
Abstract
Early life stages of marine calcifiers are particularly vulnerable to climate change. In the Southern Ocean aragonite undersaturation events and areas of rapid warming already occur and are predicted to increase in extent. Here, we present the first study to successfully hatch the polar pteropod Limacina helicina antarctica and observe the potential impact of exposure to increased temperature and aragonite undersaturation resulting from ocean acidification (OA) on the early life stage survival and shell morphology. High larval mortality (up to 39%) was observed in individuals exposed to perturbed conditions. Warming and OA induced extensive shell malformation and dissolution, respectively, increasing shell fragility. Furthermore, shell growth decreased, with variation between treatments and exposure time. Our results demonstrate that short-term exposure through passing through hotspots of OA and warming poses a serious threat to pteropod recruitment and long-term population viability.
Similar content being viewed by others

Explore related subjects
Find the latest articles, discoveries, and news in related topics.Avoid common mistakes on your manuscript.
Introduction
A multitude of concurrent drivers poses a pernicious, global threat to marine ecosystems and their services (Bijma et al. 2013). With increases in temperature of 0.6–2.0 °C, decreases in pH of 0.1–0.4 units and shoaling of the carbonate compensation depth forecasted to occur by 2100 (RCP 2.6–8.5), ocean acidification (OA), and warming pose an acute threat to marine organisms (IPCC 2013; Heinze et al. 2015). Understanding the impact of these environmental perturbations on marine biota remains a major challenge (Kroeker et al. 2013), since a range of intra- and inter-specific responses to multi-stressors have been observed (Wernberg et al. 2012).
Early life history stages are suspected to be especially vulnerable to environmental change (Dupont and Thorndyke 2009), although large variations in tolerance have been observed (Foo and Byrne 2017). Viability and survival of these stages are vital for successful recruitment and long-term population stability. Furthermore, early exposure to environmental stressors could alter vulnerability of later developmental stages through latent effects (such as mortality and shell size), adding to the overall impact (Kroeker et al. 2013; Suckling et al. 2014).
High latitude ecosystems are expected to experience OA first due to seasonal amplification, freshening, and colder temperatures enhancing CO2 solubility (Doney et al. 2009; Fabry et al. 2009). Such processes alter the ratio of dissolved inorganic carbon (DIC) and total alkalinity (TA) to values where calcium carbonate becomes susceptible to dissolution. This poses a significant threat to polar marine organisms that form calcium carbonate shells, skeletons, or internal structures (Mostofa et al. 2016). In the Southern Ocean carbonate undersaturation events have already been observed (Bednaršek et al. 2012a) and are predicted to occur more frequently over the coming decades (McNeil and Matear 2008).
Thecosome pteropods (holoplanktonic gastropods) can dominate high latitude zooplankton communities (Hunt et al. 2008). In polar regions, shelled pteropods are one of the major components within food webs, acting as a food source of carnivorous zooplankton, fishes, and a number of higher predators (Falk-Petersen and Sargent 2001). Furthermore, polar pteropods contribute significantly to carbon and carbonate export to the deep ocean through the sinking of dead individuals and faecal pellets (Manno et al. 2007, 2009). Limacina helicina are true polar pteropods with its two species, Limacina helicina antarctica and Limacina helicina helicina occurring within the Antarctic and Arctic, respectively (Hunt et al. 2010). They are considered sentinels of OA, since their shells consist of aragonite, a relatively soluble polymorph of calcium carbonate (Mucci 1983; Bednaršek et al. 2014). In the Southern Ocean, L. helicina antarctica reside within the surface ocean where aragonite undersaturation events and ‘hotspots’ of rapid warming have already been identified and are predicted to become more frequent (McNeil and Matear 2008; Gutt et al. 2015; Vaughan et al. 2003). This is of particular concern in the northern Scotia Sea region, since it has the largest measured seasonal cycle of surface ocean CO2 in the Southern Ocean (Jones et al. 2012, 2015) as well as upwelling events of CO2 enriched deep water to the surface and hotspots of warming (Bednaršek et al. 2012a; Whitehouse et al. 2008; Gille 2002).
Shell dissolution of juvenile L. helicina antarctica has already been reported in natural populations within the Scotia Sea (Bednaršek et al. 2012a), while numerous incubation experiments under predicted OA levels of polar pteropod juveniles and adults suggest a range of other negative physiological responses (Comeau et al. 2009; Lischka et al. 2011; Manno et al. 2012; Peck et al. 2016a, b; Seibel et al. 2012). Manno et al. (2016) demonstrated that maternal and embryonic exposure of L. helicina antarctica to acidified conditions reduced the percentage of eggs developing to later stages by 80%. However, responses to concurrent OA and warming remain unresolved (Bednaršek et al. 2016a, b) particularly with regard multi-stressor responses of larval L. helicina antarctica. To date, studies of larval pteropods have focussed on incubating North Atlantic and Mediterranean species with Limacina retroversa exhibiting increased mortality (Thabet et al. 2015) and Cavolinia inflexa shell malformations (Comeau et al. 2010a) as a result of ocean acidification. Due to the key ecological and biogeochemical roles of L. helicina antarctica in polar regions, alteration of larval shell morphology alongside reduced recruitment to adulthood could have major implications on the Southern Ocean ecosystem.
In this study, we successfully hatched cultivated polar shelled pteropods (L. helicina antarctica) and used these to examine responses to the singular and combined impact of acidification and warming on posthatch shell development, morphology, and survival. These short-term incubations aim to simulate the experience of larvae (veliger stage) to variations in their environment as a result of present day and predicted heterogeneity in warm and acidified waters in the Southern Ocean (McNeil and Matear 2008; Gutt et al. 2015; Vaughan et al. 2003). Larval survivorship and fitness underpin recruitment success and any negative impacts from a high CO2 world can ultimately reduce long-term population viability in this region (Przeslawski et al. 2008).
Methods
Limacina helicina antarctica were collected aboard the RRS James Clark Ross (Cruise number JR304) within the Scotia Sea (57°36′20.5″S, 43°40′22.2″W) in November 2014 using a motion-compensated Bongo net (100 and 200 µm mesh sizes), vertically hauled from 200 m. The motion compensation reduced stress on pteropods during collection and avoided mechanical damage to shells. Ambient sea-surface conditions at 10 m were characterised by a sea-surface temperature and salinity of 1.62 and 34.3 °C, respectively, with a TA (total alkalinity) of 2320 µmol/kg and a pH (total scale) of 8.09.
