
Abstract
Feeding and food selection of burbot (Lota lota L.) larvae reared in illuminated cages were studied. The experiment was carried out in mesotrophic Lake Maróz, in north-eastern Poland, for 6 weeks in two successive years. The initial stocking density was 1,250 larvae (20 DPH) per cage. Food selection according to the zooplankton groups (Rotifera, Cladocera and Copepoda) and length classes was expressed by the Strauss linear selectivity index (L). Zooplankton species composition in the lake was similar in the two seasons of the study and organisms shorter than 0.5 mm prevailed in the plankton. The mean number of prey found in burbot alimentary tracts increased from about 40 up to over 200 during the course of the study. A very large inter-individual variation in the amount of food organisms consumed by fish was noted. Analysis of the values of the Strauss food selectivity index shows that at the beginning of the first year of the experiment, burbot larvae preferred copepods, most numerous in the environment at that time; later, fish tended to select cladocerans. In the second year of the study, fish more often ate copepods, irrespective of their quantities in the environment. During the whole study, reared burbot larvae did not eat rotifers, even when they were numerous in cages. Similarly to the rotifers, the smallest planktonic organisms, measuring up to 0.5 mm in length, were typically neglected by fish, while the 0.6–1.0 mm group was most frequently selected. There were also considerable individual differences between particular burbot specimens in their food preferences.
Similar content being viewed by others

Introduction
Burbot, Lota lota (L.), is the only freshwater fish species, which belongs to the order Gadiformes (Nelson 1994). Since the 20th century, due to the changes in the freshwater environment, thermal anomalies and significant overfishing, a regular decrease in the population of burbot within the entire area of its appearance has been observed (Paragamian et al. 2005, 2008; Hardy et al. 2008). In Poland, this species is included in the Red list of freshwater ichthyofauna of Poland (Witkowski et al. 1999). Currently, the fish has gained an increasing interest from the environmental and economic perspective (Kujawa et al. 2002), which has resulted in the development of controlled larval rearing procedures that will ensure production of sufficient material for stock enhancement. This applies to both reproductive biotechnology (Kujawa et al. 2002; Kucharczyk et al. 2004; Żarski et al. 2010) and the initial rearing of larvae under controlled conditions (Wolnicki et al. 2001, 2002; Kujawa et al. 2002; Shiri Harzevili et al. 2003, 2004; Żarski et al. 2009; Barron et al. 2012; Wocher et al. 2013).
The usability of formulated diets during the initial rearing is limited, and Artemia nauplii have been proven to be most suitable food for burbot larvae (Wolnicki et al. 2002; Żarski et al. 2009; Wocher et al. 2013).
The quality of stocking material determines the success of stocking. Hartmann (1983) suggests that one of the reasons why the stocking of water bodies with larvae reared under controlled conditions often fails may be that the larvae, already preconditioned in hatcheries to accept a certain size and type of food, are not flexible enough to change their strategies ad hoc in the wild. The rearing of fish larvae in illuminated cages is one of the methods of stocking material production based on zooplankton, without additional feeding. Cages are illuminated at night to attract zooplankton. References are available on the use of illuminated net cages for the rearing of larvae of the following fish: coregonid (Mamcarz 1995), Asian sea bass Lates calcarifer (Fermin et al. 1996), tench Tinca tinca (Pyka 1981), pike-perch Stizostedion lucioperca (Hilge and Steffens 1996), perch Perca fluviatilis (Skrzypczak et al. 1998), asp Aspius aspius (Mamcarz et al. 2001), bream Abramis brama (Žiliukienė 2005) and pike Esox lucius (Mamcarz et al. 1998; Žiliukienė and Žiliukas 2006). Paragamian et al. (2011) reared burbot larvae L. lota maculosa in cages, at low densities, with and without supplementary feeding. The supplementary feeding consisted of zooplankton collected from an offsite location and added to the cages. Unfortunately, data on producing the stocking material of burbot in illuminated net cages are scarce. The purpose of the present work has been to study feeding and food selection of burbot larvae reared in illuminated net cages.
