
Abstract
Cannibalism is a common phenomenon that occurs in over 30 fish families. Most of the species are valuable objects of aquaculture and cannibalism is highly undesirable. Cannibalism is an aggressive behaviour, which can be caused by stress induced by various population and environmental factors and can occur in two main forms: early larval and late juvenile (or adult). Each of these forms can lead to severe losses of stock. Losses are caused not only by fish eating each other, but also by an increase in the number of infections (the result of body surface damage). It is important to understand the phenomenon of cannibalism in order to properly prevent, diagnose and reduce it. The main aim of this study was to present and examine some of the well-examined methods of cannibalism mitigation. Most of the methods consist of creating rearing conditions similar to natural conditions and to reduce stress. Moreover, because of large interspecies differences, similar methods can result in different effects depending on the species. Additional studies are needed to better understand the biology of each cultured species with particular emphasis on the impact of domestication, as well as to improve fish rearing conditions.
Similar content being viewed by others


Introduction
Cannibalism is a predatory feeding strategy that involves killing and eating individuals of the same species. In nature, this phenomenon can occur between parents and offspring, in a given age group between siblings or between individuals from separate populations (regardless of the kinship) (Smith and Reay 1991; Baras and Jobling 2002; Qin et al. 2004 and see Table 1). This behaviour represents a specific protective measure against an excessive density of a given population and an alternative source of food when other food resources become scarce (van den Bosch et al. 1988).
Cannibalism is reported in both wild and farmed fish, particularly in aquaculture of predatory fish. This behaviour may reflect disrupted welfare, poorly adapted rearing conditions (especially with regard to feeding schedules or modes of food distribution), or unmet behavioural needs in recently domesticated species (Kaleta 2013). Another possible cause of cannibalism may be the establishment of the group hierarchy among old larvae and subsequent stages of ontogeny (Smith and Reay 1991). This phenomenon is associated not only with a dramatic decline in fish survival, but also in injuries in situations when the prey is too large to be totally consumed by a cannibal. Furthermore, cannibalism generates substantial economic losses (Baras and Jobling 2002; Kestemont et al. 2003; Qin et al. 2004; Król and Zieliński 2015).
The objective of this paper was to review the current knowledge of cannibalism in freshwater fish reared in aquaculture, including clinical aspects for veterinarians involved in the examination of fish. We also review the methods applied to mitigate the prevalence of cannibalism.
Causes, occurrence and development
Cannibalism in cultured fish occurs in different age groups and its type and development depend mostly on the species and production technology. It represents a major problem in the commercial production of many fish species (especially predatory species) and ranges from 15% to over 90% of individuals (Hecht and Appelbaum 1988). This phenomenon is mainly observed in fish from the following families: airbreathing catfish (Clariidae), pikes (Esocidae), percids (Percidae), characids (Characidae), latids (Latidae), gadids (Gadidae) and in over 30 other families, including cyprinids (Cyprinidae) and salmonids (Salmonidae), (Smith and Reay 1991; Hecht and Pienaar 1993; Qin et al. 2004).
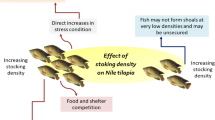
Under rearing conditions, the intensity of cannibalism is directly influenced by population and abiotic factors, along with genetic predispositions (Fig. 1). However, the extent to which cannibalistic behaviour is genetically controlled in fish remains controversial, and it is still unclear if cannibals in many species are “natural born killers” or just “lottery winners” (Baras and Jobling 2002; Król et al. 2013; Yang et al. 2015). Some studies have demonstrated that this phenomenon is to some extent genetically determined (Thibault 1974; Ferguson and Noakes 1983; van Damme et al. 1989; Amundsen et al. 1999; Yang et al. 2015). However, the interactions between genotype and environment play a major role in the expression of cannibalism (Baras and Jobling 2002; Yang et al. 2015). Intracohort cannibalism may also be sex-based, especially in species that show sexual growth dimorphism during early ontogeny, e.g. Vundu catfish (Heterobranchus longifilis) (Baras 1999a), or in species with diet dimorphism, e.g. old hake females (family Gadidae) (Garrison and Link 2000). Moreover, the influence of the genotype of a maternal parent on cannibalism in progeny is higher than the paternal one (Mousseau 1998; Yang et al. 2015).

Population and environmental factors influencing cannibalism
Population factors that influence cannibalism include the size and density of larvae, with both being positively correlated with the incidence of cannibalistic behaviour. Abiotic factors mainly encompass the availability and energy value of feed, diet supplementation, feeding schedule, temperature and clarity of water, light intensity, photoperiod, shape and colour of internal walls of rearing tanks or the lack of refuges for small-sized individuals (Smith and Reay 1991; Britz and Pienaar 1992; Hecht and Pienaar 1993; Folkvord and Otterå 1993; Baras et al. 2000a; Baras and Jobling 2002; Kestemont et al. 2003; Szczepkowski 2009; Król and Zieliński 2015). As all factors interact with each other, they should be approached holistically (Hecht and Pienaar 1993; Kestemont et al. 2003) in order to mitigate cannibalism. These factors are discussed further in “Methods used to mitigate cannibalistic behaviour” section.
Intracohort cannibalism is typical of farming conditions and is divided into two types: type I, the so-called “early” type, which occurs in the larval phase and is independent of a diversity in fish sizes, where the victim is not completely ingested or consumed; and type II, a later form associated with heterogeneous growth, when the victim is consumed whole (Hecht and Appelbaum 1988; Smith and Reay 1991; Baras and Jobling 2002). A potential type I cannibal mostly attacks its prey from the tail end, eats only a damaged part of the tail and disgorges the rest of the larva as it is unable to swallow it due to the lack of difference in size. This does not affect some species, i.e. those with very large mouth dimensions or sharp oral teeth in the early ontogenetic stage—in such situations, cannibals can attack siblings’ heads or trunks (Hecht and Appelbaum 1988; Baras 1999b; Baras and Jobling 2002). Type II cannibalism is determined by the mouth dimensions of a potential cannibal in relation to a potential prey, particularly to its body depth (Hecht and Appelbaum 1988; Smith and Reay 1991; Sogard and Olla 1994). The gape size limitations differ between species and ontogenetic stages, owing to the allometric growth of mouthparts (Baras and Jobling 2002). For type II cannibalism, prey is attacked from the head end or from the side and usually swallowed whole (Baras and Jobling 2002; Kestemont et al. 2003). However, because of a large predator-to-prey ratio, in type II cannibalism the mode of ingestion may be of little importance (Baras and Jobling 2002). An exception for this is Acanthopterygians, where prey is eaten head first as a cannibal then avoids the spiny fin rays of the dorsal or pectoral fins which threaten a predator. Otherwise, the erected rays may cause the death of the cannibal and its prey (Brabrand 1995; Qin et al. 2004). In most fish species, type I cannibalism is replaced by type II together with increasing gape size limitations, heterogeneity of a stock and growing chances of swallowing a potential prey. Type I cannibalism may trigger growth heterogeneity because digestible nutrients are often higher in fish than in plankton prey, which leads to the growth advantage of cannibals and, in effect, to the occurrence of type II cannibalism (Baras and Jobling 2002; Król and Zieliński 2015).
The outcomes of these two types of cannibalism differ in the level of mortality. In the case of type I, it ranges between 1.5 and 12.0% of the initial stock of European perch (Perca fluviatilis) larvae (Baras et al. 2003; Babiak et al. 2004; Król and Zieliński 2015) and from 10 to 17% in the stock of pikeperch (Sander lucioperca) larvae (Zakęś and Demska-Zakęś 1996; Zakęś 2012; Król and Zakęś 2016) or even 40% in dorada (Brycon moorei) (Baras et al. 2000b). Type II usually generates significantly higher losses and is mainly reported in a stock characterised by a large heterogeneity in size (Król and Zieliński 2015).
Hormonal background
Intracohort aggression has often been linked to the mortality of aquaculture organisms (Manley et al. 2015). It is hormonally regulated via different modulators (Table 2), and is the basis of cannibalistic behaviour (Hecht and Appelbaum 1988; Folkvord 1997; Yang et al. 2015; Vallon et al. 2016).