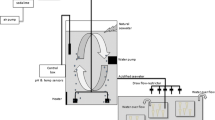
Live adult females were identified following the description of Lalli and Wells (1978) and examined under a light microscope (Olympus SZX16 fitted with a Cannon EOS 60D). Actively swimming individuals with no signs of damage (shell and body) and fully translucent shells were acclimated within filtered seawater (0.22 µm) for 8 h at 1.66 ± 0.03 °C (Spartel incubator with a C-400 circulator unit and an FC-500 in-line cooler, temperature measured every 2 h). After this, individuals that were actively swimming were placed individually within 500 ml incubation jars (non-pyrogenic polystyrene, Corning®) filled with filtered seawater and maintained at 1.22 ± 0.41 °C. Jars were stored in darkness and sealed with no headspace to limit CO2 exchange and inspected at least every 6 h.
After 9 days, some adults spawned eggs within a 2-h period. These were immediately removed using a wide mouthed Pasteur pipette to avoid egg cannibalism and damage. Mothers were in a good state of health during egg production (i.e., swimming and maintaining fully transparent shells) (Peck et al. 2016a, b). Egg ribbons were placed separately into ambient incubatory conditions within 65 ml jars of filtered seawater (1.02 ± 0.31 °C). Following further 7 days of incubation, veligers emerged simultaneously from four egg ribbons, each having been laid by a different female. These were mixed and randomly transferred into experimental conditions via stretched glass pipettes.
Experimental design
All incubations took place within a controlled temperature room aboard the RRS James Clark Ross. Posthatch veligers were examined under a light microscope and five actively swimming individuals with fully translucent shells were placed together into each of the 60 incubation jars (65 ml, non-pyrogenic polystyrene, Corning®). Fifteen jars were placed into each of the four treatments exemplifying ambient (1.7 °C and pH 8.1), warm (3.5 °C and pH 8.1), acidified (1.7 °C and pH 7.6) or acidified-warm (3.5 °C and pH 7.6) conditions (Table 1) based upon predictions in 2100. All seawater was filtered (0.22 µm), since the current evidence suggests L. helicina antarctica spawn in the autumn and overwinter as larvae when food availability is naturally low (Hunt et al. 2008; Lischka and Riebesell 2016). Furthermore, filtration removes the possibility of biological activity altering the carbonate chemistry within the treatment bottles from the target values. To examine the impact of exposure time, three bottles were removed from each treatment every day for 5 days (Supplementary material Figure 1). Each bottle was gently decanted into deep-welled glass petri dishes and veligers were inspected under a light microscope for 5 min each. Those that were actively swimming and/or showed ciliate velum activity were classed as alive. Maximum shell length was measured using a graticule and condition of the larvae noted before preservation, ensuring no secondary preservation effects occurred. All veligers were subsequently rinsed with de-ionised water three times. For preservation, two specimens from each bottle were air dried upon a filter, while the remainder were placed into Eppendorf tubes filled with 70% buffered ethanol.
Ethanol preserved veligers were dehydrated through a series of ethanol solutions (50, 70, 80, 90, 95, and 100%, 5 min each) to stop shell collapse while dried veligers needed no further preparation. All veligers were subsequently mounted on carbon tape and imaged at 1200× magnification using a variable pressure scanning electron microscope (SEM) (TM3000, Hitachi). Only specimens that were living at the end of the incubation were considered for shell analysis (n = 233). Using the SEM, the apical shell surface was inspected for the presence or absence of pitting (deep holes in the shell surface), etching (the partial dissolution of the upper shell surface observed by exposure of the granular or prismatic layer beneath), and malformation (deviation of growth from the expected smooth spiral) (Fig. 4). Since, statistically, there was complete separation in the presence/absence of etching and pitting (for example in etching, presence was either 100 or 0%), this resolution of shell analysis was considered appropriate. The maximum shell diameter was also measured using the SEM and light microscope graticule to approximate shell size over the exposure period.
Seawater manipulation
A Spartel incubator with a C-400 circulator unit and an FC-500 in-line cooler housed within the ship’s cold room was used to control incubation temperature. Temperature was measured every 4 h throughout the entire incubatory period (PreSens fibox 4).
Seawater pH was manipulated through additions of HCl (hydrochloric acid) and NaHCO3 (sodium bicarbonate) calculated by the seacarb software and maintained in a closed system (Lavigne and Gattuso 2010). Gas bubbling and addition of acid/bicarbonate and/or carbonate are considered some of the best methods to mimic ocean acidification (Gattuso et al. 2011). Acid/base addition was used in this instance due to the short incubation period, bottle volumes, and logistical restraints besides minimising the risk of damage to veligers as observed from bubbling (Howes et al. 2014; Gattuso and Lavigne 2009). Although pH may be lower than expected from this technique, we believe that the values are within the range predicted to occur by 2100 (RCP 8.5) in the Southern Ocean (Table 1) (IPCC 2013; Gattuso et al. 2011; Gattuso and Lavigne 2009; SCAR 2009; Schulz et al. 2009; McNeil and Matear 2008). The calculation utilised measurements of pH (total scale) made with a pH electrode (Metrohm 826) and TA through applying a sea-surface salinity (S) and temperature (T) algorithm based on Lee et al. (2006) and refined through a recent spatially intensive carbonate chemistry survey in this region (M.P. Humphreys, pers. comm.):
To determine the impact of the manipulations on the incubation water, we extracted a 250 ml sub-sample of initial incubatory conditions, fixed with mercuric chloride, and stored in a borosilicate bottle for subsequent analysis of TA and DIC. TA was measured by potentiometric titration and DIC by coulometry using a VINDTA (Versatile Instrument for the Determination of Titration Alkalinity, version 3C). Accuracy (TA = 2.5 µmol/kg; DIC = 1.1 µmol/kg) was determined using certified reference materials (Scripps Institution of Oceanography). pH was determined at the start and end of the incubation experiment. Aragonite saturation state was indirectly estimated from TA and DIC values using CO2SYS software with the constants of Mehrbach et al. (1973) refitted by Dickson and Millero (1987) and sulphate dissolution constants by Dickson (1990). Carbonate system parameters of the incubations are shown in Table 1.