Materials and methods
Broodstock and primary rearing
The experiment was conducted during two consecutive years. In both years of the experiment, burbot spawners originating from the Szczecin Lagoon (north-western Poland) were caught and transported to a hatchery of the Department of Lake and River Fisheries at the University of Warmia and Mazury in Olsztyn (Poland). Spawners were kept in 1,000 dm3 tanks (Kujawa et al. 1999). Their artificial reproduction was conducted without hormonal stimulation according to the methodology described by Żarski et al. (2010). Eggs were incubated in Weiss jars at 2 °C. Shortly before hatching, the eggs were moved to larger tanks (200 dm3), in which hatching and initial rearing took place. The temperature of water was raised to 8 °C (±0.3) during mass hatching and then to 10 °C (±0.2) for two weeks during resorption of the yolk sac by the larvae (as described by Kujawa et al. 2002).
Initial rearing in the laboratory was conducted in an open water system for 20 days at 12 °C (Kujawa et al. 2000). The inflow of water from the top and gentle aeration was provided. During the rearing, physicochemical parameters of water were monitored (dissolved oxygen, ammonia concentration, nitrates, nitrites and pH). The content of dissolved oxygen did not drop below 6 mg dm−3, while the concentration of ammonia was below 0.2 mg dm−3, nitrates—below 20 mg dm−3, nitrites—below 0.04 mg dm−3 and the pH ranged from 7.2 to 7.8. The photoperiod was 15 h (15L:9D). Burbot larvae were fed ad libitum 4 times a day using freshly hatched Artemia nauplii. After 20 days, the larvae were transferred to the illuminated net cages.
Cage rearing conditions
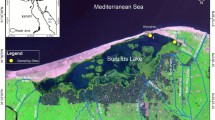
Burbot larvae were reared in illuminated cages in mesotrophic Lake Maróz (332.5 ha, max. depth 41.0 m, north-eastern Poland). Two different batches of larval fish spent six weeks in cages in two successive years. The experiment lasted for 42 days: from 30 April to 11 June in the first year and from 10 April to 22 May in the second year. The water temperature of the lake measured at a depth of 1 m, once a week on days of sample collection, varied from 10 to 15.5 °C and from 4.5 to 17 °C during the first and the second year, respectively.
Every year, two floating cages with fish and one for zooplankton were set, each measuring 1 m × 1 m and 1.5 m in depth, with a technological (rearing) capacity of 1 m3. At first, the cages were fitted with a net mesh size 1.8 mm, which—after three weeks of rearing fish larvae—was replaced with the mesh size 3.5 mm. Each cage was stocked with 1,250 burbot larvae (initially reared in controlled conditions), so the density was about 1.25 fish per dm3.
In order to attract zooplankton, the cages were illuminated at night with a bulb (60 W/24 V, 600 lx) placed 15 cm above the water surface, in the middle of a cage. The exterior of the cages was cleaned up once a week to prevent silt and algal accumulation.
Methods
For determination of the food accessibility to reared burbot larvae, samples of zooplankton were collected from an empty cage of the same size located among cages stocked with fish. Presumably, in all the cages, zooplankton was attracted by light in the same way (Okubo 1972). Samples were collected at midnight once a week (7 samples in each experimental year) by passing a vertically set plankton net (mesh size 0.2 mm) from 1.0 m depth to the water surface. At the same time, fish samples (n = 20 individuals) were collected from every cage. Immediately afterwards, the zooplankton and fish samples were preserved in 4 % formaldehyde.
Microscopic examination of zooplankton samples was performed so as to determine the species structure and count the individuals. All or at least 30 specimens from each plankton species (except Rotifera) identified in a sample were measured (±0.1 mm) using an ocular scale. In the fish samples, all individuals were measured (TL ± 1 mm) and weighed (±0.1 mg). Having dissected the alimentary tract from a fish, its content was examined under a microscope for prey composition and number. Again, all or at least 30 specimens from each plankton species (except Rotifera) found in the alimentary tract were measured (±0.1 mm). Altogether, 14 zooplankton and 480 fish samples were examined. Differences between two years of rearing in total length and body weight of fish as well as the number of ingested zooplankton organisms were analysed with the use of t test (p < 0.05). Statistical analysis was performed with the STATISTICA 9.0 (StatSoft, USA) software.
