
Abstract
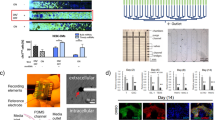
Maintaining cell viability is a major challenge associated with transplanting cells into ischemic myocardium to restore function. A likely contributor to significant cell death during cardiac cell therapy is hypoxia/anoxia. We developed a system that enabled quantification and association of cell survival with oxygen and nutrient values within in vitro constructs. Myoblasts were suspended in 2% collagen gels in 1 cm diameter × 1 cm deep constructs. At 48 ± 3 h post-seeding, oxygen levels were measured using microelectrodes and gels were snap-frozen. Bioluminescence metabolite imaging and TUNEL staining were performed on cryosections. Oxygen and glucose consumption and lactate production rates were calculated by fitting data to Fick’s second law of diffusion with Michaelis–Menten kinetics. Oxygen levels dropped to 0 mmHg and glucose levels dropped from 4.28 to 3.18 mM within the first 2000 μm of construct depth. Cell viability dropped to approximately 40% over that same distance and continued to drop further into the construct. We believe this system provides a reproducible and controllable test bed to compare survival, proliferation, and phenotype of various cell inputs (e.g., myoblasts, mesenchymal stem cells, and cardiac stem cells) and the impact of different treatment regimens on the likelihood of survival of transplanted cells.








Similar content being viewed by others

References
Beauchamp J. R., Pagel C. N., Partridge T. A. 1997 A dual-marker system for quantitative studies of myoblast transplantation in the mouse. Transplantation 63(12):1794–1797
Braun R. D., Lanzen J. L., Snyder S. A., Dewhirst M. W. 2001 Comparison of tumor and normal tissue oxygen tension measurements using OxyLite or microelectrodes in rodents. Am. J. Physiol. Heart Circ. Physiol. 280(6):H2533–2544
Brosemer R. W., Rutter W. J. 1961 The effect of oxygen tension on the growth and metabolism of a mammalian cell. Exp. Cell Res. 25:101–113
Bruick R. K. 2003 Oxygen sensing in the hypoxic response pathway: regulation of the hypoxia-inducible transcription factor. Genes Dev. 17(21):2614–2623
Carmeliet P., Dor Y., Herbert J. M., Fukumura D., Brusselmans K., Dewerchin M., Neeman M., Bono F., Abramovitch R., Maxwell P., Koch C. J., Ratcliffe P., Moons L., Jain R. K., Collen D., Keshert E. 1998 Role of HIF-1alpha in hypoxia-mediated apoptosis, cell proliferation and tumour angiogenesis. Nature 394(6692):485–490
Carrier R. L., Rupnick M., Langer R., Schoen F. J., Freed L. E., Vunjak-Novakovic G. 2002 Perfusion improves tissue architecture of engineered cardiac muscle. Tissue Eng. 8(2):175–188
Casey T. M., Arthur P. G. 2000 Hibernation in noncontracting mammalian cardiomyocytes. Circulation 102(25):3124–3129
Casey T. M., Pakay J. L., Guppy M., Arthur P. G. 2002 Hypoxia causes downregulation of protein and RNA synthesis in noncontracting Mammalian cardiomyocytes. Circ. Res. 90(7):777–783
Colton C. K. 1995 Implantable biohybrid artificial organs. Cell Transplant. 4(4):415–436
Dales S. 1960 Effects of anaerobosis on the rates of multiplication of mammalian cells cultured in vitro. Can. J. Biochem. Phisiol. 38:871–878
Debska G., Kicinska A., Skalska J., Szewczyk A., May R., Elger C. E., Kunz W. S. 2002 Opening of potassium channels modulates mitochondrial function in rat skeletal muscle. Biochim. Biophys. Acta 1556(2–3):97–105
Dib N., McCarthy P., Campbell A., Yeager M., Pagani F. D., Wright S., MacLellan W. R., Fonarow G., Eisen H. J., Michler R. E., Binkley P., Buchele D., Korn R., Ghazoul M., Dinsmore J., Opie S. R., Diethrich E. 2005 Feasibility and safety of autologous myoblast transplantation in patients with ischemic cardiomyopathy. Cell Transplant. 14(1):11–19
Froese G. 1962 The respiration of ascites tumour cells at low oxygen concentrations. Biochim. Biophys. Acta 57:509–519
Hammond E. M., Giaccia A. J. 2005 The role of p53 in hypoxia-induced apoptosis. Biochem. Biophys. Res. Commun. 331(3):718–725
Janssens, S., K. Theunissen, M. Boogaerts, and F. Van de Werf. Bone marrow cell transfer in acute myocardial infarction. Nature clinical practice 3(Suppl 1):S69–72, March 2006
