
Abstract
Aims/hypothesis
The metabolic syndrome, with associated vasculopathy, is a major cause of cardiovascular disease and nephropathy. Impaired nitric oxide (NO) metabolism and endothelial function is an important component of the disease process. Increasing the availability of arginine, the precursor of NO, might enhance vascular function and protect against end-stage disease.
Materials and methods
Insulin-resistant JCR:LA-cp rats were treated with arginine–silicate–inositol complex or arginine–HCl at 1.0 g kg−1 day−1 (expressed as arginine–HCl) from 8 to 13 weeks of age. The contractile/relaxant function of thoracic aortae and coronary arteries was assessed in vitro. Kidneys were assessed for severity of glomerular sclerosis.
Results
Arginine–silicate complex, but not arginine–HCl, normalised the hypercontractile response of the aorta to phenylephrine via an NO-dependent pathway. Coronary artery function, as indicated by reactive hyperaemia to warm ischaemia, was enhanced by both arginine compounds. In addition, the arginine–silicate complex increased coronary vasodilatation in response to bradykinin. Glomerular sclerosis was significantly reduced in rats treated with the arginine–silicate complex.
Conclusions/interpretation
Treatment with exogenous arginine, in an efficiently absorbed form, improves vascular function and reduces nephropathy in an animal model of insulin resistance and cardiovascular disease, via mechanism(s) independent of insulin concentration. Enhancement of NO metabolism through increased availability of the precursor arginine appears to offer protection against micro- and macrovascular disease associated with the metabolic syndrome and insulin resistance.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The metabolic syndrome is an increasingly common cause of cardiovascular morbidity and mortality worldwide [1, 2]. A crucial feature of this disorder is endothelial and vascular dysfunction that develops during the prediabetic phase of type 2 diabetes. The concomitant insulin resistance and hyperinsulinaemia appear to be a major determinant of early-stage vasculopathy, atherosclerosis and ischaemic cardiovascular disease [3, 4]. Effective reduction in early-phase vascular dysfunction, including aortic responsiveness/contractility, offers potential protection against cardiovascular disease and mortality [5]. Recent advances in pharmaceutical intervention, including novel insulin sensitisers, have reduced insulin levels and facilitated the reduction of vasculopathy and endothelial dysfunction [6, 7]. However, current insulin-sensitising compounds, such as troglitazone, are often responsible for a range of additional effects that are not well tolerated in the clinical setting [8]. It also remains unclear whether early-phase vasculopathy can be reduced directly without the use of the intensive pharmaceutical regimes required to treat the fully developed metabolic syndrome or type 2 diabetes. Consequently, there is a potentially significant role for novel pharmacological and nutritional interventions that target early-phase vasculopathy directly [8].
The JCR:LA-cp rat is a unique strain that spontaneously develops the range of dysfunction and pathophysiology associated with the metabolic syndrome in humans. Rats that are homozygous for the autosomal recessive cp gene (cp/cp) develop obesity, hyperlipidaemia, profound insulin resistance, glomerular sclerosis and atherosclerosis with enhanced vascular contractility and reduced vascular relaxation [9–14]. Animals that are heterozygous (cp/+) or homozygous normal (+/+) are indistinguishable, lean and metabolically normal. Most importantly, the cp/cp male phenotype spontaneously develops ischaemic lesions of the heart and is prone to stress-induced myocardial infarcts that can be fatal [15], a common clinical complication of the metabolic syndrome. Insulin resistance develops rapidly in young male rats between 4 and 7 weeks of age and is highly correlated with cardiovascular disease in this strain [10]. In addition, recent pilot studies have shown that the male cp/cp rat has increased urinary albumin excretion at 12 weeks of age, when insulin levels are very high, consistent with the presence of glomerular sclerosis in young adult (12 weeks of age) cp/cp rats (Russell et al. unpublished observations).