Three taxonomic groups (Rotifera, Cladocera and Copepoda) and five length classes (0.0–0.5, 0.6–1.0, 1.1–1.5, 1.6–2.0 mm and over 2 mm) were distinguished in zooplankton samples. Rotifera belonged to the smallest group. Food selection by burbot larvae with reference to the above zooplankton groups and length classes was analysed using the Strauss linear index of food selection (Strauss 1979):
where r i is the relative abundance of prey in the gut (as a percentage of the total gut contents) and pi the relative abundance of the same prey in the zooplankton sample. The value of the L index ranges from −100 to +100. Positive values imply preference and negative ones signal avoidance. For random feeding, the expected value of L is zero. t tests at 5 % significance were run to determine whether L differed significantly from zero (Strauss 1979; Sutela and Huusko 1998). If a given taxonomic or length group of zooplankton was present in the environment but was not ingested by fish, t test statistics and standard deviation (SD) values equalled zero.
Results
The composition of zooplankton species was similar in the two seasons of the study. Rotifers were dominated by Keratella quadrata, Keratella cochlearis, Syncheta sp., Asplanchna priodonta, Ascomorpha sp., Brachionus angularis. Bosmina longirostris, Bosmina coregoni and Daphnia cucullata prevailed among cladocerans. Copepods were dominated by larval and juvenile stages, i.e. nauplii and copepodites, as well as Cyclops sp., Eudiaptomus graciloides, Mesocyclops leuckarti.
In the first season of the experiment, the cage zooplankton density ranged from 93 indiv. dm−3 on day 7 to 4,380 indiv. dm−3 on day 14 of rearing (Fig. 1). In the first two weeks, Copepoda prevailed in the plankton, corresponding to 61.5 % of the total number of organisms; in the following weeks, Cladocera were more abundant (up to 68.3 %). Rotifera reached 26.6 % (Fig. 2a). During the study, organisms shorter than 0.5 mm (small cladocerans, juvenile copepods and rotifers) prevailed in the plankton, making up from 51.3 to 97.9 % of the total number of organisms (Fig. 2b). Specimens from the 0.6–1.0 mm and 1.1–1.5 mm length classes were less numerous (to 41.0 and 10.3 %, respectively). Organisms larger than 1.5 mm were scarce.

Zooplankton density in illuminated cages in Lake Maróz in the two consecutive years (1 and 2) of the experiment

Percentage composition of zooplankton (a, b, c, d) in illuminated cages in Lake Maróz in the two consecutive years (1 and 2) of the experiment
In the second season of burbot rearing, the lowest concentration of zooplankton (about 400 indiv. dm−3) occurred in April (day 1 and 14), while the highest concentration (7,292 indiv. dm−3) was observed at the end of the study (Fig. 1). Copepoda predominated in the first three weeks (59.6–90.0 % of the total number of organisms), but Cladocera were more numerous (to 83.0 %) in the final 2 weeks. Rotifera made up 59 % of the total number of organisms (Fig. 2c). In the first phase of the experiment (2 weeks), organisms of the classes 0.6–1.0 and 1.1–1.5 mm had a significant share in the size structure of zooplankton (corresponding to 61.0 and 34.0 % of the total population) (Fig. 2d). These were mainly Copepoda (copepodites, Cyclops sp., Mesocyclops leuckarti). Afterwards, the smallest organisms (less than 0.5 mm long) prevailed, constituting up to 96 % of the total number of planktonic organisms. Organisms larger than 1.5 mm occurred sporadically.
In the first season of the experiment, the cages were stocked with burbot juveniles with the mean length (TL) 22.4 mm (±3.3) and weight 101.1 mg (±44.4); in the second season, the stocked fish were 15.2 mm (±1.2) long and weighed 40.5 mg (±7.2) on average (Fig. 3). At the end of the study, the mean length of the fish was 33.1 mm (±3.5) and 26.7 mm (±3.5), while the mean weight reached 272.6 mg (±71.2) and 156.7 mg (±47.2) in the first and second year, respectively (Fig. 3). Significant differences in the total length and body weight of fish (p < 0.05) between the two years of experiment were observed on the corresponding days of the experiment. The survival rate was 77.1 % in the first and 63.9 % in the second year.

