). Scale bars, 100 μm. e, EAE mice treated with benztropine (10 mg per kg) or vehicle in prophylactic mode and spinal cord sections from mice representative of the average group scores on day 11 and 14, immunostained with antibody specific to MBP. Arrows point to regions of lymphocyte infiltration. Scale bars, 100 μm. f, Electron microscopy images showing myelin around axons in normal mice, vehicle-treated mice and mice in remission. Scale bars as indicated. g, Analysis of electron microscopy images indicating distribution of axonal diameters measured for 4 groups. h, Analysis of electron microscopy images indicating distribution of g-ratios of axons for 4 groups. i, Scatterplot of g-ratios in relation to spinal cord axonal diameters (n = 1,000, ***P < 0.001, one-way ANOVA, exponential trend line). j, Quantification of the number of axons associated with oligodendrocytes (n = 25, mean and s.e.m., **P < 0.01, t-test). Oligodendrocytes were identified visually by their cytoplasmic processes wrapping around axons.
Extended Data Figure 5 Benztropine has no effect on in vitro and in vivo immunological responses in EAE mice.
a, Benztropine and various muscarinic antagonists have no effect on in vitro T-cell proliferation measured using carboxyfluorescein succinimidyl ester (CFSE) labelling, whereas mycophenolate and FTY720 suppress T-cell proliferation as determined by the percentage of CD4+ T-cell-gated populations positive for the given marker. b, c, Various muscarinic antagonists have no effect on T-cell activation as measured by CD4+CD25+, CD4+CD69+, CD8+CD25+ and CD8+CD69+ cell populations. FTY720 and mycophenolate serve as positive controls for suppression of T-cell activation. d, Representative flow cytometry scatter plots show similar numbers of CD4+, CD8+, and CD44Hi cells in spleens isolated from vehicle- and benztropine-treated mice. e, f, Total splenocytes isolated from benztropine (10 mg per kg) or vehicle treated (14 days in the prophylactic mode) naive SJL/J (e) or EAE (f) mice analysed for various populations of immune cells and cytokine secretion. Benztropine treatment had no effect on the numbers of total splenocytes, CD4+ T cells, CD8+ T cells, CD4+CD44Hi T cells and CD8+CD44Hi T cells. Benztropine treatment showed a minor, but significant decrease in the number of B cells (n = 5, mean and s.e.m., *P < 0.05, t-test). Benztropine had no effect on cytokine production from CD4+ T cells expressing IL-2, IL-10, TNF-α or IFN-γ. (n = 5, mean and s.e.m.). g, Benztropine showed no effect on keyhole limpet hemocyanin protein conjugated to 2,4,6-trinitrophenyl hapten (TNP-KLH)-induced T-cell-dependent B-cell response. Mice were injected with TNP-KLH in adjuvant and treated with vehicle or benztropine (10 mg per kg) daily. Serum was isolated at various time points and anti-TNP-IgG levels were measured by ELISA. (3 replicate ELISAs, n = 5 mice per group, mean and s.e.m.).
Extended Data Figure 6 Benztropine does not affect derivation and in vitro polarization of macrophages from bone marrow derived monocytes.
a, Flow cytometry analysis of bone marrow derived monocytes treated in vitro with either DMSO (<0.1% v/v) or benztropine (5 µM) for 24 h followed by 24 h treatment with LPS (100 ng ml−1) plus IFNγ (20 ng ml−1) for the expression of M1 markers: CD86, MHC-II and CD80, or 24 h treatment with IL-4 plus IL-13 (20 ng ml−1 each) for the expression of M2 marker CD206. b, M1/M2 polarized macrophages re-stimulated using either LPS (100 ng ml−1) plus IFNγ (20 ng ml−1) (M1) or IL-4 plus IL-13 (20 ng ml−1 each) (M2) for 16 h in the presence of either benztropine (5 μM) or DMSO and analysed for the expression of M1 (CD80) or M2 (CD206) markers by flow cytometry. c, d, Treatment with LPS (100 ng ml−1) plus IFNγ (20 ng ml−1) induced the expression of the prototypical M1 cytokine TNF-α as detected by intracellular flow cytometry (c) and ELISA (d) with no significant differences between DMSO or benztropine (5 µM) treated cells (data representative of 2 replicate experiments).
Extended Data Figure 7 Benztropine does not affect in vivo polarization of macrophages in the spleen or spinal cord.
EAE mice were treated with benztropine (10 mg per kg) or vehicle for 14 days in the prophylactic mode. a, Mean clinical EAE scores for mice treated with vehicle or benztropine (n = 6, mean and s.e.m. for spleens and spinal cords, n = 12 for isolated spinal leukocytes analysis). b–e, Spleens and spinal leukocytes were isolated from the mice as described in Methods. Total RNA was isolated, reverse transcribed and gene expression was measured by qRT–PCR. Expression for each marker was normalized to the average gene expression of the vehicle group. No significant differences were observed in the expression of markers of macrophage polarization in the spleen (b), whole spinal cords (c) and leukocytes (d, e) isolated from spinal cords (n = 6 mice per group for spleens and spinal cords, n = 12 mice per group (n = 6 for qRT–PCR) for spinal leukocytes analysis. Error bars represent s.e.m.).