A number of interventions including MEDICA 16, d-fenfluramine, and the novel agent, S15261, have been shown to reduce insulin resistance and lower plasma insulin levels in the cp/cp rat with associated reduction of vasculopathy and cardiac dysfunction [6, 7, 16]. A significant reduction of insulin levels in the cp/cp rat is associated with improved vascular function, reduced atherosclerosis and prevention of ischaemic lesions of the heart. Since the vascular dysfunction of the cp/cp rat is related to impaired nitric oxide (NO) metabolism [14], improvement in NO availability might result in functional improvement in the vascular system. NO is derived from arginine as a precursor through the action of NO synthase (NOS) [17], suggesting that arginine administration could protect against cardiovascular disease in the presence of insulin resistance. To date, the clinical efficacy of dietary arginine supplementation has been limited, due to poorly absorbed formulations with a consequently low bioavailability of arginine. We have investigated the efficacy of a novel arginine–silicate–inositol complex that was developed to enhance the systemic bioavailability of arginine in vivo. The silicate–inositol complex has physicochemical properties that increase the rate of absorption of arginine across the intestinal tract and significantly raise the concentration of available arginine in the circulation, compared with conventional preparations (such as arginine–HCl). The arginine–silicate complex has been described and shown to be non-toxic in several ex vivo test systems [18–20]. We hypothesised that the beneficial effects of arginine supplementation would occur independently of any improvement in insulin sensitivity. The present study assessed the efficacy of the novel arginine–silicate–inositol complex in improving the function of aortic and coronary arteries and attenuating progression of nephropathy in the JCR:LA-cp rat.
Materials and methods
Animals and treatment
Male rats of the JCR:LA-cp strain, obese (cp/cp) and lean (+/?, a 2:1 mix of heterozygotes [cp/+] and homozygote normals [+/+]), were raised and housed in our original breeding colony at the University of Alberta, as described previously [16]. The strain has recently been rederived and established at Charles River Laboratories (Wilmington, MA, USA) with the designation Crl:JCR(LA)-Lepr cp. The care of the animals and experimental procedures were in conformity with the guidelines of the Canadian Council on Animal Care and subject to prior institutional review and approval.
At 7 weeks of age, the animals were conditioned to a sham tail-bleeding procedure and the cp/cp rats randomised to either one of two treatment diets or control diet. Treatment diets were supplemented with either arginine–HCl at a dose of 1 g kg−1 day−1 or arginine–silicate–inositol complex at a dose of 1.81 g kg−1 day–1 (equivalent arginine dose per body weight) and provided to the animals from 8 to 13 weeks of age. Rats were weighed and their food consumption measured twice a week during the intervention period. Arginine preparations were incorporated into powdered feed and the concentration was adjusted weekly so as to maintain the desired dose of each agent on a g/kg body weight basis [7]. Lean (+/?) and obese (cp/cp) control groups were given feed that was prepared using the same protocol with the exception of incorporated arginine.
Insulin and glucose
The insulin and glucose metabolism of the cp/cp rat is abnormally responsive to stress or disturbance. We have established a procedure to collect blood samples from conscious rats under a specific protocol during the dark (active) phase of their diurnal cycle in order to reduce stress and associated variability. At 12 weeks of age, the animals were deprived of food for 16 h, warmed on a heated table to ensure vasodilatation of the tail and 0.5 ml blood was taken from the tip of the tail of conscious rats. The rats were killed 1 week later (at 13 weeks of age), blood was collected for separation of plasma, urine was sampled directly from the bladder for assay of albumin:creatinine and tissues were taken for vascular function studies.
Vascular function
The vascular function of aortic rings with intact endothelium was assessed using established methods [21]. Briefly, rats were anaesthetised using isofluorane in oxygen. The chest cavity was exposed and the thoracic aorta excised, trimmed of adhering fat and connective tissue and cut into 3-mm-long transverse rings. Aortic rings were mounted on stainless steel hooks under 1.5-g resting tension in 10-ml organ baths and bathed at 37°C in Krebs solution (containing in mmol/l: NaCl 116, KCl 5.4, CaCl2 1.2, MgCl2 2.0, Na2PO4 1.2, glucose 10 and NaHCO3 19) and gassed with 95% O2 and 5% CO2. Tension was recorded isometrically with Grass FTO3C transducers (Grass Medical Instruments, Quincy, MA, USA) and displayed on a Digi-Med tissue-force analyser (Model 210; Micro-Med, Louisville, KY, USA) linked to an IBM-compatible computer that acquired data digitally using DMSI 210/4 (Micro-Med) software.
The contractile response of endothelium-intact rings of aortae to phenylephrine (PE) was assessed through concentration-response curves for PE (1 to 300 mol/l). The basal NO-mediated relaxation of aortic rings (pre-contracted with PE to 80% of maximal contraction) was assessed by determining the concentration response to the endothelial NO-releasing agent acetylcholine (ACh) and the NO donor sodium nitroprusside (SNP). Direct assessment of NO-mediated effects was also determined through addition of N G-nitro-l-arginine methyl ester (l-NAME), at 10−4mol/l, in order to inhibit NOS activity [17].