The mean weight and length of burbot Lota lota larvae reared in illuminated cages in Lake Maróz in the two following years (1 and 2) of experiment; vertical bars represent SD; different letters indicate statistical differences between groups at p < 0.05
The mean number of prey found in burbot alimentary tracts increased from about 40 individuals on the day 7 of the rearing up to over 200 during the first year of the study (Fig. 4). During the second year, the mean number of prey fluctuated from about 30 to nearly 90 specimens. From day 14 of the experiment on, there were significant differences in the number of ingested prey between the two years and the respective days of rearing. The amounts of food organisms consumed by fish in both seasons were highly varied. The alimentary tracts contained from 1 up to 618 and up to 453 zooplankton organisms in the first and second year of the experiment, respectively. There was only one fish found with an empty gut. No cannibal fish, i.e. with burbot larvae in the digestive tract, were found.

The mean number of prey in alimentary tracts of Lota lota larvae reared in two successive years (1 and 2) in illuminated cages; vertical bars represent SD; different letters indicate statistical differences between groups at p < 0.05
The analysis of the Strauss food selectivity index values shows that at the beginning of the first season of the experiment, burbot larvae preferred copepods (mainly Cyclops sp. and copepodites) (Table 1), most numerous in environment at that time (Fig. 2a). In the next two weeks (day 14), values of the L for Copepoda were still positive, although cladocerans began to predominate in the environment. From the third week of the rearing to the end of the experiment, the fish tended to select cladocerans (mainly Bosmina sp. and less often Dapnia sp.), even though their share in the zooplankton structure decreased (Table 1, Fig. 2a).
Copepods (copepodites) were often selected as food in the second rearing season (Table 1). This is clearly demonstrated in the last 2 weeks of the experiment, when the fish preferred copepods although the largest group in the environment was cladocerans (Fig. 2c). Even if the fish chose to feed on both Cladocera and Copepoda (as on days 14 and 21 of the rearing), the Strauss index was higher for copepods (Table 1). It was only at the beginning of the experiment and on day 28 of the rearing that burbot larvae preferred cladocerans (Daphnia sp. and Bosmina sp.) as food.
During the whole study, the reared burbot did not eat rotifers (Tables 1, 2), even if they were numerous in the cages (Fig. 2c). Similarly to rotifers, the smallest planktonic organisms, measuring up to 0.5 mm in length, were typically neglected by fish, regardless of their number in the environment (Table 2, Fig. 2b, d). In contrast, positive food selectivity was observed for the class of organisms of 0.6–1.0 mm during almost the entire experiment, despite their small share in the structure of zooplankton. Positive selectivity for organisms larger than 1 mm, rare in the environment, was seldom reported. Positive values of the selectivity index suggest that fish randomly ingested such large organism. Negative values, often significantly different from zero, mean that fish frequently avoided such big zooplankton.
Considerable inter-individual differences were also noticed between particular burbot juveniles in their food preferences. For example, one specimen sampled on day 21 of the second season of rearing showed positive selectivity for copepods at L = + 77.4, and its preference for cladocerans was negative (L = −8.87), whereas the respective preferences of another individual were at the levels of L = −22.6 and L = + 91.1. Similar differences were observed for the length classes of zooplankton. On the same day and in the same cage as above, the Strauss index values for the smallest organisms (to 0.5 mm) varied from L = −84.7 to L = + 15.3 and for the 0.6–1.0 mm group from L = −7.8 to L = + 83.8.
Discussion
Successful rearing of fish larvae in cages depends on environmental conditions, of which food availability is among the most important factors (Flüchter 1980; Paragamian et al. 2011). The method of rearing burbot larvae in illuminated cages was based on the positive phototaxy of crustacean plankton. The composition of the zooplankton species in Lake Maróz was similar in the two seasons of the study, with the prevailing share of organisms shorter than 0.5 mm in the plankton (Fig. 2). Considering the fact that the optimum number of zooplankton is estimated at 200–260 indiv. dm−3 (Dąbrowski 1976), the plankton concentration in the cages during the study, usually exceeding 400 indiv. dm−3 (Fig. 1), should have been sufficient for burbot larvae. This is confirmed by the absence of starving larvae as well as fish cannibals. At plankton densities of about 100 indiv. dm−3, any additional amounts of zooplankton improved the survival rate of larval burbot (Paragamian et al. 2011).
Fluctuations in the zooplankton abundance and composition affect food selection by larval burbot. The use of planktonic organisms by fish larvae is determined by both species composition and their size structure (Huusko et al. 1988).