Statistical analysis
Data were analysed using R (2015). All larvae were considered when estimating mortality between treatment and days (n = 300) exposed; however, only larvae that were living at the end of the incubatory period and were not damaged during processing were included within the analysis of shell morphology and size (n = 233 where ambient n = 66, acidified n = 44, warm = 59, acidified-warm = 54). A binomial (logit) generalised linear model (GLM) was used to estimate whether there were any differences in etching, pitting, and malformation presence between treatments and days exposed. A gamma (identity) GLM and a binomial (logit) GLM estimated differences in shell size and mortality between treatments and days of exposure, respectively. Complete separation between treatments for the presence of shell etching and pitting was found; therefore, a Bayesian analysis with non-informative prior assumptions (Gelman et al. 2008) was utilised from the arm package (Gelman and Su 2016). Model selection was informed using the information theoretic approach using the MuMIn package (Barton 2016) to identify the models with delta Akaike information criterion < 4 and the highest Akaike weights (Supplementary material Table 8) alongside comparisons of R 2 and likelihood ratio tests using the lmtest package (Zeileis and Hothorn 2002). Model validation included checking assumptions of residual normality and Homoscedasticity, overdispersion, autocorrelation, collinearity, and independence. For post hoc analysis, Tukey’s pairwise comparisons were performed using the lsmeans (Lenth 2002). A two-way factorial analysis was also used within the same model frameworks as above to highlight the presence of any interactions between warming, acidification, and exposure time on larval shell morphology, size, and mortality.
Confidence intervals (CIs) for mean mortality x were calculated by the following:
where n is the number of living larvae per incubation bottle at the start of the incubation, σ the standard deviation, and Zα/2 the Z-table value for a given α value. Confidence intervals for the mean occurrence of shell malformation, pitting, and etching as well as larval mortality were calculated by the modified Wald method (Agresti and Coull 1998).
Results
Change in larval pteropod mortality
Significantly more larval fatalities occurred within warm, acidified, and acidified-warm conditions overall in comparison to ambient treatments (p < 0.01) (Fig. 1a). However, the number of fatalities did not change with the amount of time exposed to these conditions (p > 0.05) (Supplementary materials Table 1). A post hoc analysis showed that throughout, mortality was significantly higher in acidified conditions (38.7%, n = 29) and acidified-warm conditions (25.3%, n = 19) compared to that in warm (12%, n = 9) and ambient conditions (2.7%, n = 2) (p < 0.001). Furthermore, a factorial analysis indicated that acidification (p < 0.001), rather than warming (p > 0.05), increases larval mortality. However, larval mortality significantly increases when warming and acidification are combined (p < 0.01) (Supplementary materials Table 2). For a summary of mortalities, see Supplementary materials Table 3.

Limacina helicina antarctica larval mortality (a) and shell size (b) over time during incubation under ambient (pink), warm (orange), acidified (blue), and acidified-warm (green) conditions. Only larvae that were alive upon harvesting were included in the analysis of shell size. Bars denote 95% confidence intervals between treatment bottles
Change in larval pteropod shell size
Larvae in all treatments significantly increased their shell size during the incubation (p < 0.001). Over the entire exposure time, larvae experiencing acidified (p < 0.05) and acidified-warm (p < 0.01) conditions were smaller than in ambient and warm conditions, which were similar (p > 0.05) (Fig. 1b) (Supplementary materials Table 4). Post hoc analysis indicated that shells incubated in acidified-warm conditions were significantly smaller than those in ambient conditions (p > 0.001) but were not different from those in acidified conditions (p > 0.05). Furthermore, during the first 3 days of exposure, shell size was similar between ambient, warm, and acidified conditions, but subsequently, shell size was smaller in warm and acidified conditions relative to ambient conditions (p < 0.01). The rate of change in shell size was significantly lower in acidified-warm conditions on day 2 (p < 0.05), warm conditions on day 4 and 5 (p < 0.001), and acidified conditions on day 5 (p < 0.05) (Supplementary materials Table 4). This resulted in shell size on day 5 being smaller on exposure to warming (104.5 ± 1.11 µm, n = 13), acidified (106.0 ± 2.0 µm, n = 10), and acidified-warm conditions (105.2 ± 1.2 µm, n = 12) compared to ambient conditions (113.0 ± 0.9 µm, n = 14). The factorial analysis indicated that shell growth was primarily reduced by exposure to acidified conditions (p < 0.001) rather than warm (p > 0.05) with no interaction between them (p > 0.05).
Change in larval pteropod shell morphology
Malformation Significantly more larval shell malformations were present in response to warm and acidified-warm conditions compared to ambient and acidified conditions (p < 0.0001) (Fig. 2a) (Supplementary material Table 6). Post hoc analysis indicated a similar number of larvae developed malformations on exposure to acidified-warm (62.5%, n = 35) and warm conditions (49.2%, n = 29) (p > 0.05). Likewise, there was no difference in the number of malformations between acidified (9.1%, n = 4) and ambient conditions (3%, n = 2) (p > 0.05). Malformations occurred as a result of exposure to warm conditions (p > 0.001) rather than acidification (p > 0.05) with no interaction between them (p > 0.05) (Supplementary material Table 7). The number of shell malformations was highly dependent on the amount of time that the individuals were exposed to each condition (p < 0.0001, n = 223), with larval malformation instances significantly increasing after 3 days of exposure (p < 0.001). There was an 82% increase in malformation occurrence within acidified-warm conditions after the first 3 days of exposure. Furthermore, the number of malformations gradually increased in warming conditions with none being present on day 1–92% being malformed after 5 days.

Limacina helicina antarctica larval shell condition showing percentage occurrence of a malformation, b pitting, and c etching over time during incubation under ambient (pink), warm (orange), acidified (blue), and acidified-warm (green) conditions. Bars denote 95% confidence intervals between treatment bottles. Only larvae that were alive upon harvesting were included (n = 223)
Pitting Larvae that experienced warm, acidified, and acidified-warm conditions all displayed significantly higher amounts of shell pitting than those incubated in ambient conditions (Fig. 2b) (p < 0.001, n = 223) (Supplementary material Table 6). Larval shells with the most pitting (47%, n = 28) were found in warm conditions, although this was not statistically different from the pitting instances on larval shells exposed to acidified-warm (40%, n = 20) or acidified (23%, n = 10) conditions (p > 0.05). Warming and acidification both increased the instances of pitting; however, the combination of acidified-warm conditions does not increase pitting instances as much as would be expected from an additive or a synergistic response (p < 0.01, n = 223) (Supplementary material Table 7). The amount of time larvae were exposed to each condition did not alter the number of pitting instances (p > 0.05, n = 223).