Heart perfusions
Hearts were excised concurrently with aortae, cannulated and perfused at 100 mmHg constant pressure on an Isolated Heart Apparatus (Size 3) equipped with a Type 700 transit time flowmeter (Hugo Sachs Elektronik-Harvard Apparatus, March-Hugstetten, Germany). The hearts were perfused with Krebs–Henseleit solution (containing in mmol/l: NaCl 118, KCl 4.7, CaCl2 1.2, MgCl2 2.0, Na2HPO4 1.2, glucose 10 and NaHCO3 19) and gassed with 95% O2 and 5% CO2. Bradykinin was added to the perfusate upstream to the debubbling chamber using a calibrated roller pump (Varioperpex II Pump 2120; LKB-Produkter, Bromma, Sweden). The following variables were collected: heart rate, perfusion pressure and coronary flow using data acquisition software (Isoheart W program V 1.2; Hugo Sachs).
Materials
Rat chow was Lab Diet 5001 (PMI Nutrition International, Brentwood, MO, USA) throughout. Arginine–silicate–inositol complex was supplied by Nutrition 21, Purchase, NY, USA. Reagents and chemicals, including arginine–HCl, ACh, bradykinin, PE and SNP, were obtained from Sigma Chemical (Oakville, ON, Canada).
Analytical methods
Plasma glucose was measured by the use of a rapid glucose oxidase technique (Beckman Instruments, Brea, CA, USA). Insulin was assayed by a double antibody RIA technique (Kabi Pharmacia, Uppsala, Sweden) and rat insulin standards. Urine albumin and creatinine measurements were performed on a Beckman Coulter LX20i analyser using immuno-turbidimetric and Jaffé methods, respectively. Plasma arginine concentrations were measured using a Beckman 6300 Amino Acid Analyser.
Histology
Kidneys were fixed in formalin and subjected to conventional processing and sectioning, followed by haematoxylin and eosin (H&E) staining. The extent of glomerular sclerosis was determined using a similar process to that of Shäfer et al. [22]. Four fields of view of the right kidney of each rat were examined at ×100 and all glomeruli rated as normal or sclerotic. Results were expressed as the percent of glomeruli that exhibited sclerosis.
Statistical analysis
Results are expressed as means±SEM and were analysed using SigmaStat (Jandel Scientific, San Rafael, CA, USA) and plotted using SigmaPlot (SPSS, Chicago, IL, USA) and Prism (Graphpad, San Diego, CA, USA). Data were compared using one-way ANOVA followed by multiple comparison tests. Concentration-response curves were analysed using the program ALLFIT [23], which fits the complete dataset to the logistic equation and permits independent testing of differences between individual parameters. A value of p<0.05 was taken as being statistically significant.
Results
Food intake and body weight
The intake of food by rats treated with either arginine–HCl or arginine–silicate complex did not differ from controls throughout the treatment period (37.9±0.8 vs 37.4±1.1 g/day for control and arginine–silicate groups, respectively, at 12 weeks of age, p>0.05). Similarly, arginine supplementation had no effect on body weight at 12 weeks of age (480±6.7, 478±7.8 and 485±12 g for control, arginine–silicate and arginine–HCl, respectively).
Plasma arginine concentrations
The plasma concentrations of arginine in untreated +/? and cp/cp control rats were not significantly different. Rats treated with the arginine–silicate complex had significantly higher plasma concentrations of arginine than the cp/cp controls (1.45±0.09 vs 1.18±0.05 nmol/l, p<0.05), In contrast, rats treated with arginine–HCl had non-significantly higher concentrations (1.33±0.06 nmol/l, p>0.05), despite the equivalent dose of arginine for each treatment.
Plasma insulin and glucose concentrations
Male cp/cp rats are hyperinsulinaemic and had higher fasting insulin levels than +/? controls (304±29 vs 18±4 mU/l, p<0.001). Our hypothesis was that beneficial effects of arginine supplementation would occur independently of any improvement in insulin sensitivity. In fact, fasted cp/cp rats treated with the arginine–silicate complex (but not arginine–HCl) had significantly higher plasma insulin levels than controls (485±70 mU/l, p<0.05, and 453±58 mU/l, p=0.054, respectively). Fasting plasma glucose concentrations of the cp/cp rats were not significantly affected by the arginine supplementation (7.94±0.02 and 8.55±0.03 mmol/l for the arginine–silicate complex and arginine–HCl, respectively, vs 7.88±0.03 mmol/l for cp/cp control rats).