Burbot larvae reared in the present study were in the second ontogenetic stage of feeding according to Hartmann (1983), when the size rather than number of eaten particles increases and the quality of food changes with the fish length. The mean number of prey ingested by the reared burbot larvae increased from about 40 items to even over 200 during the experiment (Fig. 4). This increase was greater in the first year of the study, which may be related to the larger size of fish (Fig. 3). A very large variation in the amount of food organisms consumed by fish was noted (Fig. 4, bars of the standard deviation). Similar results were obtained by Žiliukienė (2005) for bream Abramis brama and Žiliukienė and Žiliukas (2006) for pike Esox lucius larvae. Such big inter-specimen variation in the amount of ingested food reported in the cited studies is indicated by the range of indices of the alimentary tract filling, e.g. 67.1–576.9 ‰, at the average length of bream larvae 12.5 mm (Žiliukienė 2005). Differences in the amounts of ingested prey could have resulted from differences between individual fish. Reared burbot larvae were different in size (Fig. 3, bars of the standard deviation). The success of feeding depends on several factors, including the size of a larva and its previous feeding experience. Houde and Schekter (1980) demonstrated the effect of an increase in body weight of sea bream Archosargus rhomboidalis on its ability to catch prey. The number of successful catches rose from 53 to 76 % when dry matter of larvae increased from 10 to 200 μg. According to Braum (1967), the success rate was the same (3.1 %) for larvae of Coregonus wartmanii at both 1–8 days and 9–16 days of rearing if larvae were fed for the first time. In contrast, it increased from 3.2 to 21.6 % if larvae had previous experience of being fed before observations.
Ghan and Sprules (1993) found that the number of prey in burbot larvae stomachs and the prey size most often selected by burbot increased as they grew. The researchers concluded that the prey width rather than length limited the size of prey ingested. The values of the Strauss food selectivity index indicate that at the beginning of the first year of the experiment, burbot larvae selected copepods, most numerous in the environment at that time; afterwards (from the third week of rearing), the fish tended to prefer cladocerans (Table 1). In the second year of the study, fish more often ate copepods, irrespective of their quantities in the environment. Rotifers and the smallest planktonic organisms, measuring up to 0.5 mm in length, were usually neglected by fish, even when they were numerous in cages (Tables 1, 2). Prey from the 0.6–1.0 mm group was most frequently selected. There were also considerable individual differences between particular burbot specimens in their food preferences. References demonstrate similar tendencies for other fish species (e.g. peled, Coregonus peled and bream) reared in illuminated cages (Furgała-Selezniow et al. 2005; Žiliukienė 2005).
Studies of Hartmann (1983) showed that burbot was the only fish of the seven species examined in Lake Constance, which started to eat increasingly larger particles from the onset of foraging. Therefore, it can be assumed that the mouth size and force of suction, i.e. the parameters which are related to the fish size and which restrict possible dimensions of prey organisms, especially in the first days of fish life (Hartmann 1986; Ponton and Müller 1990), were not the limiting parameters in the present experiment. Our results are congruent with the optimal foraging theory (OFT) and the pattern of larvae (Gerking 1994), which state that in the long run, larvae ingest the most abundant and intermediate-size zooplankton rather than select the biggest planktonic organism available, even if they are physically able to ingest it. Similar results were obtained by Huusko et al. 1988 and Ponton and Müller 1990. Gerking (1994) suggests that larvae switch from a reward system based on the time of swallowing to the speed of digestion and this corresponds to the OFT.
The conducted experiments have shown that burbot larvae could be reared in illuminated cages without additional feeding at densities over 1,000 fish m−3, and the size of prey was not a limiting factor for burbot more than 15 mm in length (TL). The larvae tended to select medium-size prey, irrespective of their number in the environment. In order to develop a mass-rearing technology of burbot larvae in illuminated cages, further studies on rearing in different densities, on the availability of food and on the food selectivity by fish of different sizes are needed.
It is necessary to compare the quality of stocking material of burbot reared under controlled conditions and fed with Artemia nauplii and formulated diets with burbot reared in illuminated cages with zooplankton as the only food.