Etching There were significantly more cases of shell etching in acidified and acidified-warm conditions compared to ambient and warm conditions (p < 0.001, n = 223) (Fig. 2c) (Supplementary material Table 6). The presence of etching was attributable to acidification only (p < 0.001, n = 223) with no effect of exposure to warm conditions or an interaction (p > 0.05, n = 223) (Supplementary material Table 7). Larvae incubated in ambient and warm conditions exhibited either no or few cases (2.97%, n = 3) of etching, respectively, with no significant difference between the conditions (p > 0.05). Conversely, after 1 day (24 h) of exposure to acidified and acidified-warm conditions, all larvae had shell etching present, and there was no change in the instances of etching over time (p > 0.05, n = 223).
Overall shell morphology
Those individuals that exhibited etching and malformations together without pitting most frequently occurred within acidified-warm conditions (87%, n = 34), with 10% (n = 4) in acidified and 3% (n = 1) in warm conditions. A similar number of larvae developed malformation and pitting without etching within warm and acidified-warm conditions at 45% (n = 12) and 55% (n = 10), respectively. Larvae developed both shell etching and pitting without malformations within acidified-warm (66%, n = 19) and acidified (35%, n = 10) conditions only. All the larvae that displayed shell pitting, malformation, and etching together occurred within acidified-warm conditions (Fig. 3). The SEM images in Fig. 4 highlight these general combinations of shell morphology.

Larval Limacina helicina antarctica shell morphology over 5 days of exposure to ambient, warm, acidified, and acidified-warm conditions. Each bar shows the treatments where a combination of different shell morphologies (pitting, malformation, and etching) developed on the same single larval shell. Only larvae that were alive upon harvesting were included (n = 223)

Limacina helicina antarctica larval shell morphology as a result of 5 days of exposure to a ambient, b warm, c acidified, and d acidified-warm conditions. Examples of malformation (1), pitting (2) and etching (3) are highlighted with arrows. Larvae were alive upon harvesting
Discussion
Ocean acidification increases larval mortality
We demonstrate that veligers of L. helicina antarctica are sensitive to warm, acidified, and acidified-warm oceanic conditions predicted for 2100 in the Scotia Sea (IPCC 2013; McNeil and Matear 2008), given that there was a high level of larval mortality on exposure to these conditions. The previous studies on Arctic juvenile and adult pteropods incubated in acidified conditions (over 5 and 8 days–1 month) found survivorship of 80–100% (Comeau et al. 2009; Manno et al. 2012; Lischka and Riebesell 2012). Here, we show lower survival (down to 61%) of larval L. helicina antarctica. A similar low survivorship was also found on incubation of larval L. retroversa in acidified conditions indicating increased sensitivity of early life stages (Thabet et al. 2015). This fits the general trend that the early stage molluscs are more vulnerable to acidification than adults (Kroeker et al. 2013; Waldbusser et al. 2015a). Acidification rather than warming appears to be the main driver of increased mortality. Furthermore, there was no synergistic or additive increase in mortality through the addition of warming to acidification. Acidification, therefore, poses the greatest threat to survivorship of larval L. helicina antarctica. However, warming has other sub-lethal influences on shell production and maintenance that may increase vulnerability in the natural environment. Interestingly, mortality did not change with exposure time, suggesting that either fatalities were sensitive phenotypes that would have died regardless of the exposure timeframe or because larvae were more sensitive in the first day of exposure and the more vulnerable died sooner. The short time-scale over which these effects were observed has particular relevance to the environmental experience of pteropods, which are most likely to be exposed to such conditions through contact with mesoscale bodies of water where such altered conditions prevail (Bednaršek et al. 2012a). In the Southern Ocean, pH exhibits spatiotemporal variation with water masses, meltwater, season, and phytoplankton productivity (Kapsenberg et al. 2015; Schram et al. 2015). Furthermore, the continued uptake of anthropogenic CO2 by the surface ocean is predicted to make undersaturation events occur more frequently over the coming decades where water bodies may become corrosive to aragonite during wintertime by 2038 and will be widespread across the Southern Ocean by 2100 (McNeil and Matear 2008).
Ocean acidification and warming decreases shell size
For shell growth to occur, larvae take up carbonate ions from the surrounding seawater and concentrate them within the isolated extrapallial space. With acidification, the concentration of surrounding carbonate ions declines and, therefore, more energy is needed for calcification. Larval shell size increased in all treatments, even when exposed to acidified and warm conditions. Continuing shell calcification, despite exposure to acidified conditions and Ω ar < 1, has also be observed in Arctic L. retroversa (Manno et al. 2012) and L. helicina helicina (Lischka et al. 2011; Comeau et al. 2010b). Previously, Comeau et al. (2009, 2010b) demonstrated that calcification stopped when the saturation state of aragonite was below 0.7 in Arctic L. helicina helicina. Larvae in the present study were incubated at Ω ar = 0.62 and 0.61 in acidified and acidified-warm conditions, respectively, and shell growth continued. This suggests some resilience of these early life stages to short-term exposure to OA (Lischka et al. 2011; Comeau et al. 2010a); however, shell morphology was still altered. Larval shell sizes were smaller upon exposure to acidified and acidified-warm conditions than in ambient conditions. Smaller shell size with exposure to acidification has also been observed in L. helicina juveniles and adults (Comeau et al. 2009; Lischka et al. 2011; Comeau et al. 2012). We found that warming and acidification did not interact and further impact larval shell growth, indicating that warming did not mitigate the impact of OA.
Decreased shell size and delayed growth can be attributed to impeded shell deposition, dissolution exceeding calcification, and reduced energetic capacity (Watson et al. 2009). Since larval shell growth increased on exposure to acidified conditions initially and shell etching was observed throughout the 5-day period, it is unlikely that the mechanistic capacity of shell formation was exceeded. However, increased acid–base regulation in acidified conditions is energetically demanding and may explain smaller shell sizes. Altering external conditions increases the energetic demand of maintaining homeostasis and where these costs cannot be met; complete or partial metabolic suppression may be induced as an adaptive strategy to extend survival time (Pörtner 2008).