In some species or life stages, an organism also may be aggressive or cannibalistic toward a conspecific when defending its own space (Hecht and Pienaar 1993; Baras and Jobling 2002), fighting for food during low feeding frequency and periods of low food availability, when conspecifics offer the only type of alternative prey (Hecht and Pienaar 1993; Baras and Jobling 2002; Kestemont et al. 2003). However, there are also exceptions, for example, in Vundu catfish, type II cannibalism requires no preliminary aggression or chase, or even contact with the prey, suggesting that cannibalism can be independent of aggressive behaviour (Baras 1999b).
Diagnostics
Due to the complex causes of cannibalism, diagnostics should always start with case history—gathering information from the owner of the fish farm, which should be as extensive as possible. The questions should include the exact percentage of losses with a particular interest in the age of fish and the diversity in growth. It is important to consider how the losses were spread among a given age group and individual tanks. In order to rule out welfare issues, water parameters should be analysed in the preceding period (at least 2 weeks). Such a procedure enables detection of possible situations, which may trigger stress and behavioural changes, for example, alterations in temperature, oxygen or pH level, power outages, bio-filter faults or excessive waste. The owner should also be asked about situations that may have disrupted the established hierarchy, namely, transfer or sorting fish and tank cleaning. Data on nutrition is equally important; they refer to both feed types and feeding strategies.
The next step is a physical examination which includes a thorough inspection of fish with special attention paid to any injuries (inspection with a magnifying glass, a microscopic examination under a small magnification) and their localization. This should help to exclude the injuries caused by handling, tank design or coexisting diseases. In the case of tail injuries, type I cannibalism is suspected while the wounds to the head and body sides may indicate type II cannibalism. In addition, the gastrointestinal content of a potential cannibal may be examined. In most species, fish of different sizes are differentially sensitive to handling, but handling generally has no significant effect on fish survival (Baras et al. 2011a). However, special attention is needed when very small and fragile Percids larvae are examined, as they are by nature very sensitive to any handling and parameter alterations, which results in deaths (Król and Zieliński 2015) and may be mistaken for an outcome of cannibalism.
Methods used to mitigate cannibalistic behaviour
Different approaches may be implemented to reduce the incidence of cannibalistic behaviour and resulting mortality. Previous research suggests that applying different methods simultaneously will produce a synergistic effect (Qin et al. 2004) and that methods should be adjusted according to a given species and its biology.
Abiding by density standards
In nature, cannibalism is determined by the population density and usually occurs in years with high numbers of larvae (Baras and Jobling 2002). In aquaculture, excessive density may reduce or increase the incidence of cannibalistic behaviour, depending on the biology of a given species. With limited availability of food or improper feeding techniques (limited access, bullying of weaker individuals), an increased number of larvae is associated with lower resources per larva, which results in using a rescue source of food, i.e. siblings. Under rearing conditions, intensified cannibalistic events due to excessive stock density (despite the correct amount of food) may result from a higher chance of a cannibal-prey interaction (especially in species demonstrating “meet-and-eat” behaviour) (Sogard and Olla 1994; Baras and Jobling 2002).
On the other hand, increasing stock density may reduce aggression, especially that associated with territorial behaviour and setting the dominance hierarchy (Smith and Reay 1991; Sloman and Armstrong 2002). It is suggested that with high densities, a limited surface results in a lower number of aggressive individuals (in reference to the whole population) protecting the territory and demonstrating cannibalistic attitudes (Kucharczyk et al. 1998; Baras and Jobling 2002). In addition, at a certain stock density, fish start forming shoals and cannibals become perceptively confused and cannot select a prey (as the number of potential prey is too high) (Baras and Jobling 2002).
The impact of different stock density levels has been studied in many cultured fish species. A proportional increase in cannibalistic behaviour resulting from rising stock density and an increase in mortality, as a result, has been observed, for instance, in pike (Esox lucius) (Kucharczyk et al. 1998), pikeperch (Szkudlarek and Zakȩś 2007), European perch (Baras et al. 2003) and African sharptooth catfish (Clarias gariepinus) larvae (Solomon and Udoji 2011). For these species, it is suggested to rear larvae initially at higher stock densities and then to reduce the stock proportionally (Kestemont et al. 2003). An opposite relation is seen in such species as Atlantic cod (Gadus morhua) (Baskerville-Bridges and Kling 2000), Vundu catfish (Imorou Toko et al. 2008) or European chub (Leuciscus cephalus) (Żarski et al. 2008), in which there is a reduction in cannibalistic behaviour at high stock densities.
Size-sorting
In commercial farming of many fish species, size-sorting is carried out in order to ensure good husbandry practice, which is economically justified as it allows a more accurate estimation of feed consumption and weight gains in fish (Jobling et al. 1993). On the other hand, size-sorting would also have the effect of disrupting established dominance hierarchies, and it is widely believed that size-sorting results in greater gain in biomass because the suppressive effect of larger fish on the growth of smaller individuals is removed at regular intervals (Jobling et al. 1993). Dominance hierarchies often re-establish soon after fish have been rearranged into new groups, especially when group size is low and the fish are homogenous in size. The establishment of a dominance hierarchy is generally accompanied by heterogeneous food intake and growth, generally to the detriment of the smallest fish, thereby resulting in an overall slower growth rate, greater size dispersal and increased skewness (Azaza et al. 2010).
Size-sorting regarding type I cannibalism is connected mainly with maintaining the ontogenetically homogenous stock, because sorting does not reduce type I cannibalism, as this form is independent of larval sizes. At early stages, intraspecific predation is not greatly limited by size heterogeneity, as cannibals ingest conspecific prey (Ribeiro and Qin 2016). Synchronization of hatching and initial diversity of larval sizes play an important role among many fish species in maintaining a homogenously-sized stock. Both of them are associated with the size of a female and the number of her reproductive seasons. Very small females and those used reproductively for the first time demonstrate a tendency for producing a significantly smaller brood (Król and Zieliński 2015). That is why mixing of progenies with contrasted hatching times (with exception of fish at the same ontogenetic stage and homogenous in size) or progenies from females of different sizes, should be avoided -female size, egg size and size at hatching may be correlated (Wootton 1989; Baras and Jobling 2002; Steenfeldt et al. 2011; Trabelsi et al. 2013; Król and Zieliński 2015; Trabelsi et al. 2016). Therefore, it is recommended to select proper sized females at the stage of spawn collection. This is of particular importance for such species as Atlantic cod (Folkvord and Otterå 1993), African catfish (Hecht and Appelbaum 1988), rainbow trout (Oncorhynchus mykiss) (Baras and Jobling 2002), largemouth bass (Micropterus salmoides) (DeAnglis et al. 1980), walleye (Sander vitreus) (Loadman and Moodie 1986), European seabass (Dicentrarchus labrax) (Katavić et al. 1989) and dorada (Baras et al. 2000a).
In walleye (Cuff 1980) and dorada (Baras et al. 2000b), initial size heterogeneity among embryos presumably has a limited effect on the impact of type I cannibalism. In contrast, the emergence of cannibalism was dependent strictly on the development of oral teeth at the embryonic stage (Baras et al. 2000a, b), suggesting that fish hatching slightly later, or developing at a slightly slower rate than others, incur a major penalty for this very short delay. In particular, this is likely to apply to fish species with oral teeth that facilitate the exercise of incomplete cannibalism (which does not require that the cannibal be larger than its victim), as individuals with a slightly faster growth than others during the first days of exogenous feeding would not be safe from cannibalism (Baras et al. 2014). The size advantage gained through the exercise of type I cannibalism at the embryonic and larval stages, later enables cannibals to exert type II cannibalism over smaller siblings, resulting in high mortality throughout the larval and early juvenile stages (Hecht and Appelbaum 1988; Baras 1999b). The same principle applies to deformed fish, which may constitute easily accessible prey to cannibals, contribute to their faster growth and further predation on normal siblings.
Because of size-dependence of type II cannibalism, substantial research has been done on morphometric assessments of the prevalence of cannibalism depending on the ratio of the mouth-dimensions of cannibals and the body dimensions of victims (Folkvord and Otterå 1993; Mélard et al. 1996; Hseu et al. 2003b; Fessehaye et al. 2005; Mandiki et al. 2007; Ribeiro and Qin 2013). Due to species–dependent morphological differences, the variability of dimensions during ontogeny and behaviour typical of a given species, diverse mathematical models are applied. The most commonly used models include those based on direct measurements of total length (TL) of cannibals and their prey or models based on such parameters as mouth width (MW), mouth gape (MG), body depth (BD) of a prey, head depth (HD) of a prey and head width (HW) of a prey (Folkvord and Otterå 1993; Baras and Dalmeida 2001; Hseu 2002; Kailasam et al. 2002; Kestemont et al. 2003; Hseu et al. 2003b; Fessehaye et al. 2005; Wallat et al. 2005; Hseu et al. 2007; Policar et al. 2013; Baras et al. 2014; Hseu and Huang 2014; Ribeiro and Qin 2015).