References
Barron JM, Jensen NR, Anders PJ, Egan JP, Ireland SC, Cain KD (2012) Effects of temperature on the intensive culture performance of larval and juvenile North American burbot (Lota lota maculosa). Aquaculture 364–365:67–73
Braum E (1967) The survival of fish larvae in reference of their feeding behaviour and the food supply. In: Gerking SD (ed) Biological basis of freshwater fish production. Oxford, Blackwell Sci Publ, pp 113–131
Dąbrowski K (1976) How to calculate the optimal density of food for fish larvae. Env Biol Fish 1:87–89
Fermin AC, Bolivar MEC, Gaitan A (1996) Nursery rearing of the Asian sea bass, Lates calcarifer, fry in illuminated floating net cages with different feeding regimes and stocking densities. Aquac Living Resour 9:43–49
Flüchter J (1980) Review of the present knowledge of rearing whitefish (Coregonidae) larvae. Aquaculture 19:191–208
Furgała-Selezniow G, Mamcarz A, Skrzypczak A (2005) Food selection of peled larvae (Coregonus peled Gmel.), reared in illuminated cages in different water bodies. Electr J Pol Agric Univ, Fisheries 8(2). Available Online http://www.ejpau.media.pl/volume8/issue2/art34.html. Cited 7 Feb 2013
Gerking SD (1994) Feeding ecology of fish. Academic Press, A Division of Harcourt Brace & Company, San Diego
Ghan D, Sprules WG (1993) Diet, prey selection, and growth of larval and juvenile burbot Lota lota (L.). J Fish Biol 42:47–64
Hardy R, Paragamian VL, Neufeld MD (2008) Zooplankton communities and burbot relative abundance of some oligotrophic lakes of Idaho, USA and British Columbia, Canada. Am Fish Soc Symp 59:79–89
Hartmann J (1983) Two feeding strategies of young fishes. Arch Hydrobiol 96:496–509
Hartmann J (1986) Interspecific predictors of selected prey of young fishes. Arch Hydrobiol Beih Ergebn Limnol 22:373–386
Hilge V, Steffens W (1996) Aquaculture of fry and fingerling of pike-perch (Stizostedion lucioperca L.)—a short review. J Appl Ichthyol 12:167–170
Houde ED, Schekter RC (1980) Feeding by marine fish larvae: developmental and functional responses. Environ Biol Fishes 5:315–334
Huusko A, Sutela T, Karjalainen J, Auvinen H, Alasaarela E (1988) Feeding of vendace (Coregonus albula L.) fry in a natural-state lake and a regulated lake in Northern Finland. Finnish Fish Res 9:447–456
Kucharczyk D, Mamcarz A, Kujawa R, Babiak I, Glogowski J, Sarosiek B, Kowalski R, Skrzypczak A, Targonska-Dietrich K, Chwaluczyk R (2004) Burbot (Lota lota L.) reproduction in captivity: comparison of different methods. In: Yannick J (ed) Elevage de la lotte (Lota lota). Recherche et perspectives. FLAC, Nancy, pp 12–16
Kujawa R, Kucharczyk D, Mamcarz A (1999) A model system for keeping spawners of wild and domestic fish before artificial spawning. Aquac Eng 20:85–89
Kujawa R, Mamcarz A, Kucharczyk D, Skrzypczak A (2000) An experimental unit for rearing of larval freshwater fish. Folia Univ Agric Stetin Piscatoria 26:103–108
Kujawa R, Kucharczyk D, Mamcarz A (2002) Miętus. Wyd IRS
Mamcarz A (1995) Rearing of coregonid (Coregonus sp.) larvae in illuminated cages: a review. Arch Hydrobiol Spec Issues Adv Limnol 46:287–292
Mamcarz A, Kucharczyk D, Kujawa R, Skrzypczak A, Furgala-Selezniow G (1998) Ontogeny of feeding habits in northern pike, Esox lucius (Esocidae), larvae reared in illuminated cages. Ital J Zool 65:251–253
Mamcarz A, Kujawa R, Furgala-Selezniow G, Kucharczyk D (2001) Preliminary results of larval and fry asp (Aspius aspius L.) rearing in the illuminated cages. In: Hendry CI, Van Stappen G, Wille M, Sorgeloos P (eds) Larvi 2001—Fish and shellfish larviculture symposium, vol 30. European Aquaculture Society, Spec Publ, Oostende Belgium, pp 328–331
Nelson JS (1994) Fishes of the world. Wiley, New York, pp 227–238
