
Abstract
The European bison is the largest terrestrial mammal in Europe. After extinction in the wild at the beginning of the twentieth century, it was re-introduced to Białowieża Forest and other woodlands in Eastern Europe. In this paper, we analysed the movements of European bison beyond the continuous woodlands of the Białowieża Forest (NE Poland) between 1964 and 2010. In total, 1,117 direct observations of bison were collected. The number of males moving out of the forest fluctuated during the study period, whilst the number of females steadily increased. The number of male observations outside of the forest per annum was dependent on the population size and snow depth, whilst the number of cows in mixed groups moving outside of the forest was correlated with the population size only. Males were observed mainly alone (50 % of observations) or in small groups of two to three individuals (25 %); however, distribution of group size differed from those observed in the population. There was a significant difference between the direction of movement of males and females out of the forest—males moved mainly west and southwest, whilst females moved to the north. This was also significantly different from the expected movement direction. The mean distance of bison observations from the forest border was 1.8 ± 0.13 km and did not differ significantly between sexes. After 1990, males were observed significantly farther away from the forest (2.2 km) than in the previous years (0.9 km). Most observations (94 % of bulls and 93 % of cows) were up to 5 km from the forest edge. The range of bison in the vicinity of the Białowieża Forest was strictly seasonal. Most observations (78 % in males and 88 % in females) were recorded from November to April. Increasing utilisation of areas beyond the forest habitats may be driven by different factors but most probably it is related to range expansion and the bison's preference for open habitats. The strong seasonal pattern of bison movements indicate that the partial seasonal migrations were initiated in the Białowieża population.
Similar content being viewed by others


Introduction
Recolonisation by native species into its historical range is, in general, realised on the way of pulsed expansion (Caughley 1970; Larter et al. 2000). This process is usually density-dependent and driven by intraspecific competition for food as increases in local areas are followed by pulses of dispersal leading to range expansions (Larter et al. 2000). As the areas available for wild animals, especially large ungulates, are usually limited, range expansion often leads to human–wildlife conflicts (Plumb et al. 2009; Bleier et al. 2012; Hofman-Kamińska and Kowalczyk 2012).
European bison (Bison bonasus), the largest terrestrial mammal in Europe, is being managed as a forest specialist. However, it has been proposed that it is an example of a refugee species that, due to combination of increasing replacement of open steppe by forest cover after the last postglacial period and increasing human pressure, was forced into forests as a refuge habitat (Cromsigt et al. 2012; Kerley et al. 2012). The last natural population of the lowland European bison survived in the Białowieża Forest until the beginning of the twentieth century. Warfare and poaching caused a drastic population decline and finally, the extinction of the population in 1919 (Sztolcman 1926). Wild-living populations of bison were restored using captive animals and released into the wild at first in the Polish part of the Białowieża Forest in 1952 (Krasiński 1978) and then into forest habitats in Belarus, Lithuania, Russia, Slovakia and Ukraine. The Białowieża Forest currently has the largest bison population numbering over 900 bison (480 bison in the Polish part and 430 in the Belarussian part) (Raczyński 2011) and has been separated since 1981 by the border fence between Poland and Belarus (Kowalczyk et al. 2012). In winter, bison are supplementarily fed with hay at permanent feeding sites in order to maintain their body condition, reduce potential damages caused by bison to the tree stands and farm crops and reduce the level of migration out of the forest (Krasiński 1978; Krasińska and Krasiński 2007; Kozlo and Bunevich 2009). Furthermore, the bison population in the Białowieża Forest has been culled regularly since the 1970s to reduce growth of the population and remove invalid animals (Krasińska and Krasiński 2007).