Aortic vascular function
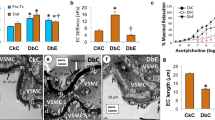
The contractile dose-response of aortic rings to PE are shown in Fig. 1, with corresponding calculated values for parameters of the logistic equation curve presented in Table 1. In the absence of l-NAME, the maximum contractile response of the cp/cp control aortae was significantly greater than that of the +/? aortae, consistent with previous reports [14] (Fig. 1, left panel). Treatment of the cp/cp rats with the arginine–silicate complex (but not arginine–HCl) reduced the maximum PE-mediated contraction to that of the +/? controls (p<0.0001, vs cp/cp control). However, there was no statistically significant difference between the EC50 values for PE for any of the experimental groups. The contractile response of aortic rings to PE in the presence of l-NAME was consistently greater for all groups. Interestingly, the inhibition of NOS obviated the beneficial effect of the arginine–silicate treatment, suggesting an endothelium-dependent mode of action. The relaxant response of aortic rings (pre-contracted with EC80 concentration of PE) to ACh and SNP is shown in Fig. 2. Corresponding values of the logistic equation parameters are summarised in Table 2. The presence of l-NAME reduced the relaxant response to ACh in all groups to 0–3% (data not shown). The EC50 of the response to ACh was significantly lower in rings from +/? rats than in rings from cp/cp control animals (p<0.05), which is consistent with previous reports [6]. Furthermore, aortic rings from both arginine–silicate-treated and arginine–HCl-treated groups had significantly lower EC50 values than cp/cp controls (Table 2). In contrast, there were no statistically significant differences in the relaxant responses to SNP in any of the groups (Fig. 2, right panel, and Table 2).

Contractile response to PE of aortic rings from JCR:LA-cp rats in the absence (a) and presence (b) of the NOS inhibitor l-NAME (10−4mol/l). Open circles, +/? control; filled circles, cp/cp control; open squares, cp/cp treated with arginine–silicate; open diamonds, cp/cp treated with arginine–HCl. Values are means±SEM, 8–10 rats per group. The lines are fitted logistic curves, with the values of the coefficients shown in Table 1

Dose-response curves for the relaxation of PE pre-contracted aortic rings to the NO-releasing agent ACh (a) and the NO donor SNP (b). Open circles, +/? control; filled circles, cp/cp control; open squares, cp/cp treated with arginine–silicate; open diamonds, cp/cp treated with arginine–HCl. The values are means±SEM, 8–10 rats per group. The lines are fitted logistic curves, with values of the coefficients shown in Table 2
Coronary artery function
Coronary artery function was characterised in the Langendorf heart preparation using the variables of baseline coronary flow, reactive hyperaemia following 60 s of ischaemia, and the flow dose-response to bradykinin. Table 3 summarises the results. There was no difference in baseline coronary flow between groups, confirming the viability of hearts from all groups under the experimental protocol. Reactive hyperaemia was significantly lower in hearts from cp/cp control rats compared with those from +/? rats, consistent with coronary artery dysfunction [24]. Treatment of the cp/cp rats with both arginine preparations significantly improved the reactive hyperaemia (p<0.05 and p<0.01 for the silicate complex and HCl, respectively), with the values being comparable with those of the +/? hearts. Similarly, the relative coronary blood flow (flow/baseline flow) in response to bradykinin of hearts from cp/cp rats that received arginine–silicate treatment (in both maximum flow and EC50) was normalised to that of hearts from +/? animals (p<0.001, Table 3).
Renal effects
The concentration of albumin in the urine of cp/cp control rats was more than 5-fold greater than that of +/? rats (74.7±17.6 vs 13.5±2.0 mg/l, p<0.005, respectively), a finding which is typically associated with increased incidence of kidney damage. There was no difference in the urinary albumin concentrations between arginine–silicate-complex-treated, arginine–HCl-treated cp/cp groups and controls (83.0±15.8 and 71.5±16.9 mg/l, respectively). Similar results were found for the albumin : creatinine ratio (data not shown). Interestingly, however, histological analysis of glomeruli of the cp/cp rats demonstrated a strikingly higher frequency of sclerosis than in glomeruli of the +/? rats (30.5±5.5 vs 1.3±0.8%, p<0.001). Notably, arginine–silicate-treated rats showed less than half the incidence of sclerotic damage to the glomeruli, a finding which was not observed in the arginine–HCl-treated rats (9.3±3.1%, p<0.05 and 24.8±5.2%, respectively). Figure 3 shows representative micrographs of normal glomeruli of +/? rats and abnormal glomeruli of cp/cp rats, together with glomeruli of arginine–silicate-treated cp/cp rats.