Selecting size of prey has been well-described for orange-spotted grouper (Ephinephelus coioides) which can consume prey almost as large as the cannibal. However, they rarely do so and seemingly prefer much smaller prey. Studies have shown that when a cannibal is offered prey of three sizes, orange-spotted grouper cannibals do prefer the smaller prey to the larger ones (Hseu and Huang 2014). Similar situations have been found in other cannibalistic fish such as yellow perch (Perca flavescens) (Post and Evans 1989), Arctic charr (Salvelinus alpinus) (Amundsen et al. 1995), Atlantic cod (Blom and Folkvord 1997), pike (Nilsson 1999), dorada (Baras et al. 2000b), Pangasius djambal (Baras et al. 2010) and Amazonian catfish Pseudoplatystoma punctifer (Baras et al. 2011b).
The size-sorting method based on comparing the length is the simplest and most practical in use, but is applicable only for a few species with direct ontogeny. However, there is some information provided, for example, in barramundi (Lates calcarifer), it is believed that a difference in length of 50% should be regarded as the absolute threshold at which cannibals should be removed (Ribeiro and Qin 2013). In other species, the threshold is 88% for pike (Szczepkowski 2009), 75% for Atlantic cod (Folkvord and Otterå 1993) and 55% for Arctic char (S. alpinus) (Jobling et al. 1993). In the case of Nile tilapia (Oreochromis niloticus), it is recommended to keep the largest individual bigger than the smallest one by a maximum of 13 times (Fessehaye et al. 2005).
In pikeperch, it has been demonstrated that larvae sorting has a beneficial impact on an increase in their survival through reduced cannibalistic behaviour (Szczepkowski et al. 2011). A similar effect of sorting has been reported for turbot (Scophthalmus maximus) (Sunde et al. 1998), African catfish (Biu et al. 2015), and yellow perch (Wallat et al. 2005). However, for the European perch, sorting has a negative impact on cannibalism prevalence and weight gains (Mélard et al. 1996). Arctic charr reared in size-sorted groups also suffer from a growth disadvantage when compared with groups containing fish of a wider range of sizes (Baardvik and Jobling 1990). In these species, removal of the largest individuals is most likely associated with establishing a new hierarchy in a stock, which triggers a reoccurrence of type I cannibalism (Mandiki et al. 2007). In this species, it is worth considering removing the smallest larvae that are the target of cannibals, as such an approach allows levelling the sizes and maintaining the current hierarchy in a stock at the same time (Baras and Jobling 2002).
Balanced nutrition and supplementation
Correct selection of feed, in terms of its physical and chemical characteristics (e.g. size and texture of pellets, drowning speed, shape, colour, hardness, digestibility and beneficial value of FCR), is an important element of the preventive strategy against cannibalistic behaviour (Zakęś and Demska-Zakęś 1996; Altaff and Janakiraman 2013; Król and Zieliński 2015; Ljubobratović et al. 2015). However, rearing period and feeding schedule, specific for each species, are equally important (such as daily feed ration, feeding frequency, feed access surface and transition from live feed to commercial feed) (Jobling et al. 1993; Folkvord and Otterå 1993; Francis and Bengtson 1999; Baras et al. 2000a; Kestemont et al. 2003; Szczepkowski 2009; Conceição et al. 2010; Altaff and Janakiraman 2013; Piotrowska et al. 2013; Król and Zieliński 2015). This is important, because any type of feeding restriction not only promotes forage activities by hunger motivation, encounter rates of conspecifics and prey vulnerability (Hecht and Pienaar 1993; Baras et al. 2013; Ribeiro and Qin 2016), but can also lead to food competition, variability in food intake and establishment of a dominant hierarchy. Every situation which leads to inequities in food intake, results in growth depensation and size heterogeneity (Hecht and Pienaar 1993; Baras and Jobling 2002; Kestemont et al. 2003; Baras et al. 2013; Ribeiro 2015). The combination of all of the above mentioned factors, including the biology of a given species, should satisfy nutritional requirements, support homogenous growth of larvae and eliminate competition for food.
Live feed is usually preferred by fish larvae (Conceição et al. 2010; Ljubobratović et al. 2015). Importantly, it should be adjusted to the requirements and capacities of a given species. Some authors recommend mixed feeding to mitigate cannibalism, for example, in the case of Koi carp (Cyprinus carpio koi), the best performance results (growth and survival ratio) and the least expressed cannibalistic behaviour are reported with feeding rotifer larvae (Brachionus plicatilis) and copepods (Apocyclops dengizicus) (Altaff and Janakiraman 2013). For Atlantic sturgeon larvae (Acipenser oxyrhynchus oxyrhynchus), most often fed with Artemia sp. nauplii, it has been shown that mixing the nauplii with micronematodes of the Panagrellus sp. family is beneficial, as this reduces mortality and incidence of cannibalistic behaviour and is favourable from an economic standpoint (Piotrowska et al. 2013). However, in some cases, mixed feeding may also cause different individuals to feed on different types of food with different nutritional values, which may result in growing size heterogeneity (Baras et al. 2014).
In fish fed with commercial feed from the beginning of rearing period, the addition of live feed may generate rational effects which reduce the prevalence of cannibalism, e.g. Daphnia in the feeding of African catfish and rainbow trout (Hecht and Pienaar 1993). Decreased territorial aggression and cannibalistic behaviour when live feed is supplemented for African catfish and Vundu catfish is most likely caused by redirecting food aggression onto another, actively swimming source of food (Solomon and Udoji 2011). In addition, growth and survival parameters are better in fish fed with mixed feed in comparison to fish fed only with commercial feed, which is related to the differences in the protein-fat ratio (Akbary et al. 2010) and the secretion of additional proteolytic enzymes by the nauplii, which enhances food absorption in larvae (Bengtson et al. 1991). As a result, larvae fed with mixed feed are more efficient in feed conversion and better tolerate the transition to commercial feed.
The weaning from live food to commercial feed is a time point of particular importance for many fish species (Strand et al. 2007). This also applies to feeding perch larvae. When feed is changed, fish should be transitioned throughout a certain period (when both live food and commercial feed are used) long enough to allow larvae to become familiar with a new type of food. Individuals that fail are weaker and smaller and thus become easy prey for potential cannibals (Hecht and Pienaar 1993; Strand et al. 2007). The duration of the individual feeding stages also plays a role. For example, different experiments suggest that the initial phase during which only live food is used, actually slows the growth rate and increases the incidence of deformities in pikeperch (Zakęś and Demska-Zakęś 1996). In addition, excessively long feeding with live food makes fish become used to this type of feed and makes them unwilling to accept a new form of feed, thus triggering stress and a resultant increase in cannibalistic behaviour (Conceição et al. 2010).
Experiments are currently underway that are attempting to determine which feed additives may inhibit the intensity of cannibalism. l-tryptophan is one of them; this amino acid is a serotonin precursor and serotonin exhibits an inhibitory effect on aggressive behaviour via the increased serotoninergic activity of the brain (Winberg et al. 2001). Stressors such as social subordination (a low position in the dominance hierarchy), handling and exposure to predators trigger a rapid activation of the serotoninergic system in the brain (Lepage et al. 2003). Diet supplementation with l-tryptophan impacts the course of stress response in rainbow trout by lowering the level of cortisol in serum. However, with long-term administration, the regulatory mechanisms which limit this effect are switched on (Lepage et al. 2003; Mandiki et al. 2016). The level of absorbed l-tryptophan is positively correlated with the amount of ingested food: the highest doses are taken by the strongest individuals with the ultimate positions in the social hierarchy. Hence, aggressive and cannibalistic behaviours are most effectively suppressed in such animals. Short-term supplementation with l-tryptophan in the diet leads to weakening of the hierarchic structure, lowers the level of social stress, reduces the negative impact of stress on the immunity, levels the sizes of animals and, as a result, reduces the severity of cannibalism (Winberg et al. 2001).