Extended Data Figure 8 Benztropine does not affect clinical severity in an adoptive transfer model of EAE.
a, b, Incidence of adoptive transfer of EAE (a) and mean clinical EAE scores (b) in mice injected with splenocytes isolated from benztropine- or vehicle-treated donor groups. T cells obtained from either benztropine- (BT, 10 mg per kg) or vehicle-treated donor EAE mice and further expanded in the presence or absence of benztropine (5 μM) were able to adoptively transfer EAE to naive recipient mice. Benztropine-treated recipient mice showed little to no clinical symptoms of EAE compared to vehicle-treated recipient mice, whether injected with benztropine- or vehicle-treated donor splenocytes (n = 6 mice, mean and s.e.m., *P < 0.05, t-test). c, Schematic for the adoptive transfer EAE model. d, Table showing various groups and treatments. e, ELISA for anti-PLP IgG shows equivalent PLP response in donor mice treated with either vehicle or benztropine (n = 30, mean and s.e.m).
Extended Data Figure 9 Quantification of myelin staining in the cuprizone model.
a, Luxol fast blue (LFB) and H&E staining was performed on sections from the corpus callosum region of brains isolated from mice treated either with benztropine (10 mg per kg) or vehicle after 7 weeks of exposure to cuprizone. b, Images were converted to a 256 shade grey scale. c, The 256 shades of grey were divided into 5 bins of 50 shades each (1–50, 51–100, 105–150, 151–200 and 201–256). Number of objects in the corpus callosum region in each bin were counted using Image-Pro plus. d, Representative images of Image-Pro rendering of the quantification of objects in each bin. e, Quantification of Luxol fast blue staining on week 2 shows an increase in the darker pixels (1–50 and 51–100) with benztropine treatment along with corresponding reduction in the number of lighter pixels (151–200). Six images per mouse were analysed and four mice per group were used at each time point (mean and s.d., *P < 0.05, t-test). Scale bars, 200 μm.
Extended Data Figure 10 Effect of the addition of benztropine to interferon-β and FTY720 treatments.
a, b, EAE severity scores for mice treated with various doses of FTY720 (a) or interferon-β (b). c, Mice treated therapeutically with FTY720 (1 mg per kg) in combination with a sub-optimal dose of benztropine (BT, 2.5 mg per kg) show significantly decreased clinical severity compared to FTY720 (1 mg per kg) or benztropine (2.5 mg per kg) alone. d, EAE mice treated with interferon-β (IFN 10,000 U per mouse) in combination with benztropine (2.5 mg per kg) show significantly decreased clinical severity compared to interferon-β (IFN; 10,000 U per mouse) or benztropine (2.5 mg per kg) alone. e, EAE mice treated with a tenfold lower dose of FTY720 (0.1 mg per kg) in combination with benztropine (2.5 mg per kg). f, EAE mice treated with a tenfold lower dose of FTY720 (0.1 mg per kg) in combination with benztropine (2.5 mg per kg) show clinical severity comparable to optimal dose of FTY720 (1 mg per kg) (n = 8 mice per group, mean and s.e.m., *P < 0.05; t-test). g, i, Spinal cord sections from EAE mice treated with the indicated drug(s) for 14 days in the prophylactic mode and immunostained for CD45 (immune cells) and GSTπ (oligodendrocytes) showing infiltration (g) and oligodendrocytes (i). h, j, Quantification of the number of CD45+ (h) and GSTπ+ (j) cells showing a decrease in infiltrating cells with FTY720 treatment and an increase in oligodendrocytes numbers with benztropine treatment and synergy between benztropine (2.5 mg per kg) and FTY720 (0.1 mg per kg) (n = 5, mean and s.e.m., ns, not significant). Scale bars, 100 μm. k, Mean clinical EAE scores for mice at the time of spinal cord isolation (n = 8, mean and s.e.m).
Supplementary information
Supplementary Tables
This file contains Supplementary Tables 1-5. (PDF 321 kb)
Rights and permissions
About this article
Cite this article
Deshmukh, V., Tardif, V., Lyssiotis, C. et al. A regenerative approach to the treatment of multiple sclerosis. Nature 502, 327–332 (2013). https://doi.org/10.1038/nature12647
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/nature12647
- Springer Nature Limited
This article is cited by
-
The aminosteroid U73122 promotes oligodendrocytes generation and myelin formation
Acta Pharmacologica Sinica (2024)
-
Norepinephrine modulates calcium dynamics in cortical oligodendrocyte precursor cells promoting proliferation during arousal in mice
Nature Neuroscience (2023)
-
Artificial axons as a biomimetic 3D myelination platform for the discovery and validation of promyelinating compounds
Scientific Reports (2023)
-
Cholinergic control of Th17 cell pathogenicity in experimental autoimmune encephalomyelitis
Cell Death & Differentiation (2023)
-
Inflammation in multiple sclerosis: consequences for remyelination and disease progression
Nature Reviews Neurology (2023)