Representative renal histology of JCR:LA-cp rats at 12 weeks of age. a +/? control rat showing normal glomerular structure. b cp/cp control rat showing one moderately and two severely sclerotic glomeruli. c, d Arginine–silicate-treated cp/cp rats showing completely normal glomerular morphology. H&E stain
Discussion
The rationale for this study was that the efficacy of arginine supplementation could be enhanced with a novel complex of arginine, silicate and inositol through increased absorption and bioavailability in vivo. Increases in arginine bioavailability, and subsequent increases in NO production, could potentially offer the means to attenuate vascular dysfunction and the pathophysiology associated with the early stages of the metabolic syndrome and type 2 diabetes. The dosage chosen is in a reasonable range for a physiological, as opposed to a pharmacological, dietary supplementation in the rat, with its high relative food intake and rapid metabolism compared with humans.
Arginine administration had minimal effects on food intake and body weight with an unexpected increase in insulin concentrations. However, arginine is a powerful insulin secretagogue, especially in the cp/cp rat [25], and this may underlie the increase in fasting insulin levels observed in the highly hyperinsulinaemic rats. Increased plasma insulin levels were not accompanied by any change in the plasma glucose concentrations, which under normal conditions are highly regulated and stable in the cp/cp rat (albeit at the expense of high circulating insulin concentrations) [9]. Reductions in body weight and insulin concentrations have been shown to correspond to the reduction in cardiovascular complications, including vascular dysfunction in the cp/cp rat [26]. Given this, it is intriguing that the arginine–silicate–inositol complex reduced vascular complications in the JCR:LA-cp rat in the presence of increased plasma insulin levels. Thus, elevated plasma insulin found in arginine-treated rats appears to be a predictable response to the increased concentration of an insulin secretagogue; however, beneficial effects on the vessel wall seem to have counterbalanced the possible deleterious effects. The findings from this study confirm that the mechanisms underlying the vascular damage and vasculopathy associated with the metabolic syndrome are complex, and may involve multiple pathways. Perhaps more importantly, these observations suggest that beneficial actions of arginine (when prepared as the silicate–inositol complex), occur independently of protective effects that are usually attributed to reduction of hyperinsulinaemia.
Hypercontractility to noradrenergic stimulus, both hormonal and neurogenic, is an important mediator of end-stage vascular disease, leading to vasospasm, resultant ischaemia and infarct. Sudden death from such events is a common terminal event in cardiovascular disease. In the cp/cp rat, arterial dysfunction involves both the endothelium and the vascular smooth muscle cells [27–29]. The reduction in maximal contractile response to the noradrenergic agonist PE in the arginine–silicate-treated rats is clearly due to improved endothelial function as indicated by the EC50 for ACh, normalised maximal relaxation and obviation of the effect on the PE response by l-NAME. The absence of a significant reduction in contractility in the arginine–HCl-treated rats is consistent with the lower plasma concentrations of arginine achieved in this group. A reduced EC50 for ACh was found with both arginine compounds, indicating enhanced sensitivity of the endothelial cells to ACh. We conclude, from these findings, that there is potential improvement in intracellular signalling and increased NO release leading to beneficial effects on vascular function. The JCR:LA-cp rats, both obese and lean, are normotensive [30] and thus blood pressure is unlikely to have been reduced by the arginine treatment. However, the reduced sensitivity to noradrenergic stimuli might reduce vascular tone and propensity to vasospasm in vivo.
The improvements in coronary artery function observed in this study are functionally significant, especially given the morphological and physiological differences between the smaller and more vulnerable vessels of the coronary circulation and the aorta. The coronary circulation in cp/cp rats showed an improved relaxant response to short hypoxia after treatment with arginine–silicate, as well as improved response to the endogenous peptide, bradykinin. The increased sensitivity to bradykinin is also physiologically relevant as bradykinin is protective against myocardial ischaemia [31] and plays a role in the cardioprotective action of ACE inhibitors in the cp/cp rat [32].