Enrichment of live feed with probiotic preparations is another measure resulting in reduction of cannibalism. The use of probiotics in aquaculture undoubtedly brings about benefits such as growth promotion, increased survival and immunomodulation (Gatesoupe 1999; de Azevedo and Braga 2012). It has been proven that they contribute to a reduction in type II cannibalistic behaviour in the rearing of European perch. This effect was generated by supplementing live feed (Artemia nauplii) with probiotic Bacillus licheniformis, Bacillus subtilis and Bacillus pumilus strains (Mandiki et al. 2011).
Temperature regulation
Environmental temperature has an enormous impact on the metabolic rate in fish, which rises with increasing water temperature (Baras and Jobling 2002; Barron et al. 2012; Frisk et al. 2012). For most fish species, this results in an increased demand for food, faster depletion of food resources, marked heterogeneity of larvae and, as a result, in potential cannibalistic behaviour (Sunde et al. 1998; Baras and Jobling 2002; Kestemont et al. 2003; Zakęś 2012). Rearing temperature should fall within the thermal comfort zone, which is specific for a given species and population. Fish raised at different temperatures generally have different growth-feed ratios, thereby the comparisons of the survival rates between fish of identical ages is not informative, because they will be at a different level of ontogenetic development (Baras et al. 2011a). However, due to the interval nature of this parameter, it is possible to rear fish closer to a lower or higher end of the comfort zone and, in this way, manipulate the metabolism rate.
Rearing at lower temperatures can exert a beneficial impact on retarding cannibalism and is connected with a stunted larvae growth rate (Kestemont et al. 2003). Pikeperch larvae behave similarly (Zakęś 2012), although in burbot (Lota lota) the impact of higher temperature on an increase in cannibalistic incidents has only been reported for larval and juvenile developmental stages and the relationship with temperature has not been examined (Barron et al. 2012). Pike larvae behave the opposite; the lowest losses due to cannibalistic behaviour have been observed at the highest temperatures investigated (28 °C) (Szczepkowski 2009). However, the influence of ontogenetic differences cannot be excluded in the cases mentioned here.
In facilities with the possibility of water temperature regulation, i.e. recirculating aquaculture systems, in order to optimize the outcome of rearing under periodic feeding conditions, the following scheme may be applied: after the last daily feeding, the water temperature may be slowly decreased (safe gradient up to 5 °C) (Baras and Jobling 2002). Such an approach should ensure optimal growth and reduce the severity of cannibalistic behaviour. Unfortunately, this method cannot be applied in species which demand constant feeding 24 h a day or in many outdoor rearing systems. However, such temperature fluctuations exist naturally in these type of rearing systems (lower temperatures at night) but without the possibility of regulation (Azaza et al. 2010).
Proper light conditions and water clarity
Light intensity, length of lighting period and water clarity impact the regulation of circadian rhythms and, therefore, influence the prevalence of cannibalism. The spectral bandwidth of light available for vision is narrower under water than on land due to the spectrally selective absorption of natural water. The colour of natural water is highly variable (Kusmic and Gualtieri 2000). In general, freshwater bodies are subject to greater variations in light than marine waters owing to the higher concentrations of suspended organic and mineral matter and organic compounds in a solution, causing a shift in the peak transmission towards the red wavelength in varying amounts, within the range of 480– 590 nm (Knowles and Dartnall 1977).
Larvae of many fish species, which rely mainly on sight in food seeking (Baras and Jobling 2002), demonstrate positive phototaxy and are short-sighted (they see at a distance approximately 1/3 of their body length), which influences feed seeking, since to ingest food, they have to see it. This is favoured by clear lighting, the colour of tank walls contrasting with feed colour and a longer lighting period, which prolongs the feed-seeking period and shortens nocturnal fasting period (which prompts cannibalism) (Baras et al. 2000a; Tamazouzt et al. 2000; Kestemont et al. 2003; Qin et al. 2004). In these species, a prolonged period of darkness is associated with reduced activity and staying near the bottom of a tank, which leads to locally increased stock density and more severe aggressive behaviour (Baras et al. 2000a). Therefore, for such species as pike (Szczepkowski 2009), pikeperch (Król and Zakęś 2016) or European perch (Tamazouzt et al. 2000; Król and Zieliński 2015), the best rearing outcomes are obtained when fish are kept under a prolonged lighting period and at a proper light intensity for a given species.
Nevertheless, long periods of good visibility translate into long periods for contact between a potential cannibal and its prey, which may contribute to an increased frequency of cannibalistic behaviour and higher heterogeneity in a stock, representing a positive feedback mechanism (Baras and Jobling 2002). Moreover, excessive light intensity may be a source of stress and intensify aggressive behaviour, as reported in Nile tilapia and Barramundi perch (Baras et al. 2000a; Jesu Arockiaraj and Appelbaum 2011). In these species, reduced light intensity and water clarity have a beneficial effect on survival and performance.
Species that prefer darkness (such as African catfish—which finds food mainly with chemoreceptors and sensor organs) react to excessively intense light with intensified behaviour associated with chasing resting individuals, searching for refuges, gathering at the bottom of a tank and increased territorial aggression associated with locally increased stock density (Britz and Pienaar 1992; Baras and Jobling 2002). In addition, chemical substances are released from wounds (injuries incurred when attacked) into the environment which are a strong stimulus triggering cannibalistic behaviour (Mukai et al. 2013). Hence, turbid water, limiting the inflow of light and prolonging the darkness period for a darkness-preferring species, prompts a reduction in aggression and cannibalism as well as prolonged feeding and more homogenous weight gains (Solomon and Udoji 2011).
In farming facilities with the possibility of light wavelength regulation, a spectrum with added light intensity could be used to gain better rearing results. Sallehudin and Mukai (2014) demonstrated that the juveniles of African catfish had low biting activity under light intensities of 0.0017 and 0.021 µmol/m2/s for white, blue, and yellow light wavelengths, and 0.19 and 1.40 µmol/m2/s for blue and red light wavelengths. Only the blue light wavelength showed low biting activity for all levels of light intensity. Similar results were obtained in beluga (Banan et al. 2011), where blue light (by establishing comfort) resulted in increased growth performance and had a positive effect on physiological conditions. Contrary to this, red light had a negative impact on beluga growth performance.
Providing refuges for weaker and poorly developing individuals
It is important to design refuges for smaller and weaker individuals that may become potential prey for cannibals. The efficacy of refuges in preventing cannibalism depends to a certain extent on lighting conditions. This aspect has been studied in barramundi perch (Qin et al. 2004) and African catfish (Hecht and Appelbaum 1988; Pienaar 1990). In these species, a significant reduction in cannibalistic behaviour and territorial aggression has been demonstrated to occur after introduction of refuges (Solomon and Udoji 2011). The amount of aggressive behaviour associated with chasing and biting was also reduced.
The size, design and number of refuges should be adapted to the biology of a given species. With too large refuges (letting in both small and large individuals), the opposite effect, i.e. an increased level of cannibalism, may occur (Qin et al. 2004). Incorrect design will not bring the expected effects if hiding requires swimming at depths unnatural for a given species (Francis and Bengtson 1999). In addition, the number of refuges should be sufficiently high to satisfy the needs of all individuals; if the number of refuges is too low, territorial aggression and fighting for refuges may rise (Britz and Pienaar 1992). Moreover, special care should be taken for darkness-preferring species such as European chub. If an opaque, dark hiding place is designed and too intense light is applied, fish could gather in such an opaque hiding place due to their marked preference for darkened locations. With incorrect lighting, fish would be less eager to leave refuges for feeding and ultimately they could die of starvation or oxygen deficiency.
Major limitations in the implementation of this method are the constant need for modification of refuge dimensions according to the growth ratio, especially during periods of quick changes in development, as well as the fact that refuges hinder many routine operations and can cause sanitary problems. For these reasons many consider using refuges laborious and inconvenient for culture.