Juvenile L. helicina antarctica exposed to acidic conditions suppressed their metabolic rate (Seibel et al. 2012), while Arctic L. helicina helicina exposed to acidified-warm conditions increased their metabolic rate (Lischka and Riebesell 2016). Altering metabolic rate enables energetic allocation to essential physiological processes at the expense of other processes, including shell formation (Pörtner 2008). Food availability has been shown to mediate the impact of ocean acidification in calcifying organisms; therefore, it is possible that with food acquisition, the impacts observed within this study could decline (Seibel et al. 2012; Ramajo et al. 2016). L. helicina antarctica veligers are able to feed directly after hatching and are, therefore, probably not dependent on egg stores (Paranjape 1968; Böer et al. 2005). Current estimations vary in whether L. helicina antarctica overwinter as larvae, when food availability is naturally low or in the summer, when it is high (Hunt et al. 2008; Bednaršek et al. 2012b; Wang et al. 2017). Since shell growth reduced by day 3 and ceased by day 4 in all treatments, we hypothesise that here, larvae may have been initially utilising endogenous reserves and that these were depleted at differing rates between treatments depending on the energetic cost of maintaining homeostasis, finally inducing a stasis in shell size.
Shell morphology is altered in a high CO2 world
We show that larvae incubated under acidified-warm conditions displayed a combination of both shell etching and shell malformation. The previous studies suggested that warming may offset the negative impacts of ocean acidification (sea urchin: Brennand et al. 2010, coral: McCulloch et al. 2012), although others have revealed cumulative (diatom: Boyd et al. 2015) and even synergistic interactions (pteropod: Lischka and Riebesell 2012). Here, we demonstrate that the impacts of warming and acidification on larval shell morphology are separate, with warming initiating shell malformations and acidification resulting in shell etching. This lack of interaction between warming and acidification has also been observed in Arctic L. helicina helicina and L. retroversa juveniles (Lischka et al. 2011; Comeau et al. 2010b; Lischka and Riebesell 2012). It suggests that warming and acidification impact different metabolic processes, thus resulting in malformation and etching, respectively, in pteropod larvae.
Temperature has been shown to have a significant effect on biomineralisation processes and growth across a number of calcifying species (Gazeau et al. 2013). Increased temperature can boost shell growth within an organism’s thermal tolerance window and aid acclimatisation to warming (Somero 2010), but shell microstructure can be altered when this optimum is exceeded (Mackenzie et al. 2014), resulting in shell malformations as observed in the current study (Fig. 1). Regions of strain on a shell, such as areas of attachment, are particularly susceptible to disruption which could explain the banding of malformations occurring parallel to the aperture observed in the current study. Since pitting and malformation occurred in warm and acidified-warm conditions, this suggests that pitting is a result of malformation, perhaps as points of failure where shells surpass their physical limits and causing deformation. Acidification was also shown to cause shell pitting, which is consistent with prior studies (Auzoux-Bordenave et al. 2010; Bednaršek et al. 2012b). Shell pitting can, therefore, occur as a result of two different processes and could signify exposure to acidification, warming, or both. Overall, this demonstrates the structural fragility and loss of integrity of larval shells in high CO2 conditions.
In contrast to warming, the ability of larvae to counteract acidification depends on a combination of the energetic capability to repair shell damage internally (Lischka et al. 2011; Waldbusser et al. 2013, 2015b), the effectiveness and intactness of the protective organic matrix (periostracum) surrounding the shell (Peck et al. 2016a, b), and the ability to regulate ion and acid–base balance to maintain pH at the site of calcification (Thorp and Covich 2009). On exposure of L. helicina antarctica larvae to acidified conditions, shell etching occurred in 100% of individuals from day 1, suggesting a failure in one of these mechanisms. This percentage is higher than previously observed in later life stages (Bednaršek et al. 2012b, 2014; Seibel et al. 2012), indicating that L. helicina antarctica larvae may be particularly sensitive to shell etching, although direct comparison is difficult due to variation in species, origin, and methodology (Gazeau et al. 2013). Many early life history stages of gastropods lack specialised ion-regulatory mechanisms required for acid–base maintenance (Ries 2011a, b). However, since shell size continued to increase and etching occurred on the upper shell surface, it is unlikely that this was the main cause of shell dissolution. Numerous early larval stages of benthic gastropods secrete amorphous calcium carbonate, which is more prone to dissolution, before a transition to aragonite (Weiss et al. 2002; Melzner et al. 2011; Duquette et al. 2017). If this was true for pteropods, it would explain why the protoconches of arctic L. helicina helicina, that represent larval shells, are particularly susceptible to shell dissolution compared to outer whorls formed in later life stages (Peck et al. 2016a, b). Regardless of the shell composition, the periostracum may have been breached, ineffective, or absent for etching to have occurred (Peck et al. 2016a, b), although the exact role of a pteropod’s periostracum as protection against ocean acidification requires further investigation (Ries 2011a, b; Bednaršek et al. 2016a, b). Since etching did not occur in patterns indicative of abrasion or cracking and there was no mechanism for this to occur, it is unlikely that the periostacum was breached. Furthermore, a mechanism allowing isolation of the extrapallial space from the surrounding undersaturated seawater is needed for calcification to proceed, suggesting that a periostracum is present. We suggest that the periostracum is not as developed in the newly hatched larvae as in later life stages of pteropods and is, therefore, inadequate in protecting larval shells from acidification.
The capacity of pteropods to maintain a viable population distribution and abundance in the Southern Ocean depends on their capability to recruit successfully. We showed that OA and warming do not act synergistically, with the nature of the impacts on viability being recognisably different between the two. Survivorship was mainly influenced by the level of acidification, while the effects of warming were more likely to be sub-lethal and did not increase mortality levels when combined with acidification. We demonstrate that the short-term exposures that are likely to be experienced in the natural environment of larval L. helicina antarctica will have a major impact on survivorship and consequently, population stability in these regions.