Jones M., Bonting S. L. 1956 Some relations between growth and carbohydrate metabolism in tissue cultures. Exp. Cell Res. 10(3):631–639
Koh G. Y., Klug M. G., Soonpaa M. H., Field L. J. 1993 Differentiation and long-term survival of C2C12 myoblast grafts in heart. J. Clin. Invest. 92(3):1548–1554
Lamarcq L., Lorimier P., Negoescu A., Labat-Moleur F., Durrant I., Brambilla E. 1995 Comparison of seven bio- and chemiluminescent reagents for in situ detection of antigens and nucleic acids. J. Biolumin. Chemilumin. 10(4):247–256
Lazarus B., Messina A., Barker J. E., Hurley J. V., Romeo R., Morrison W. A., Knight K. R. 2000 The role of mast cells in ischaemia-reperfusion injury in murine skeletal muscle. J. Pathol. 191(4):443–448
Leinwand L. A. 2003 Hope for a broken heart? Cell 114(6):658–659
Lewis J. L., Deloria L. B., Oyen-Tiesma M., Thompson R. C. Jr., Ericson M., Oegema T. R. Jr. 2003 Cell death after cartilage impact occurs around matrix cracks. J. Orthop. Res. 21(5):881–887
Li R. K., Yau T. M., Sakai T., Mickle D. A., Weisel R. D. 1998 Cell therapy to repair broken hearts. Can. J. Cardiol. 14(5):735–744
Linsenmeier R. A., Yancey C. M. 1989 Effects of hyperoxia on the oxygen distribution in the intact cat retina. Invest. Ophthalmol. Vis. Sci. 30(4):612–618
Liu Y., Griffith M., Watsky M. A., Forrester J. V., Kuffova L., Grant D., Merrett K., Carlsson D. J. 2006 Properties of porcine and recombinant human collagen matrices for optically clear tissue engineering applications. Biomacromolecules 7(6):1819–1828
Lundberg P., Kuchel P. W. 1997 Diffusion of solutes in agarose and alginate gels: 1H and 23Na PFGSE and 23Na TQF NMR studies. Magn. Reson. Med. 37(1):44–52
Mangi A. A., Noiseux N., Kong D., He H., Rezvani M., Ingwall J. S., Dzau V. J. 2003 Mesenchymal stem cells modified with Akt prevent remodeling and restore performance of infarcted hearts. Nat. Med. 9(9):1195–1201
McGoron A. J., Nair P., Schubert R. W. 1997 Michaelis–Menten kinetics model of oxygen consumption by rat brain slices following hypoxia. Ann. Biomed. Eng. 25(3):565–572
Meirhaeghe A., Crowley V., Lenaghan C., Lelliott C., Green K., Stewart A., Hart K., Schinner S., Sethi J. K., Yeo G., Brand M. D., Cortright R. N., O’Rahilly S., Montague C., Vidal-Puig A. J. 2003 Characterization of the human, mouse and rat PGC1 beta (peroxisome-proliferator-activated receptor-gamma co-activator 1 beta) gene in vitro and in vivo. Biochem. J. 373(Pt 1):155–165
Menasche P. 2003 Skeletal muscle satellite cell transplantation. Cardiovasc Res. 58(2):351–357
Menasche P. 2003 Myoblast-based cell transplantation. Heart Fail. Rev. 8(3):221–227
Menasche P., Hagege A., Scorsin M., Pouzet B., Desnos M., Duboc D., Schwartz K., Vilquin J., Marroleau J. 2001 Myoblast transplantation for heart failure. Lancet 357(9252):279–280
Mueller-Klieser W., Walenta S. 1993 Geographical mapping of metabolites in biological tissue with quantitative bioluminescence and single photon imaging. Histochem. J. 25(6):407–420
Negoescu A., Guillermet C., Lorimier P., Brambilla E., Labat-Moleur F. 1998 Importance of DNA fragmentation in apoptosis with regard to TUNEL specificity. Biomed. Pharmacother. 52(6):252–258
Negoescu A., Lorimier P., Labat-Moleur F., Drouet C., Robert C., Guillermet C., Brambilla C., Brambilla E. 1996 In situ apoptotic cell labeling by the TUNEL method: improvement and evaluation on cell preparations. J. Histochem. Cytochem. 44(9):959–968
Ott H. C., Davis B. H., Taylor D. A. 2005 Cell therapy for heart failure—muscle, bone marrow, blood, and cardiac-derived stem cells. Semin. Thorac. Cardiovasc. Surg. 17(4):348–360
Radisic M., Malda J., Epping E., Geng W., Langer R., Vunjak-Novakovic G. 2006 Oxygen gradients correlate with cell density and cell viability in engineered cardiac tissue. Biotechnol. Bioeng. 93(2):332–343
Schachinger V., Assmus B., Britten M. B., Honold J., Lehmann R., Teupe C., Abolmaali N. D., Vogl T. J., Hofmann W. K., Martin H., Dimmeler S., Zeiher A. M. 2004 Transplantation of progenitor cells and regeneration enhancement in acute myocardial infarction: final one-year results of the TOPCARE-AMI Trial. J. Am. Coll. Cardiol. 44(8):1690–1699