The cp/cp rats show a 5-fold increase in urinary albumin at 12 weeks of age, compared with +/? animals. Albuminuria reflects glomerular microvascular damage, with increased permeability, and has been proposed to be an early marker for cardiovascular disease in type 1 diabetes [33]. Neither the arginine–silicate nor arginine–HCl treatment used in this study had any effect on the albumin levels in JCR:LA-cp animals. Nonetheless, the arginine–silicate-complex-treated rats showed a significant reduction in the degree of glomerular sclerosis, similar to that seen with the neutral endopeptidase inhibitor AVE7688, an agent that also improves vascular function in the cp/cp rat [22, 34]. It could be inferred, from this observation, that increased glomerular permeability or other renal dysfunction that lead to albuminuria and to damage to the glomerular capillary structure with subsequent glomerular sclerosis may involve distinct and independent pathways. The results are consistent with two different modifiable mechanisms, potentially operating at the cellular level, both having an endpoint that leads to vascular damage and dysfunction. We speculate that enhanced NO metabolism and release (associated with increased plasma arginine concentrations) may counteract the effects of endothelial contractile dysfunction, even in the presence of continuing hyperinsulinaemia. Furthermore, these pharmacological actions are consistent with the fact that glomerular sclerosis may be attributed to capillary or arteriolar vasospasm and ischaemia, a process that is known to occur at the arterial level in the heart of the cp/cp rat [13]. Our results are also consistent with those of Albrecht et al. [35], who reported that chronic treatment with approximately 1.5 g kg−1 day−1 of arginine (chemical form unspecified) showed significant reduction in transplant-associated glomerular sclerosis in rats. These authors demonstrated a significant reduction in the expression of interstitial alpha smooth muscle actin in l-arginine-treated animals, which in turn is thought to be partly responsible for the increased interstitial fibrosis associated with sclerotic damage [36]. Similarly, Vos et al. [37], using an allograft model of kidney damage, also reported that l-arginine supplementation can improve the glomerular filtration rate and minimise glomerular oedema. Moreover, Vos et al. have shown that glomerulosclerosis-associated inflammation can be driven by the nuclear factor kappa B pathway, which is typically inhibited by NO [38]. Consequently, there seems to be a merging and attractive consensus that beneficial effects of arginine may be due (at least in part) to the anti-inflammatory properties of increasing local NO availability at the site of the Bowman’s capsule [38, 39].
In this study, the beneficial effect of arginine supplementation in the absence of any reduction in plasma insulin concentrations was unexpected, and unique in our experience with this model. We have reported previously that the insulin resistance of the cp/cp rat is accompanied by enhanced serine kinase activity and attenuated IRS-1 phosphorylation and phosphatidylinositol-3 kinase activity following insulin stimulation [40, 41]. The abnormal signal transduction may well extend to other pathways that are related and that can be upregulated by arginine or NO through as yet unknown mechanisms.
Collectively, the results show that the arginine–silicate–inositol complex is absorbed effectively, without any evidence of toxicity in experimental rats. There were significant beneficial effects on vascular function and glomerular status, consistent with protection against end-stage effects of macro- and microvascular disease of the metabolic syndrome. The effects occurred in the absence of any reduction in hyperinsulinaemia, suggesting direct beneficial effects on endothelial cell function and NO metabolism. We further speculate that arginine–silicate complex may confer an additional reduction in vascular complications, particularly if combined with effective insulin-sensitising compounds. While the results of studies in non-human species are not always confirmed in humans, the rat is a close analogue of humans in many respects and has proven to be an effective model for pharmaceutical and nutrition research. The JCR:LA-cp strain mimics the metabolic and pathophysiological aspects of the metabolic syndrome very closely. As far as we know, our findings are unique with respect to the physiological significance of arginine supplementation and have potential implications for the prevention and clinical treatment of cardiovascular and renal disease associated with the metabolic syndrome and type 2 diabetes.