References
Agresti A, Coull BA (1998) Approximate is better than ‘exact’ for interval estimation of binomial proportions. Am Stat 52(2):119–126
Auzoux-Bordenave S, Badou A, Gaume B, Berland S, Helléouet M, Milet C, Huchette S (2010) Ultrastructure, chemistry and mineralogy of the growing shell of the European abalone Haliotis tuberculata. J Struct Biol 171:277–290
Barton K (2016) MuMIn: multi-model inference. R package version 1.15.6. https://CRAN.R-project.org/package=MuMIn. Accessed 03 May 2016
Bednaršek N, Tarling GA, Bakker DCE, Fielding S, Cohen A, Kuzirian A, McCorkle D, Lézé B, Montagna R (2012a) Description and quantification of pteropod shell dissolution: a sensitive bioindicator of ocean acidification. Glob Change Biol 18(7):2378–2388
Bednaršek N, Tarling GA, Bakker DCE, Fielding S, Jones EM, Venables HJ, Ward P, Kuzirian A, Lézé B, Feely RA, Murphy EJ (2012b) Extensive dissolution of live pteropods in the Southern Ocean. Nat Geosci 5(12):881–885
Bednaršek N, Tarling GA, Bakker DCE, Fielding S, Feely RA (2014) Dissolution dominating calcification process in polar pteropods close to the point of aragonite undersaturation. PLoS One 9(10):e109183. https://doi.org/10.1371/journal.pone.0109183
Bednaršek N, Harvey CJ, Kaplan IC, Feely RA, Moz J (2016a) Pteropods on the edge: cumulative effects of ocean acidification, warming and deoxygenation. Prog Oceanogr 145:1–24
Bednaršek N, Johnson J, Feely RA (2016b) Comment on Peck et al.: vulnerability of pteropod (Limacina helicina) to ocean acidification: shell dissolution occurs despite an intact organic layer. Deep Sea Res Part II 127:53–56
Bijma J, Pörtner HO, Yesson C, Rogers AD (2013) Climate change and the oceans—what does the future hold? Mar Pollut Bull 74(2):495–505
Böer M, Gannefors C, Kattner G, Graeve M, Hop H, Falk-Petersen S (2005) The Arctic pteropod Clione limacina: seasonal lipid dynamics and life-strategy. Mar Biol 147(3):707–717
Boyd PW, Dillingham PW, McGraw CM, Armstrong EA, Cornwall CE, Feng YY, Hurd CL, Gault-Ringold M, Roleda MY, Timmins-Schiffman E, Nunn BL (2015) Physiological responses of a Southern Ocean diatom to complex future ocean conditions. Nature Clim Change 6:207–213
Brennand HS, Soars N, Dworjanyn SA, Davis AR, Byrne M (2010) Impact of ocean warming and ocean acidification on larval development and calcification in the sea urchin Tripneustes gratilla. PLoS One 5(6):e11372. https://doi.org/10.1371/journal.pone.0011372
Comeau S, Gorsky G, Jeffree R, Teyssie JL, Gattuso JP (2009) Impact of ocean acidification on a key Arctic pelagic mollusc (Limacina helicina). Biogeosciences 6(9):1877–1882
Comeau S, Gorsky G, Alliouane S, Gattuso JP (2010a) Larvae of the pteropod Cavolinia inflexa exposed to aragonite undersaturation are viable but shell-less. Mar Biol 157(10):2341–2345
Comeau S, Jeffree R, Teyssié JL, Gattuso JP (2010b) Response of the Arctic pteropod Limacina helicina to projected future environmental conditions. PLoS One 5(6):e11362. https://doi.org/10.1371/journal.pone.0011362
Comeau S, Alliouane S, Gattuso J, June P, Pen O, Alliouane S, Gattuso J (2012) Effects of ocean acidification on overwintering juvenile Arctic pteropods Limacina helicina. Mar Ecol Prog Ser 456:279–284
Dickson AG (1990) Standard potential of the reaction: AgCl(s) + 1/2 H2(g) = Ag(s) + HCl(aq), and the standard acidity constant of the ion HSO4 − in synthetic sea water from 273.15 to 318.15 K. K. J Chem Thermodyn 22:113–127
Dickson AG, Millero FJ (1987) Comparison of the equilibrium constants for the dissociation of carbonic acid in seawater media. Deep Sea Res Part I 34(111):1733–1743
Doney SC, Fabry VJ, Feely RA, Kleypas JA (2009) Ocean acidification: the other CO2 problem. Annu Rev Mar Sci 1(1):169–192
Dupont S, Thorndyke MC (2009) Impact of CO2-driven ocean acidification on invertebrate’s early life-history-what we know, what we need to know and what we can do. Biogeosciences 6:3109–3131
Duquette, A McClintock JB, Amsler CD, Pérez-Huerta A, Milazzo M, Hall-Spencer JM (2017) Effects of ocean acidification on the shells of four Mediterranean gastropod species near a CO2 seep. Mar Pollut Bull (in press)
Fabry VJ, McClintock JB, Mathis JT, Grebmeier JM (2009) Ocean acidification at high latitudes: the bellwether. Oceanography 22(4):160–171
Falk-Petersen S, Sargent JR (2001) Lipids and fatty acids in Clione limacina and Limacina helicina in Svalbard waters and the Arctic Ocean: trophic implications. Polar Biol 24(3):163–170
Foo AF, Byrne M (2017) Marine gametes in a changing ocean: impacts of climate change stressors on fecundity and the egg. Mar Environ Res. https://doi.org/10.1016/j.marenvres.2017.02.004 (in press)
Gattuso JP, Lavigne H (2009) Approaches and software tools to investigate the impact of ocean acidification. Biogeosciences 6(10):2121–2133
Gattuso JP, Gao K, Lee K, Rost B, Schulz KG (2011) Approaches and tools to manipulate the carbonate chemistry. In: Riebesell U, Fabry VJ, Hansson L, Gattuso JP (eds) Guide to best practices for ocean acidification research and data reporting. Publications Office of the European Union, Luxembourg, pp 243–258
Gazeau F, Parker LM, Comeau S, Gattuso JP, O’Connor WA, Martin S, Pörtner HO, Ross PM (2013) Impacts of ocean acidification on marine shelled molluscs. Mar Biol 160(8):2207–2245
Gelman A, Su Y (2016) Arm: data analysis using regression and multilevel/hierarchical models; R package version 1.8-6. Cambridge University Press, Cambridge
Gelman A, Jakulin A, Pittau MG, Su YS (2008) A weakly informative default prior distribution for logistic and other regression models. Ann Appl Stat 2(4):1360–1383
Gille ST (2002) Warming of the Southern Ocean since the 1950s. Science 295(5558):1275–1277
Gutt J, Bertler N, Bracegirdle TJ, Buschmann A, Comiso J, Hosie G, Isla E, Schloss IR, Smith CR, Tournadre J, Xavier JC (2015) The Southern Ocean ecosystem under multiple climate change stresses-an integrated circumpolar assessment. Glob Change Biol 21(4):1434–1453