Colour and shape of tanks
Light perception ability varies in fish due to morphological differences in eye anatomy (Kusmic and Gualtieri 2000). Colour of inside walls and shape of tanks should be adjusted for each fish species, in regards to the spectrum of used light and water turbidity as these factors influence visibility and, thus, feed ingestion. This is of particular importance for species that rely on sight in feed seeking (Sogard and Olla 1994; Baras and Jobling 2002; Üstündağ and Rad 2015). Depending on the fish species, light intensity or developmental stage, the same tank colour may exert a different impact (Tamazouzt et al. 2000). For example, for rainbow trout especially under low light intensities, beige coloured tanks are more suitable for rearing (Üstündağ and Rad 2015). Imanpoor and Abdollahi (2011) demonstrated that colour can have large impact on feeding rate, hematocrit, final body lipid content and all body colour parameters in juvenile Caspian kutum (Rutilus kutum). Survival was also influenced by the tank wall colour with light grey showing the best result (17%). In beluga (Huso huso), most of growth parameters suggested a positive impact of rearing in black tanks (Banan et al. 2011). In Eurasian perch, the greatest growth in weight and length was observed in tanks with light grey and white walls, which were strongly illuminated, while the lowest growth was recorded in a tank with black walls and 250 lx illumination (Tamazouzt et al. 2000). Moreover, in Eurasian perch, light walls seems to induce a strong contrast between brown-yellow food particles and the tank walls and, consequently, improve food perception as observed in some other fish species, such as seabream Sparus auratus (Ounais-Guschemann 1989) and walleye (Bristow and Summerfelt 1994).
The shape and size of a tank influences the movement of water, as it circulates differently in tanks with a round, square or rectangular bottom (Rożyński and Zakęś 2015). Depending on the shape and size of a tank, fish will have different access to its edges that may serve as refuges; smaller tanks have more of them (Baras and Jobling 2002). On the other hand, cannibals may more easily spot prey in a small tank (Sogard and Olla 1994). Large tanks allow establishment of more complex relations between individuals. In addition, tanks may differ in depth, and depending on the design the smaller the depth, the faster fish may reach the water surface to ingest feed or breathe in air. This is important in fish species that rely on sight, as the time needed to select a potential prey shortens as the prey swims closer to the water surface, which reduces the chances for cannibalistic behaviour (Baras and Jobling 2002). These factors influence the movement of fish in tanks and indirectly the weight gains and occurrence of cannibalism. In pike rearing, the lowest incidence of cannibalistic behaviour was found in tanks with a square bottom (Szczepkowski 2009) whereas for pikeperch, tanks with square bottoms were more efficient for older stages and larvae had better survival parameters in tanks with round bottoms, although in this experiment the impact of different lagoon depths cannot be excluded (Kozłowski et al. 2009).
Degree of domestication
The impact of domestication on the physiology and behaviour of fish is a new but widely discussed topic in the scientific world. However, further research is warranted. Only two fish species have been completely domesticated: carp (Cyprinus carpio carpio) and goldfish (Carassius auratus auratus) (Balon 2004). Other species still have reproductive cells acquired from wild populations, which may influence both their health and behaviour. With progressive domestication, susceptibility to stress decreases (Douxfils et al. 2011) and wild or partially domesticated fish demonstrate a higher tendency for aggressive behaviour than fish with a greater degree of domestication (Hedenskog et al. 2002). On the other hand, cannibalistic strains may arise during domestication, because the characteristics deemed by aquaculturists such as fast growth, may exacerbate cannibalism through increased size heterogeneity (Hecht and Pienaar 1993; Baras and Jobling 2002).
Health of stock and use of pharmaceuticals
The health condition of fish stocks is important regarding the possibility of the occurrence of cannibalism. Health problems and diseases can impair the growth of ill individuals, and increase the potential for cannibalism. In such cases, it is recommended to use therapeutics. One can choose between antibiotics for direct treatment or disinfectants as preventives. For food safety, the use of antibiotics should be limited as much as possible and they should not be overused. Subagja et al. (1999) demonstrated that use of disinfectants such as chloramine T and formalin in Asian catfish can significantly increase the survival rate and health status of fish stock.
Another approach to pharmaceutical treatment is the use of sedatives to reduce the level of stress and, thereby, cannibalism. However, this would only be treating the symptoms and not the root cause of the stress. Moreover, such an approach may be economically unjustified and may raise doubts as to the quality and safety of animal origin food products, as it would require determining the residue level standards (Gomułka and Antychowicz 1999; Neiffer and Stamper 2009).
Summary
Cannibalism represents a problem under intensive commercial production of different fish species and is impossible to manage without understanding the factors impacting its occurrence and degree of severity. It is thus necessary to constantly expand the knowledge of biology and natural habitats of reared species and species which have been newly introduced to aquaculture. This is becoming even more important as aquaculture is continuously developing and an increasing number of species are being adapted to intensive rearing and breeding. The key is to include consideration of the phenomenon of cannibalism in selection and breeding programmes.
Each case of cannibalism should be considered in the context of biology and specificity of a given species. There is not a single, universal preventive measure that would be equally effective in all species and for all types of cannibalism. In addition, a reduction in cannibalistic behaviour is not necessarily correlated with the best outcomes in production and, hence, the parameters should be carefully selected. Furthermore, there may need to be decisions made between fish survival and the final fish biomass.
Change history
16 June 2022
A Correction to this paper has been published: https://doi.org/10.1007/s11160-022-09717-8
References
Akbary P, Hosseini SA, Imanpoor M et al (2010) Comparison between live food and artificial diet on survival rate, growth and body chemical composition of Oncorhynchus mykiss larvae. Iran J Fish Sci 9:19–32
Altaff K, Janakiraman A (2013) Impact of different feeds on cannibalism in early larval stages of Koi carp (Cyprinus carpio). J Appl Biol Biotechnol 1:35–39
Amundsen P-A, Damsgård B, Arnesen AM et al (1995) Experimental evidence of cannibalism and prey specialization in Arctic charr, Salvelinus alpinus. Environ Biol Fishes 43:285–293. doi:10.1007/BF00005860
Amundsen PA, Svenning MA, Slikavuoplo SI (1999) An experimental comparison of cannibalistic response in different Arctic charr (Salvelinus alpinus (L.)) stocks. Ecol Freshw Fish 8:43–48. doi:10.1111/j.1600-0633.1999.tb00051.x
Azaza MS, Legendre M, Kraiem MM, Baras É (2010) Size-dependent effects of daily thermal fluctuations on the growth and size heterogeneity of Nile tilapia Oreochromis niloticus. J Fish Biol 76:669–683. doi:10.1111/j.1095-8649.2009.02524.x