Schneiderman G., Goldstick T. K. 1978 Oxygen electrode design criteria and performance characteristics: recessed cathode. J. Appl. Physiol. 45(1):145–154
Schwickert G., Walenta S., Mueller-Klieser W. 1996 Mapping and quantification of biomolecules in tumor biopsies using bioluminescence. Experientia 52(5):460–463
Shake, J. G., Gruber, P. J., Baumgartner, W. A., Senechal, G., Meyers, J., Redmond, J. M., Pittenger, M. F., and Martin, B. J. Mesenchymal stem cell implantation in a swine myocardial infarct model: engraftment and functional effects. Ann. Thorac. Surg. 73(6):1919–1925, 2002; discussion 1926
Smits A. M., van Vliet P., Hassink R. J., Goumans M. J., Doevendans P. A. 2005 The role of stem cells in cardiac regeneration. J. Cell Mol. Med. 9(1):25–36
Suzuki K., Smolenski R. T., Jayakumar J., Murtuza B., Brand N. J., Yacoub M. H. 2000 Heat shock treatment enhances graft cell survival in skeletal myoblast transplantation to the heart. Circulation 102(19 Suppl 3):III216–221
Taylor D. A., Atkins B. Z., Hungspreugs P., Jones T. R., Reedy M. C., Hutcheson K. A., Glower D. D., Kraus W. E. 1998 Regenerating functional myocardium: improved performance after skeletal myoblast transplantation. Nat. Med. 4(8):929–933
Taylor B. A., Murray J. D. 1977 Effect of the rate of oxygen consumption on muscle respiration. J. Math. Biol. 4(1):1–20
Thompson R. B., Emani S. M., Davis B. H., van den Bos E. J., Morimoto Y., Craig D., Glower D., Taylor D. A. 2003 Comparison of intracardiac cell transplantation: autologous skeletal myoblasts versus bone marrow cells. Circulation 108(Suppl 1):II264–271
Toma C., Pittenger M. F., Cahill K. S., Byrne B. J., Kessler P. D. 2002 Human mesenchymal stem cells differentiate to a cardiomyocyte phenotype in the adult murine heart. Circulation 105(1):93–98
Van Beek J. H., Loiselle D. S., Westerhof N. 1992 Calculation of oxygen diffusion across the surface of isolated perfused hearts. Am. J. Physiol. 263(4 Pt 2):H1003–1010
Vilquin J. T. 2005 Myoblast transplantation: clinical trials and perspectives. Mini-review. Acta Myol. 24(2):119–127
Walenta S., Chau T. V., Schroeder T., Lehr H. A., Kunz-Schughart L. A., Fuerst A., Mueller-Klieser W. 2003 Metabolic classification of human rectal adenocarcinomas: a novel guideline for clinical oncologists? J. Cancer Res. Clin. Oncol. 129(6):321–326
Whalen W. J., Nair P., Ganfield R. A. 1973 Measurements of oxygen tension in tissues with a micro oxygen electrode. Microvasc. Res. 5(3):254–262
Wollert K. C., Meyer G. P., Lotz J., Ringes-Lichtenberg S., Lippolt P., Breidenbach C., Fichtner S., Korte T., Hornig B., Messinger D., Arseniev L., Hertenstein B., Ganser A., Drexler H. 2004 Intracoronary autologous bone-marrow cell transfer after myocardial infarction: the BOOST randomised controlled clinical trial. Lancet 364(9429):141–148
Woo E. B., Tang A. T., Jarvis J. C., Hasleton P. S., Salmons S., Hooper T. L. 2002 Improved viability of latissimus dorsi muscle grafts after electrical prestimulation. Muscle Nerve 25(5):679–684
Yau T. M., Kim C., Li G., Zhang Y., Weisel R. D., Li R. K. 2005 Maximizing ventricular function with multimodal cell-based gene therapy. Circulation 112(9 Suppl):I123–128
Zhang M., Methot D., Poppa V., Fujio Y., Walsh K., Murry C. E. 2001 Cardiomyocyte grafting for cardiac repair: graft cell death and anti-death strategies. J. Mol. Cell Cardiol. 33(5):907–921
Acknowledgments
This work was supported in part by NHLBI/National Institutes of Health awards to Dr. Taylor (R-01 HL-63346, HL-63703). We would also like to thank Robert Nielsen and Zahid N. Rabbani for their help.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Davis, B.H., Schroeder, T., Yarmolenko, P.S. et al. An In Vitro System to Evaluate the Effects of Ischemia on Survival of Cells Used for Cell Therapy. Ann Biomed Eng 35, 1414–1424 (2007). https://doi.org/10.1007/s10439-007-9301-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10439-007-9301-2