Abbreviations
- ACh:
-
acetylcholine
- H&E:
-
haematoxylin and eosin
- l-NAME:
-
N G-nitro-l-arginine methyl ester
- NO:
-
nitric oxide
- NOS:
-
nitric oxide synthase
- PE:
-
phenylephrine
- SNP:
-
sodium nitroprusside
References
Steiner G (1994) Hyperinsulinemia and hypertriglyceridemia. J Int Med 736(Suppl):23–26
Després J-P, Lamarche B, Mauriège P et al (1996) Hyperinsulinemia as an independent risk factor for ischemic heart disease. N Engl J Med 334:952–957
Uusitupa MI, Niskanen LK, Siitonen O, Voutilainen E, Pyorala K (1990) 5-year incidence of atherosclerotic vascular disease in relation to general risk factors, insulin level, and abnormalities in lipoprotein composition in non-insulin-dependent diabetic and nondiabetic subjects. Circulation 82:27–36
O’Brien SF, Russell JC (1997) Insulin resistance and vascular wall function: lessons from animal models (Review). Endocrinol Metab 4:155–162
Woodman RJ, Chew GT, Watts GF (2005) Mechanisms, significance and treatment of vascular dysfunction in type 2 diabetes mellitus: focus on lipid-regulating therapy. Drugs 65:31–74
Russell JC, Dolphin PJ, Graham SE, Amy RM, Brindley DN (1998) Improvement of insulin sensitivity and cardiovascular outcomes in the JCR:LA-cp rat by d-fenfluramine. Diabetologia 41:380–389
Russell JC, Ravel D, Pégorier J-P et al (2000) Beneficial insulin-sensitizing and vascular effects of S15261 in the insulin-resistant JCR:LA-cp rat. J Pharmacol Exp Ther 295:753–760
Krentz AJ, Bailey CJ (2005) Oral antidiabetic agents: current role in type 2 diabetes mellitus. Drugs 65:385–411
Russell JC, Graham SE, Dolphin PJ (1999) Glucose tolerance and insulin resistance in the JCR:LA-cp rat: effect of miglitol (Bay m1099). Metabolism 48:701–706
Russell JC, Bar-Tana J, Shillabeer G et al (1998) Development of insulin resistance in the JCR:LA-cp rat: role of triacylglycerols and effects of MEDICA 16. Diabetes 47:770–778
Vance JE, Russell JC (1990) Hypersecretion of VLDL, but not HDL, by hepatocytes from the JCR:LA-corpulent rat. J Lipid Res 31:1491–1501
Elam MB, Wilcox HG, Cagen LM et al (2001) Increased hepatic VLDL secretion, lipogenesis, and SREBP-1 expression in the corpulent JCR:LA-cp rat. J Lipid Res 42:2039–2048
Russell JC, Graham SE, Richardson M (1998) Cardiovascular disease in the JCR:LA-cp rat. Mol Cell Biochem 188:113–126
O’Brien SF, Russell JC, Davidge ST (1999) Vascular wall dysfunction in JCR:LA-cp rats: effects of age and insulin resistance. Am J Physiol 277:C987–C993
Russell JC, Amy RM (1986) Myocardial and vascular lesions in the LA/N-corpulent rat. Can J Physiol Pharmacol 64:1272–1280
Russell JC, Amy RM, Graham SE, Dolphin PJ, Wood GO, Bar-Tana J (1995) Inhibition of atherosclerosis and myocardial lesions in the JCR:LA-cp rat by β, β′-tetramethylhexadecanedioic acid (MEDICA 16). Arterioscler Thromb Vasc Biol 15:918–923
Radomski MW, Salas E (1995) Nitric oxide-biological mediator, modulator and factor of injury: its role in the pathogenesis of atherosclerosis. Atherosclerosis 118:S69–S80
Juturu V, Komorowski JR, Roa KS (2004) Arginine silicate inositol complex does not induce mutation in the AMES Bacterial: reverse mutation test. FASEB J 18:A869 (Abstract)
Juturu V, Komorowski JR, Gudi R (2004) Orally administered arginine silicate inositol complex is not clastogenic in Chinese hamster ovary cells. FASEB J 18:A869 (Abstract)
Komorowski JR, Juturu V, Gudi R (2004) Arginine silicate inositol complex is not toxic in mouse micronucleus assay. FASEB J 18:A869 (Abstract)
Russell JC, McKendrick JD, Dubé GP, Dolphin PJ, Radomski MW (2001) Effects of LY117018 and the estrogen analogue, 17α-ethinylestradiol, on vascular reactivity, platelet aggregation, and lipid metabolism in the insulin-resistant JCR:LA-cp rat: role of nitric oxide. J Cardiovasc Pharmacol 37:119–128
Schäfer S, Steioff K, Linz W, Bleich M, Busch AE, Löhn M (2004) Chronic vasopeptidase inhibition normalizes diabetic endothelial dysfunction. Eur J Pharmacol 484:361–362
De Lean A, Munson PJ, Rodbard D (1978) Simultaneous analysis of families of sigmoidal curves: application to bioassay, radioligand assay, and physiological dose-response curves. Am J Physiol 235:E97–E102