Baardvik BM, Jobling M (1990) Effect of size-sorting on biomass gain and individual growth rates in Arctic charr, Salvelinus alpinus L. Aquaculture 90:11–16. doi:10.1016/0044-8486(90)90278-U
Babiak I, Mandiki SNM, Ratsinjomanana K, Kestemont P (2004) Initial weight and its variation in post-larval Eurasian perch affect quantitative characteristics of juvenile cohorts under controlled conditions. Aquaculture 234:263–276. doi:10.1016/j.aquaculture.2003.12.010
Balon EK (2004) About the oldest domesticates among fishes. J Fish Biol 65:1–27. doi:10.1111/j.1095-8649.2004.00563.x
Banan A, Kalbassi MR, Bahmani M, Sadati MAY (2011) Effects of colored light and tank color on growth indices and some physiological parameters of juvenile beluga (Huso huso). J Appl Ichthyol 27:565–570. doi:10.1111/j.1439-0426.2011.01682.x
Baras E (1999a) Functional implications of early sexual growth dimorphism in vundu. J Fish Biol 54:119–124. doi:10.1111/j.1095-8649.1999.tb00616.x
Baras É (1999b) Sibling cannibalism among juvenile vundu under controlled conditions. I. Cannibalistic behaviour, prey selection and prey size selectivity. J Fish Biol 54:82–105. doi:10.1006/jfbi.1998.0847
Baras É, Dalmeida AF (2001) Size heterogeneity prevails over kinship in shaping cannibalism among larvae of sharptooth catfish Clarias gariepinus. Aquat Living Resour 14:251–256. doi:10.1016/S0990-7440(01)01118-4
Baras E, Jobling M (2002) Dynamics of intracohort cannibalism in cultured fish. Aquac Res 33:461–479. doi:10.1046/j.1365-2109.2002.00732.x
Baras E, Maxi MYJ, Ndao M, Mélard C (2000a) Sibling cannibalism in dorada under experimental conditions. II. Effect of initial size heterogeneity, diet and light regime on early cannibalism. J Fish Biol 57:1021–1036. doi:10.1006/jfbi.2000.1365
Baras E, Ndao M, Maxi MYJ et al (2000b) Sibling cannibalism in dorada under experimental conditions. I. Ontogeny, dynamics, bioenergetics of cannibalism and prey size selectivity. J Fish Biol 57:1001–1020. doi:10.1006/jfbi.2000.1366
Baras E, Kestemont P, Mélard C (2003) Effect of stocking density on the dynamics of cannibalism in sibling larvae of Perca fluviatilis under controlled conditions. Aquaculture 219:241–255. doi:10.1016/S0044-8486(02)00349-6
Baras É, Hafsaridewi R, Slembrouck J et al (2010) Why is cannibalism so rare among cultured larvae and juveniles of Pangasius djambal? Morphological, behavioural and energetic answers. Aquaculture 305:42–51. doi:10.1016/j.aquaculture.2010.04.004
Baras É, Raynaud T, Slembrouck J et al (2011a) Interactions between temperature and size on the growth, size heterogeneity, mortality and cannibalism in cultured larvae and juveniles of the Asian catfish, Pangasianodon hypophthalmus (Sauvage). Aquac Res 42:260–276. doi:10.1111/j.1365-2109.2010.02619.x
Baras É, Silva del Aguila DV, Montalvan Naranjos GV et al (2011b) How many meals a day to minimize cannibalism when rearing larvae of the Amazonian catfish Pseudoplatystoma punctifer? The cannibal’s point of view. Aquat Living Resour 24:379–390. doi:10.1051/alr/2011141
Baras É, Hafsaridewi R, Slembrouck J et al (2013) Do cannibalistic fish possess an intrinsic higher growth capacity than others? A case study in the Asian redtail catfish Hemibagrus nemurus (Valenciennes, 1840). Aquac Res 45:68–79. doi:10.1111/j.1365-2109.2012.03205.x
Baras É, Dugué R, Legendre M (2014) Do cannibalistic fish forage optimally? An experimental study of prey size preference, bioenergetics of cannibalism and their ontogenetic variations in the African catfish Heterobranchus longifilis. Aquat Living Resour 27:51–62. doi:10.1051/alr/2014006
Barron JM, Jensen NR, Anders PJ et al (2012) Effects of temperature on the intensive culture performance of larval and juvenile North American Burbot (Lota lota maculosa). Aquaculture 364:67–73
Baskerville-Bridges B, Kling LJ (2000) Larval culture of Atlantic cod (Gadus morhua) at high stocking densities. Aquaculture 181:61–69. doi:10.1016/S0044-8486(99)00220-3
Bengtson DA, Léger P, Sorgeloos P (1991) Use of Artemia as a food source for aquaculture. In: Browne RA, Sorgeloos P, Trotman CNA (eds) Artemia biology. CRC Press, Boca Raton, pp 255–285
Biu AA, Gwana AM, Bukar-Kolo YM et al (2015) Effects of sorting on cannibalism in Catfish (Clarias gariepinus) rose in concrete tanks in maiduguri, North-Eastern Nigeria. Anim Vet Sci 3:67–73. doi:10.11648/j.avs.20150302.16
Blom G, Folkvord A (1997) A snapshot of cannibalism in 0-group Atlantic cod (Gadus morhua) in a marine pond. J Appl Ichthyol 13:177–181. doi:10.1111/j.1439-0426.1997.tb00118.x
Brabrand Å (1995) Intra-cohort cannibalism among larval stages of perch (Perca fluviatilis). Ecol Freshw Fish 4:70–76. doi:10.1111/j.1600-0633.1995.tb00119.x
Bristow BT, Summerfelt RC (1994) Performance of larval walleye cultured intensively in lear and turbid water. J World Aquac Soc 25:454–464. doi:10.1111/j.1749-7345.1994.tb00230.x
Britz PJ, Pienaar AG (1992) Laboratory experiments on the effect of light and cover on the behaviour and growth of African catfish, Clarias gariepinus (Pisces: Clariidae). J Zool 227:43–62. doi:10.1111/j.1469-7998.1992.tb04343.x
Conceição LEC, Yúfera M, Makridis P et al (2010) Live feeds for early stages of fish rearing. Aquac Res 41:613–640. doi:10.1111/j.1365-2109.2009.02242.x
Cuff WR (1980) Behavioral aspects of cannibalism in larval walleye, Stizostedion vitreum. Can J Zool 58:1504–1507
de Azevedo RV, Braga LGT (2012) Use of Probiotics in aquaculture. In: Rigobelo E (ed) Probiotic in animals. InTech, pp 103–118. doi: 10.5772/50056
DeAnglis DL, Hackney PA, Webb JC (1980) A partial differential equation model of changing sizes and numbers in a cohort of juvenile fish. Environ Biol Fishes 5:261–266. doi:10.1007/BF00005361
Douxfils J, Mandiki SNM, Marotte G et al (2011) Does domestication process affect stress response in juvenile Eurasian perch Perca fluviatilis? Comp Biochem Physiol Part A Mol Integr Physiol 159:92–99. doi:10.1016/j.cbpa.2011.01.021
Ferguson MM, Noakes DLG (1983) Movers and stayers: genetic analysis of mobility and positioning in hybrids of lake charr, Salvelinus namaycush, and brook charr, S. fontinalis (Pisces, Salmonidae). Behav Genet 13:213–222. doi:10.1007/BF01065670
Fessehaye Y, Rezk M, Bovenhuis H, Komen H (2005) Size dependent cannibalism in juvenile Nile Tilapia (Oreochromis niloticus). In: Bolivar RB, Mair GC, Fitzsimmons K (eds) 6th international symposium on Tilapia in Aquaculture Philippine International Convention Center Roxas Boulevard. Manila, Philippines, pp 230–238
Folkvord A (1997) Ontogeny of cannibalism in larval and juvenile fishes with special emphasis on Atlantic cod. In: Chambers RC, Trippel EA (eds) Early life history and recruitment in fish population. Springer Netherlands, Dordrecht, pp 251–278. doi:10.1007/978-94-009-1439-1
Folkvord A, Otterå H (1993) Effects of initial size distribution, day length, and feeding frequency on growth, survival, and cannibalism in juvenile Atlantic cod (Gadus morhua L.). Aquaculture 114:243–260. doi:10.1016/0044-8486(93)90300-N
Francis AW Jr, Bengtson DA (1999) Partitioning of fish and diet selection as methods for the reduction of cannibalism in Paralichthys dentatus larviculture. J World Aquac Soc 30:302–310. doi:10.1111/j.1749-7345.1999.tb00680.x
Frisk M, Skov PV, Steffensen JF (2012) Thermal optimum for pikeperch (Sander lucioperca) and the use of ventilation frequency as a predictor of metabolic rate. Aquaculture 324–325:151–157. doi:10.1016/j.aquaculture.2011.10.024
Garrison LP, Link JS (2000) Diets of five hake species in the northeast United States continental shelf ecosystem. Mar Ecol Prog Ser 204:243–255. doi:10.3354/meps204243