Heinze C, Meyer S, Goris N, Anderson L, Steinfeldt R, Chang N, Le Quéré C, Bakker DCE (2015) The ocean carbon sink: impacts, vulnerabilities and challenges. Earth Syst Dyn 6(1):327–358
Howes EL, Bednaršek N, Büdenbender J, Comeau S, Doubleday A, Gallager SM, Hopcroft RR, Lischka S, Maas AE, Bijma J, Gattuso JP (2014) Sink and swim: a status review of thecosome pteropod culture techniques. J Plankton Res 36(2):299–315
Hunt BPV, Pakhomov EA, Hosie GW, Siegel V, Ward P, Bernard K (2008) Pteropods in Southern Ocean ecosystems. Prog Oceanogr 78(3):193–221
Hunt B, Strugnell J, Bednarsek N, Linse K, Nelson RJ, Pakhomov E, Seibel B, Steinke D, Würzberg L (2010) Poles apart: the “bipolar” pteropod species Limacina helicina is genetically distinct between the Arctic and Antarctic oceans. PLoS One 5(3):e9835
IPCC (2013) In: Stocker TF, Qin D, Plattner GK, Tignor M, Allen SK, Boschung J, Nauels A, Xia Y, Bex V, Midgley PM (eds) Climate change 2013: the physical science basis. Contribution of working group I to the fifth assessment report of the intergovernmental panel on climate change. Cambridge University Press, Cambridge, UK and New York, NY, USA. https://doi.org/10.1017/CBO9781107415324
Jones EM, Bakker DCE, Venables HJ, Watson AJ, Georgia S (2012) Dynamic seasonal cycling of inorganic carbon downstream of South Georgia, Southern Ocean. Deep Sea Res Part II 59–60:25–35
Jones EM, Bakker DCE, Venables HJ, Hardman-Mountford NJ (2015) Seasonal cycle of CO2 from the sea ice edge to island blooms in the Scotia Sea, Southern Ocean. Mar Chem 177:490–500
Kapsenberg L, Kelley AL, Shaw EC, Martz TR, Hofmann GE (2015) Near-shore antarctic pH variability has implications for the design of ocean acidification experiments. Sci Rep 5:9638
Kroeker KJ, Kordas RL, Crim R, Hendriks IE, Ramajo L, Singh GS, Duarte CM, Gattuso JP (2013) Impacts of ocean acidification on marine organisms: quantifying sensitivities and interaction with warming. Glob Change Biol 19(6):1884–1896
Lalli CM, Wells FE (1978) Reproduction in the genus Limacina (Opisthobranchia: Thecosomata). J Zool 186:95–108
Lavigne H, Gattuso JP (2010) Seacarb: seawater carbonates chemistry with R package version 3.1.3. http://CRAN.R-project.org/package=seacarb. Accessed 03 May 2016
Lee RF, Hagen W, Kattner G (2006) Lipid storage in marine zooplankton. Mar Ecol Prog Ser 307(1863):273–306
Lenth R (2002) lsmeans: least-squares means. R package version 2.20-23. http://CRAN.R-project.org/package=lsmeans. Accessed 03 May 2016
Lischka S, Riebesell U (2012) Synergistic effects of ocean acidification and warming on overwintering pteropods in the Arctic. Glob Change Biol 18(12):3517–3528
Lischka S, Riebesell U (2016) Metabolic response of Arctic pteropods to ocean acidification and warming during the polar night/twilight phase in Kongsfjord (Spitsbergen). Polar Biol. https://doi.org/10.1007/s00300-016-2044-5
Lischka S, Büdenbender J, Boxhammer T, Riebesell U (2011) Impact of ocean acidification and elevated temperatures on early juveniles of the polar shelled pteropod Limacina helicina: mortality, shell degradation, and shell growth. Biogeosciences 8:919–932
Mackenzie CL, Ormondroyd GA, Curling SF, Ball RJ, Whiteley NM, Malham SK (2014) Ocean warming, more than acidification, reduces shell strength in a commercial shellfish species during food limitation. PLoS One 9(1):e86764. https://doi.org/10.1371/journal.pone.0086764
Manno C, Sandrini S, Tositti L, Accornero A (2007) First stages of degradation of Limacina helicina shells observed above the aragonite chemical lysocline in Terra Nova Bay (Antarctica). J Mar Syst 68(1–2):91–102
Manno C, Accornero A, Umani SF (2009) Importance of the contribution of Limacina helicina faecal pellets to the carbon pump in Terra Nova Bay (Antarctica). J Plankton Res 32(2):145–152
Manno C, Morata N, Primicerio R (2012) Limacina retroversa’s response to combined effects of ocean acidification and sea water freshening. Estuar Coast Shelf Sci 113:163–171
Manno C, Peck VL, Tarling GA (2016) Pteropod eggs released at high pCO2 lack resilience to ocean acidification. Sci Rep 6:e25752. https://doi.org/10.1038/srep25752
McCulloch M, Falter J, Trotter J, Montagna P (2012) Coral resilience to ocean acidification and global warming through pH up-regulation. Nat Clim Change 2(4):1–5
McNeil BI, Matear RJ (2008) Southern Ocean acidification: a tipping point at 450-ppm atmospheric CO2. Proc Natl Acad Sci USA 105(48):18860–18864
Mehrbach C, Culberson CH, Hawley JE, Pytkowicx RM (1973) Measurement of the apparent dissociation constants of carbonic acid in seawater at atmospheric pressure. Limnol Oceanogr 18:897–907
Melzner F, Stange P, Trübenbach K, Thomsen J, Casties I, Panknin U, Gorb SN, Gutowska MA (2011) Food supply and seawater p CO2 impact calcification and internal shell dissolution in the blue mussel Mytilus edulis. PLoS One 6(9):e24223
Mostofa KMG, Liu C, Zhai W, Minella M, Vione D, Gao K, Minakata D, Arakaki T, Yoshioka T, Hayakawa K, Konohira E (2016) Reviews and syntheses: ocean acidification and its potential impacts on marine ecosystems. Biogeosciences 13:1767–1786
Mucci A (1983) The solubility of calcite and aragonite in seawater at various salinities, temperatures, and one atmosphere total pressure. Am J Sci 283:780–799
Paranjape M (1968) The egg mass and veligers of Limacina helicina Phipps. Veliger 10:322–326
Peck VL, Tarling GA, Manno C, Harper EM (2016a) Reply to comment by Bednaršek. Deep Sea Res Part II 127:57–59
Peck VL, Tarling GA, Manno C, Harper EM, Tynan E (2016b) Outer organic layer and internal repair mechanism protects pteropod Limacina helicina from ocean acidification. Deep Sea Res Part II 127:41–52
Pörtner HO (2008) Ecosystem effects of ocean acidification in times of ocean warming: a physiologist’s view. Mar Ecol Prog Ser 373:203–217