Masayoshi H, Yoshitomo M, Yasuaki M, Hozuka A (2003) New methods to evaluate endothelial function: non-invasive method of evaluating endothelial function in humans. J Pharmacol Sci 93:405–408
Pederson RA, Campos RV, Buchan AMJ, Chisholm CB, Russell JC, Brown JC (1991) Comparison of the enteroinsular axis in two strains of obese rats: the fatty Zucker and the JCR:LA-corpulent. Int J Obes 15:461–470
Richardson M, Schmidt AM, Graham SE, Achen B, DeReske M, Russell JC (1998) Vasculopathy and insulin resistance in the JCR:LA-cp rat. Atherosclerosis 138:135–146
Absher PM, Schneider DJ, Russell JC, Sobel BE (1997) Increased proliferation of explanted vascular smooth muscle cells: a marker presaging atherogenesis. Atherosclerosis 131:187–194
Absher PM, Schneider DJ, Baldor LC, Russell JC, Sobel BE (1999) The retardation of vasculopathy induced by attenuation of insulin resistance in the corpulent JCR:LA-cp rat is reflected by decreased vascular smooth muscle cell proliferation in vivo. Atherosclerosis 143:245–251
Brunner F, Wölkert G, Russell JC, Wascher T (2000) Vascular dysfunction and myocardial contractility in the JCR:LA-corpulent rat. Cardiovasc Res 36:150–158
Russell JC, Amy RM (1986) Plasma lipids and other factors in the LA/N-corpulent rat in the presence of chronic exercise and food restriction. Can J Physiol Pharmacol 64:750–756
Linz W, Martorana PA, Schölkens BA (1990) Local inhibition of bradykinin degradation in ischemic hearts. J Cardiovasc Pharmacol 15:S99–S109
Russell JC, Graham SE, Amy RM, Dolphin PJ (1998) Inhibition of myocardial lesions in the JCR:LA-corpulent rat by captopril. J Cardiovasc Pharmacol 31:971–977
Dogra G, Rich L, Stanton K, Watts GF (2001) Endothelium-dependent and independent vasodilation studies at normoglycaemia in type I diabetes mellitus with and without microalbuminuria. Diabetologia 44:593–601
Russell JC, Kelly SE, Schäfer S (2004) Vasopeptidase inhibition improves insulin sensitivity and endothelial function in the JCR:LA-cp rat. J Cardiovasc Pharmacol 44:258–265
Albrecht EWJA, van Goora H, Smit-van Oostena A, Stegeman CA (2003) Long-term dietary-arginine supplementation attenuates proteinuria and focal glomerulosclerosis in experimental chronic renal transplant failure. Nitric Oxide 8:53–58
Maski T, Stambe C, Hill PA, Dowling J, Atkins RC, Nikolic-Paterson DJ (2004) Activation of the extracellular-signal regulated protein kinase pathway in human glomerulopathies. J Am Soc Nephrol 15:1835–1843
Vos IHC, Rabelink TJ, Dorland B et al (2001) l-Arginine supplementation improves function and reduces inflammation in renal allografts. J Am Soc Nephrol 12:361–367
Peng H-B, Rajavashisth TB, Libby P, Liao JK (1995) Nitric oxide inhibits macrophage-colony stimulating factor gene transcription in vascular endothelial cells. J Biol Chem 270:17050–17055
De Caterina R, Libby P, Peng H-B et al (1995) Nitric oxide decreases cytokine-induced endothelial activation. J Clin Invest 96:60–68
Qiao L-Y, Goldberg JL, Russell JC, Sun XJ (1999) Identification of enhanced serine kinase activity in insulin resistance. J Biol Chem 274:10625–10632
Cefalu WT, Wang ZQ, Zhang XH, Baldor LC, Russell JC (2002) Oral chromium picolinate improves carbohydrate and lipid metabolism and enhances skeletal muscle Glut-4 translocation in obese, hyperinsulinemic (JCR-LA corpulent) rats. J Nutr 132:1107–1114
Acknowledgements
This study was supported financially by Nutrition 21 Inc., Purchase, NY, USA. We thank Kristina MacNaughton for excellent technical assistance. S. D. Proctor was supported by NHMRC of Australia Grant #229030. This work was presented, in part, at the 2004 Arteriosclerosis, Thrombosis and Vascular Biology Scientific Meeting in San Francisco.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Proctor, S.D., Kelly, S.E. & Russell, J.C. A novel complex of arginine–silicate improves micro- and macrovascular function and inhibits glomerular sclerosis in insulin-resistant JCR:LA-cp rats. Diabetologia 48, 1925–1932 (2005). https://doi.org/10.1007/s00125-005-1862-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00125-005-1862-8