Okubo A (1972) A note on small organisms diffusion around an attractive center: a mathematical model. J Oceanogr Soc Jpn 28:1–7
Paragamian VL, Hardy R, Gunderman B (2005) Effects of regulated discharge on burbot migration. J Fish Biol 66:1199–1213
Paragamian VL, Pyper BJ, Daigneault J, Beamesderfer RCP, Ireland SC (2008) Population dynamics and extinction risk of burbot in the Kootenai River, Idaho, USA and British Columbia, Canada. Am Fish Soc Symp 59:213–234
Paragamian VL, Laude C, Cain KD, Barron JM, Jensen NR (2011) A novel experiment of rearing burbot larvae in cages. J Appl Ichth 27(1):16–21
Ponton D, Müller R (1990) Size of prey ingested by whitefish, Coregonus sp., larvae. Are Coregonus larvae gape-limited predators? J Fish Biol 36:67–72
Pyka J (1981) Próba podchowu młodocianych stadiow lina w oświetlanych sadzach jeziorowych. Gosp Ryb 1:12–13
Shiri Harzevili A, De Charleroy D, Auwerx J, Vught I, Van Slycken J, Dhert P, Sorgeloos P (2003) Larval rearing of burbot (Lota lota L.) using Brachionus calyciflorus rotifer as starter food. J Appl Ichth 19:84–87
Shiri Harzevili A, Dooremont I, Vught I, Auwerx J, Quataert P, De Charleroy D (2004) First feeding of burbot, Lota lota (Gadidae, Teleostei) larvae under different temperature and light conditions. Aquac Res 35:49–55
Skrzypczak A, Mamcarz A, Kujawa R, Kucharczyk D, Furgala-Selezniow G (1998) Feeding habits of larval perch, Perca fluviatilis (Percidae). Ital J Zool 65:243–245
Strauss RE (1979) Reliability estimates for Ivlev’s electivity index, the forage ratio, and a proposed linear index of food selection. Trans Am Fish Soc 108:344–352
Sutela T, Huusko A (1998) Prey selection and the density of suitable food for vendace (Coregonus albula) larvae in Lake Lentua. Arch Hydrobiol Spec Issues Adv Limnol 50:39–48
Witkowski A, Błachuta J, Kotusz J, Heese T (1999) Czerwona lista słodkowodnej ichtiofauny Polski. Chrońmy Przyr Ojczystą 55(4):5–19
Wocher H, Harsa’nyi A, Schwarz FJ (2013) Larviculture of burbot (Lota lota L.) using Artemia and weaning onto dry feed. Aquac Res 44:106–113
Wolnicki J, Myszkowski L, Kamiński R (2001) The influence of water temperature on the growth, survival, condition and biological quality of juvenile burbot, Lota lota (L.). Arch Pol Fish 9(1):79–86
Wolnicki J, Kamiński R, Myszkowski L (2002) Temperature-influenced growth and survival of burbot Lota lota (L.) larvae fed live food under controlled conditions. Arch Pol Fish 10(1):109–113
Żarski D, Sasinowski W, Kucharczyk D, Kwiatkowski M, Krejszeff S, Targońska K (2009) Mass initial rearing of burbot Lota lota (L.) larvae under controlled conditions. Pol J Nat Sci 24(1):76–84
Żarski D, Kucharczyk D, Sasinowski W, Targońska K, Mamcarz A (2010) The influence of temperature on successful reproductions of burbot, Lota lota L. under hatchery conditions. Pol J Nat Sci 25(1):93–105
Žiliukienė V (2005) The diet of Abramis brama (L.) larvae reared in illuminated cages. J Appl Ichthyol 21:406–409
Žiliukienė V, Žiliukas V (2006) Feeding of early larval pike Esox lucius reared in illuminated cages. Aquaculture 258:378–387
Acknowledgments
We are grateful to Prof. M. Jankun from the Department of Ichthyology, University of Warmia and Mazury for valuable comments and inspiring suggestions.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution License which permits any use, distribution, and reproduction in any medium, provided the original author(s) and the source are credited.
About this article
Cite this article
Furgała-Selezniow, G., Skrzypczak, A., Kucharczyk, D. et al. Food selection of burbot (Lota lota L.) larvae reared in illuminated net cages in mesotrophic Lake Maróz (north-eastern Poland). Aquacult Int 22, 41–52 (2014). https://doi.org/10.1007/s10499-013-9707-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10499-013-9707-9