Gatesoupe FJ (1999) The use of probiotics in aquaculture. Aquaculture 180:147–165. doi:10.1016/S0044-8486(99)00187-8
Gomułka P, Antychowicz J (1999) Anaesthetics used in fish. Med Weter 55:89–93
Hecht T, Appelbaum S (1988) Observations on intraspecific aggression and coeval sibling cannibalism by larval and juvenile Clarias gariepinus (Clariidae: Pisces) under controlled conditions. J Zool 214:21–44. doi:10.1111/j.1469-7998.1988.tb04984.x
Hecht T, Pienaar AG (1993) A review of cannibalism and its implications in fish larviculture. J World Aquac Soc 24:246–261. doi:10.1111/j.1749-7345.1993.tb00014.x
Hedenskog M, Petersson E, Järvi T (2002) Agonistic behavior and growth in Newly emerged brown trout (Salmo trutta L.) of sea-ranched and wild origin. Aggress Behav 28:145–153. doi:10.1002/ab.90016
Hoglund E, Bakke MJ, Overli O et al (2005) Suppression of aggressive behaviour in juvenile Atlantic cod (Gadus morhua) by l-tryptophan supplementation. Aquaculture 249:525–531. doi:10.1016/j.aquaculture.2005.04.028
Hseu JR (2002) Effects of size difference and stocking density on cannibalism rate of juvenile grouper Epinephelus coioides. Fish Sci 68:1384–1386. doi:10.1046/j.1444-2906.2002.00578.x
Hseu JR, Huang WB (2014) Application of logistic regression analysis to predict cannibalism in orange-spotted grouper Epinephelus coioides (Hamilton) fry. Aquac Res 45:868–873. doi:10.1111/are.12031
Hseu JR, Lu FI, Su HM et al (2003a) Effect of exogenous tryptophan on cannibalism, survival and growth in juvenile grouper, Epinephelus coioides. Aquaculture 218:251–263. doi:10.1016/S0044-8486(02)00503-3
Hseu JR, Chang HF, Ting YY (2003b) Morphometric prediction of cannibalism in larviculture of orange-spotted grouper, Epinephelus coioides. Aquaculture 218:203–207. doi:10.1016/S0044-8486(02)00315-0
Hseu JR, Shen PS, Bin Huang W, Hwang PP (2007) Logistic regression analysis applied to cannibalism in the giant grouper Epinephelus lanceolatus fry: short paper. Fish Sci 73:472–474. doi:10.1111/j.1444-2906.2007.01358.x
Imanpoor MR, Abdollahi M (2011) Effects of tank color on growth, stress response and skin color of Juvenile caspian kutum Rutilus frisii Kutum. Glob Vet 6:118–125
Imorou Toko I, Fiogbé ED, Kestemont P (2008) Determination of appropriate age and stocking density of vundu larvae, Heterobranchus longifilis (Valenciennes 1840), at the weaning time. Aquac Res 39:24–32. doi:10.1111/j.1365-2109.2007.01857.x
Jesu Arockiaraj A, Appelbaum S (2011) Sibling cannibalism in juvenile barramundi, Lates calcarifer (Actinopterygii: Perciformes: Centropomidae), reared under different light conditions. Acta Ichthyol Piscat 41:7–11. doi:10.3750/AIP2011.41.1.02
Jobling M, Jørgensen EH, Arnesen AM, Ringø E (1993) Feeding, growth and environmental requirements of Arctic charr: a review of aquaculture potential. Aquac Int 1:20–46. doi:10.1007/BF00692662
Kailasam M, Thirunavukkarasu AR, Chandra PK, Subburaj R (2002) Influence of size variation and feeding on cannibalism of Asian sea bass Lates calcarifer (Bloch) during hatchery rearing. Indian J Fish 49:107–113
Kaleta T (2013) Behavior of captive lower vertebrates as an indicator of animal welfare. Życie Weter 88:860–866
Kania BF, Zawadzka E, Dębski B (2012) Neurohormonal basis of aggression in fish. Med Weter 68:195–198
Katavić I, Jug-Dujaković J, Glamuzina B (1989) Cannibalism as a factor affecting the survival of intensively cultured sea bass (Dicentrarchus labrax) fingerlings. Aquaculture 77:135–143. doi:10.1016/0044-8486(89)90197-X
Kestemont P, Jourdan S, Houbart M et al (2003) Size heterogeneity, cannibalism and competition in cultured predatory fish larvae: biotic and abiotic influences. Aquaculture 227:333–356. doi:10.1016/S0044-8486(03)00513-1
Knowles A, Dartnall HJA (1977) Habitat, habit and visual pigments. In: Davson H (ed) The Eye: Photobiology of Vision, Vol 2B, 2nd edn. Academic Press, NewYork, pp 581–648
Kozłowski M, Szczepkowski M, Wunderlich K et al (2009) Results of rearing juvenile pikeperch (Sander lucioperca) in different types of tanks. Komun Rybackie 6:6–10
Król J, Zakęś Z (2016) Effect of dietary l-tryptophan on cannibalism, survival and growth in pikeperch Sander lucioperca (L.) post-larvae. Aquac Int 24:441–451. doi:10.1007/s10499-015-9936-1
Król J, Zieliński E (2015) Effects of stocking density and weaning age on cannibalism, survival and growth in European perch Perca fluviatilis larvae. Pol J Nat Sci 30:403–415
Król J, Zakęś Z, Kestemont P (2013) Emergence of cannibalism in European percid fish—size heterogeneity or natural born killers consequence? In: Larvi 2013: 6th fish & shellfish larviculture symposium, pp 231–232
Kucharczyk D, Mamcarz A, Kujawa R, Skrzypczak A (1998) Development of cannibalism in larval northern pike, Esox lucius (Esocidae). Ital J Zool 65:261–263. doi:10.1080/11250009809386829
Kusmic C, Gualtieri P (2000) Morphology and spectral sensitivities of retinal and extraretinal photoreceptors in freshwater teleosts. Micron 31:183–200. doi:10.1016/S0968-4328(99)00081-5
Lepage O, Vílchez IM, Pottinger TG, Svante W (2003) Time-course of the effect of dietary l-tryptophan on plasma cortisol levels in rainbow trout Oncorhynchus mykiss. J Exp Biol 206:3589–3599. doi:10.1242/jeb.00614
Ljubobratović U, Kucska B, Feledi T et al (2015) Effect of weaning strategies on growth and survival of Pikeperch, Sander lucioperca, larvae. Turkish J Fish Aquat Sci 15:325–331. doi:10.4194/1303-2712-v15_2_15
Loadman NL, Moodie GEE (1986) Significance of cannibalism in larval Walleye (Stizostedion vitreum). Can J Fish Aquat Sci 43:613–618
Mandiki SNM, Babiak I, Król J et al (2007) How initial predator-prey ratio affects intracohort cannibalism and growth in Eurasian perch Perca fluviatilis L. larvae and juveniles under controlled conditions. Aquaculture 268:149–155. doi:10.1016/j.aquaculture.2007.04.036
Mandiki SNM, Milla S, Wang N et al (2011) Effects of probiotic bacteria on growth parameters and immune defence in Eurasian perch Perca fluviatilis L. larvae under intensive culture conditions. Aquac Res 42:693–703. doi:10.1111/j.1365-2109.2010.02744.x
Mandiki SNM, Redivo B, Baekelandt S et al (2016) Long-term tryptophan supplementation decreased the welfare and innate immune status of pikeperch juveniles. Fish Shellfish Immunol 53:113–114. doi:10.1016/j.fsi.2016.04.090
Manley CB, Rakocinski CF, Lee PG, Blaylock RB (2015) Feeding frequency mediates aggression and cannibalism in larval hatchery-reared spotted seatrout, Cynoscion nebulosus. Aquaculture 437:155–160. doi:10.1016/j.aquaculture.2014.11.012
Mélard C, Baras E, Mary L, Kestemont P (1996) Relationships between stocking density, growth, cannibalism and survival rate in intensively cultured larvae and juveniles of perch (Perca fluviatilis). Ann Zool Fennici 33:643–651. doi:10.1016/j.infsof.2008.09.005
Mousseau T (1998) The adaptive significance of maternal effects. Trends Ecol Evol 13:403–407. doi:10.1016/S0169-5347(98)01472-4
Mukai Y, Sanudin N, Firdaus RF, Saad S (2013) Reduced cannibalistic behavior of African catfish, Clarias gariepinus, larvae under dark and dim conditions. Zoolog Sci 30:421–424. doi:10.2108/zsj.30.421
Neiffer DL, Stamper MA (2009) Fish sedation, analgesia, anesthesia, and euthanasia: considerations, methods, and types of drugs. ILAR J 50:343–360. doi:10.1093/ilar.50.4.343
Nilsson PA (1999) Foraging among cannibals and kleptoparasites: effects of prey size on pike behavior. Behav Ecol 10:557–566. doi:10.1093/beheco/10.5.557
Ounais-Guschemann N (1989) Définition d’un modèle d’élevage larvaire intensif pour la daurade Sparus auratus. Dissertation, Université d’Aix-Marseille II