Przeslawski R, Ahyong S, Byrne M, Worheides G, Hutchings P (2008) Beyond corals and fish: the effects of climate change on noncoral benthic invertebrates of tropical reefs. Glob Change Biol 14:2773–2795
Ramajo L, Pérez-León E, Hendriks IE, Marba N, Krause-Jensen D, Sejr MK, Blicher ME, Lagos NA, Olsen YS, Duarte CM (2016) Food supply confers calcifiers resistance to ocean acidification. Sci Rep. https://doi.org/10.1038/srep19374
Ries JB (2011a) A physicochemical framework for interpreting the biological calcification response to CO2 induced ocean acidification. Geochim Cosmochim Acta 75(14):4053–4064
Ries JB (2011b) Skeletal mineralogy in a high CO2 world. J Exp Mar Biol Ecol 403(1–2):54–64
Schram JB, Schoenrock KM, McClintock JB, Amsler CD, Angus RA (2015) Multi-frequency observations of seawater carbonate chemistry on the central coast of the western Antarctic Peninsula. Polar Res 34(1):25582
Schulz KG, Barcelos e Ramos J, Zeebe RE, Riebesell U (2009) CO2 perturbation experiments: similarities and differences between dissolved inorganic carbon and total alkalinity manipulations. Biogeosciences 6(10):2145–2153
Seibel BA, Maas AE, Dierssen HM (2012) Energetic plasticity underlies a variable response to ocean acidification in the pteropod, Limacina helicina antarctica. PLoS One 7(4):e30464
Somero GN (2010) The physiology of climate change: how potentials for acclimatization and genetic adaptation will determine ‘winners’ and ‘losers’. J Exp Biol 213(6):912–920
Suckling CC, Clark MS, Beveridge C, Brunner L, Hughes AD, Harper EM, Cook EJ, Davies AJ, Peck LS (2014) Experimental influence of pH on the early life-stages of sea urchins II: increasing parental exposure times gives rise to different responses. Invertebr Reprod Dev 58(3):161–175
R Development Core Team (2015) R: a language and environment for statistical computing. R Foundation for Statistical Computing, version 3.1.3. https://cran.r-project.org. Accessed 03 May 16
Thabet AA, Maas AE, Lawson GL, Tarrant AM (2015) Life cycle and early development of the thecosomatous pteropod Limacina retroversa in the Gulf of Maine, including the effect of elevated CO2 levels. Mar Biol 162(11):2235–2249
Thorp JH, Covich AP (2009) Ecology and classification of North American freshwater invertebrates. Academic press
Vaughan DG, Marshall GJ, Connolley WM, Parkinson C, Mulvaney R, Hodgson DA, King JC, Pudsey CJ, Turner J (2003) Recent rapid regional climate warming on the Antarctic Peninsula. Clim Change 60(3):243–274
Waldbusser GG, Brunner EL, Haley BA, Hales B, Langdon CJ, Prahl FG (2013) A developmental and energetic basis linking larval oyster shell formation to acidification sensitivity. Geophys Res Lett 40:2171–2176
Waldbusser GG, Hales B, Langdon CJ, Haley BA, Schrader P, Brunner EL, Gray MW, Miller CA, Gimenez I (2015a) Saturation-state sensitivity of marine bivalve larvae to ocean acidification. Nat Clim Change 5:273–280
Waldbusser GG, Hales B, Langdon CJ, Haley BA, Schrader P, Brunner EL, Gray MW, Miller CA, Gimenez I, Hutchinson G (2015b) Ocean acidification has multiple modes of action on bivalve larvae. PLoS One 10:e0128376
Wang K, Hunt BP, Liang C, Pauly D, Pakhomov EA (2017) Reassessment of the life cycle of the pteropod Limacina helicina from a high resolution interannual time series in the temperate North Pacific. ICES J Mar Sci. https://doi.org/10.1093/icesjms/fsx014
Watson SA, Southgate PC, Tyler PA, Peck LS (2009) Early larval development of the Sydney rock oyster Saccostrea glomerata under near-future predictions of CO2 driven ocean acidification. J Shellfish Res 28(3):431–437
Weiss IM, Tuross N, Addadi LIA (2002) Mollusc larval shell formation: amorphous calcium carbonate is a precursor phase for aragonite. J Exp Zool 491:478–491
Wernberg T, Smale DA, Thomsen MS (2012) A decade of climate change experiments on marine organisms: procedures, patterns and problems. Glob Change Biol 18(5):1491–1498
Whitehouse MJ, Meredith MP, Rothery P, Atkinson A, Ward P, Korb RE (2008) Rapid warming of the ocean around South Georgia, Southern Ocean, during the 20th century: forcings, characteristics and implications for lower trophic levels. Deep Sea Res Part I 55(10):1218–1228
Zeileis A, Hothorn T (2002) Diagnostic checking in regression relationships. R News 2(3):7–10
Acknowledgements
We thank the captain and the crew of the RRS James Clark Ross for their support in all the logistical operations on board. G. A. Lee, S. Humphrey, and O. Legge helped with carbonate chemistry analysis. M. Humphreys provided algorithms to determine total alkalinity. K. Sales gave advice on statistical analysis.
Author information
Authors and Affiliations
Contributions
CM conceptualised the project. JG wrote the manuscript. JG, GT, and VP carried out the fieldwork. CM, GT, VP, and DB provided theoretical overviews and help in the writing of the manuscript. DB aided with the carbonate chemistry analysis.
Corresponding author
Ethics declarations
Funding
This work was carried out as part of the EnvEast Doctoral training partnership (NE/L002582/1) and the ecosystems programme at the British Antarctic Survey during a Western Corebox cruise.
Ethical approval
All applicable international, national, and institutional guidelines for the care and use of animals were followed.
Conflict of interest
The authors declare that they have no conflict of interest or competing financial interests.
Additional information
Responsible Editor: H.-O. Pörtner.
Reviewed by J. B. McClintock and an undisclosed expert.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Gardner, J., Manno, C., Bakker, D.C.E. et al. Southern Ocean pteropods at risk from ocean warming and acidification. Mar Biol 165, 8 (2018). https://doi.org/10.1007/s00227-017-3261-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00227-017-3261-3