Pienaar AG (1990) A study of coeval sibling cannibalism in larval and juvenile fishes and its control under culture conditions. Rhodes University, Grahams Town
Piotrowska I, Szczepkowska B, Wunderlich K, Szczepkowski M (2013) Results of the larviculture of Atlantic sturgeon (Acipenser oxyrinchus) fed different types of diets. Arch Pol Fish 21:53–61. doi:10.2478/aopf-2013-0006
Policar T, Stejskal V, Kristan J et al (2013) The effect of fish size and stocking density on the weaning success of pond-cultured pikeperch Sander lucioperca L. juveniles. Aquac Int 21:869–882. doi:10.1007/s10499-012-9563-z
Post JR, Evans DO (1989) Experimental evidence of size-dependent predation mortality in juvenile yellow perch. Can J Zool 67:521–523. doi:10.1139/z89-076
Qin JG, Mittiga L, Ottolenghi F (2004) Cannibalism reduction in juvenile barramundi Lates calcarifer by providing refuges and low light. J World Aquac Soc 35:113–118. doi:10.1111/j.1749-7345.2004.tb01067.x
Ribeiro FF (2015) Cannibalism in Barramundi Lates calcarifer: understanding functional mechanisms and implication to aquaculture. Flinders University, Bedford Park
Ribeiro FF, Qin JG (2013) Modelling size-dependent cannibalism in barramundi Lates calcarifer: cannibalistic polyphenism and its implication to aquaculture. PLoS ONE 8:1–8. doi:10.1371/journal.pone.0082488
Ribeiro FF, Qin JG (2015) Prey size selection and cannibalistic behaviour of juvenile barramundi Lates calcarifer. J Fish Biol 86:1549–1566. doi:10.1111/jfb.12658
Ribeiro FF, Qin JG (2016) Bioenergetics of cannibalism in juvenile barramundi Lates calcarifer (Bloch): exploring growth advantage of fish fed live prey and formulated diet. Aquac Res 47:2324–2333. doi:10.1111/are.12685
Rożyński M, Zakęś Z (2015) Wpływ parametrów technicznych basenów i właściwości fizycznych pasz komponowanych na efekty podchowu ryb. Komun Rybackie 3:12–44
Sallehudin F, Mukai Y (2014) Cannibalistic behaviour of African catfish juveniles, Clarias gariepinus under different light wavelengths and intensities. In: Nejadkoorki F (ed) 3rd international conference on applied life sciences (ICALS2014). ISALS Publishing, Malaysia, pp 51–55
Sloman KA, Armstrong JD (2002) Physiological effects of dominance hierarchies: laboratory artefacts or natural phenomena? J Fish Biol 61:1–23. doi:10.1111/j.1095-8649.2002.tb01733.x
Smith C, Reay P (1991) Cannibalism in teleost fish. Rev Fish Biol Fish 1:41–64. doi:10.1007/BF00042661
Sogard SM, Olla BL (1994) The potential for intracohort cannibalism in age-0 walleye pollock, Theragra chalcogramma, as determined under laboratory conditions. Environ Biol Fishes 39:183–190. doi:10.1007/BF00004936
Solomon RJ, Udoji FC (2011) Canibalism among cultured African catfishes (Heterobranchus longifillis and Clarias gariepinus). Nat Sci 9(9):1–13
Steenfeldt S, Lund I, Höglund E (2011) Is batch variability in hatching time related to size heterogeneity and cannibalism in pikeperch (Sander lucioperca)? Aquac Res 42:727–732. doi:10.1111/j.1365-2109.2010.02681.x
Strand Å, Alanärä A, Staffan F, Magnhagen C (2007) Effects of tank colour and light intensity on feed intake, growth rate and energy expenditure of juvenile Eurasian perch, Perca fluviatilis L. Aquaculture 272:312–318. doi:10.1016/j.aquaculture.2007.08.052
Subagja J, Slembrouck J, Hung LT, Legendre M (1999) Larval rearing of an Asian catfish Pangasius hypophthalmus (Siluroidei, Pangasiidae): Analysis of precocious mortality and proposition of appropriate treatments. Aquat Living Resour 12(1):37–44. doi:10.1016/S0990-7440(99)80013-8
Sunde LM, Imsland AK, Folkvord A, Stefansson SO (1998) Effects of size grading on growth and survival of Juvenile turbot at two temperatures. Aquac Int 6:19–32. doi:10.1023/A:1009265602388
Szczepkowski M (2009) Impact of selected abiotic and biotic factors on the results of rearing juvenile stages of northern pike Esox lucius L. in recirculating systems. Arch Pol Fish 48:107–147. doi:10.2478/v10086-009-0010-3
Szczepkowski M, Zakęś Z, Szczepkowska B, Piotrowska I (2011) Effect of size sorting on the survival, growth and cannibalism in pikeperch (Sander lucioperca L.) larvae during intensive culture in RAS. Czech J Anim Sci 56:483–489
Szkudlarek M, Zakȩś Z (2007) Effect of stocking density on survival and growth performance of pikeperch, Sander lucioperca (L.), larvae under controlled conditions. Aquac Int 15:67–81. doi:10.1007/s10499-006-9069-7
Tamazouzt L, Chatain B, Fontaine P (2000) Tank wall colour and light level affect growth and survival of Eurasian perch larvae (Perca fluviatilis L.). Aquaculture 182:85–90. doi:10.1016/S0044-8486(99)00244-6
Thibault RE (1974) Genetics of cannibalism in a viviparous fish and its relationship to population density. Nature 251:138–140. doi:10.1038/251138a0
Trabelsi A, Gardeur JN, Teletchea F et al (2013) Hatching time effect on the intra-spawning larval morphology and growth in Northern pike (Esox lucius L.). Aquac Res 44:657–666. doi:10.1111/j.1365-2109.2011.03070.x
Trabelsi A, Jaworski A, Kamler E et al (2016) The effect of hatching time on the bioenergetics of northern pike (Esox lucius) larvae from a single egg batch during the endogenous feeding period. Fish Physiol Biochem 42:593–606. doi:10.1007/s10695-015-0162-6
Üstündağ M, Rad F (2015) Effect of different tank colors on growth performance of Rainbow trout juvenile (Oncorhynchus mykiss Walbaum, 1792). J Agric Sci 21:144–151
Vallon M, Grom C, Kalb N et al (2016) You eat what you are: personality-dependent filial cannibalism in a fish with paternal care. Ecol Evol 6:1340–1352. doi:10.1002/ece3.1966
van Damme P, Appelbaum S, Hecht T (1989) Sibling cannibalism in Koi carp, Cyprinus carpio L., larvae and juveniles reared under controlled conditions. J Fish Biol 34:855–863. doi:10.1111/j.1095-8649.1989.tb03369.x
van den Bosch F, de Roos AM, Gabriel W (1988) Cannibalism as a life boat mechanism. J Math Biol 26:619–633. doi:10.1007/BF00276144
Wallat GK, Tiu LG, Wang HP et al (2005) The effects of size grading on production efficiency and growth performance of yellow perch in earthen ponds. N Am J Aquac 67:34–41. doi:10.1577/FA04-003.1
Winberg S, Øverli Ø, Lepage O (2001) Suppression of aggression in rainbow trout (Oncorhynchus mykiss) by dietary l-tryptophan. J Exp Biol 204:3867–3876
Wootton RJ (1989) Ecology of teleost fishes. Springer, Netherlands
Yang S, Yang K, Liu C et al (2015) To what extent is cannibalism genetically controlled in fish? A case study in juvenile hybrid catfish Silurus meridionalis-asotus and the progenitors. Aquaculture 437:208–214. doi:10.1016/j.aquaculture.2014.12.005
Zakęś Z (2012) The effect of body size and water temperature on the results of intensive rearing of pike-perch, Stizostedion lucioperca (L.) fry under controlled conditions. Arch Pol Fish 20:165–172. doi:10.2478/v10086-012-0020-4
Zakęś Z, Demska-Zakęś K (1996) Effect of diets on growth and reproductive development of juvenile pikeperch, Stizostedion lucioperca (L.), reared under intensive culture conditions. Aquac Res 27:841–845. doi:10.1046/j.1365-2109.1996.00801.x
Żarski D, Kucharczyk D, Kwiatkowski M et al (2008) The effect of stocking density on the growth and survival of larval asp, Aspius aspius (L.), and European chub, Leuciscus cephalus (L.), during rearing under controlled conditions. Arch Pol Fish 16:371–381. doi:10.2478/s10086-008-0025-1
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Duk, K., Pajdak, J., Terech-Majewska, E. et al. Intracohort cannibalism and methods for its mitigation in cultured freshwater fish. Rev Fish Biol Fisheries 27, 193–208 (2017). https://doi.org/10.1007/s11160-017-9465-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11160-017-9465-2