Since being introduced in the central part of Polish side of the Białowieża Forest, the bison population has successively increased its range until the population size exceeded 100 individuals in 1965. During the next 20 years, in spite of the growth in the population size, mixed groups and the majority of bulls occupied a stable range of about 140 km2 (22 % of the area of the Białowieża Forest) (Krasiński 1978; Krasińska and Krasiński 2007). At the beginning of 1990s, the population dispersed spontaneously throughout the entire forest area (Krasińska and Krasiński 2007). Similar spatial behaviours were observed in the population in the Belarussian part of the Białowieża Forest during the first 30 years after re-introduction; however, in the 1980s, nearly 40 individuals were captured and translocated to other parts of the forest (Bunevich 2003). Range expansion to include open habitats in their range is observed in 69 % of European bison populations (Kerley et al. 2012), but this utilisation of the open habitats is regarded as a management problem because it leads to use of agricultural lands and crop depredation (Kerley et al. 2012; Hofman-Kamińska and Kowalczyk 2012). There is no historical data on seasonal migrations of lowland European bison; however, there are few mentions of the distinct movements of a few solitary individuals (Sztolcman 1926; Daszkiewicz and Samojlik 2009). Seasonal migrations of European bison are observed in the forest–field landscape in Poland (the Knyszyn Forest and West Pomeranian forests) (Krasiński et al. 1994; Bereszyński 2005; Krasińska and Krasiński 2007) and also, altitudinal seasonal migration related to weather conditions and snow cover are observed in mountainous areas (Paszkiewicz 2004; Perzanowski and Januszczak 2004; Nemtsev et al. 2003). In the closely related American plains bison (Bison bison bison), seasonal migrations were observed historically (Garretson 1938; Roe 1970) and also in more recent times (McHugh 1958; Meagher 1973, 1998; Shaw and Carter 1990). Populations of American plains bison and wood bison (Bison bison athabascae) also show range expansion that is often initiated by males (Gates and Larter 1990; Larter and Gates 1990). Dispersal of American bison populations is usually related to the decline in nutritional resources as an effect of population growth (Larter 1988; Meagher 1973; Gates and Larter 1990; Larter and Gates 1990; Plumb et al. 2009).
In this paper, we aimed to analyse long-term data on the movements of European bison out of the woodlands of the Polish part of the Białowieża Forest. We investigated: (1) spatial and seasonal distribution of movements out of the forest, (2) differences in movement pattern between males and females and (3) factors driving those movements. We hypothesised that the range expansion of the population towards open habitats resulted in access of bison to attractive winter foraging grounds and, in turn, partial seasonal migrations of bison were initiated. Knowledge on factors responsible and the course of the range expansion is essential for conservation management of this species, especially in mitigating human–bison conflict whilst allowing the status of the species to improve. It will also aid in the development of guidelines for ecological corridor planning, new re-introduction projects and further dispersal of European bison populations.
Study area
The study was conducted in the Białowieża Forest (northeastern Poland), located on the Polish-Belarussian border (52°30′–52°54′ N, 23°26′–23°56′ E), in one of the best preserved lowland forests in Europe. Deciduous and mixed tree stands constitute 94 % of the Białowieża Forest, whilst open habitats (glades with meadows, riverside open sedge and reed marshes) cover only 6 % of the forest (Sokołowski 2004). To the west, southwest and north, the forest is surrounded by open habitats dominated by pastures and meadows (24 % of open area) and arable lands (48 %) interrupted with small woodlands (25 %). The surroundings of the forest are characterised by extensive agriculture with small farms and low human density (30 people/km2; Demographic Yearbook of Poland 2009; Statistical Yearbook of Agriculture 2009). The main cultivated crops are cereals, mainly rye (51 % of arable lands) and potatoes (5 %). In winter, 25 % of the arable lands are covered by winter cultivations of cereals and rape (Statistical Yearbook of Agriculture 2009). The forest is inhabited by over 900 bison (480 in Polish and 430 in Belarussian part of the Forest), separated since 1981 by a border fence (Bunevich 2004; Krasińska and Krasiński 2007; Kowalczyk et al. 2012).
The climate of the Białowieża Forest is transitional between Atlantic and continental types. Mean annual temperature is 7 °C. The warmest month is July (average 18.4 °C) and the coldest is January (average −4.8° C). Average growing season is 200–210 days. Snow cover lasts from 60 to 96 days, with maximum recorded depth of 95 cm. Annual precipitation averages 610 mm (Jędrzejewska and Jędrzejewski 1998).
Material and methods
The European bison population in Białowieża Forest has been carefully monitored since it's re-introduction into the wild. This monitoring includes regular winter surveys, which record the number of bison, sex and age structure as well as regular monitoring of bison distribution (done by radio-tracking over the last 17 years) and responding to bison–human conflicts. Observational data on bison outside of the Białowieża Forest analysed in this paper included: (1) occasional direct observations of bison reported by farmers and verified by National Park rangers; (2) locations of collared individuals in years 2005–2010; (3) records based on ground and snow tracking; (4) locations of dead, captured or culled individuals and (5) locations of bison depredation of farm crops in the vicinity of the Białowieża Forest (2000–2008), provided by the Regional Directorate for Environmental Protection in Białystok. In total, 1,117 observations of European bison outside the area of the Białowieża Forest were collected and used in the analysis. Of 42 bison collared between 2005 and 2010, 10 individuals (five males and five females) fitted with VHF collars were seasonally observed outside of the forest. Locations of radio-collared bison constituted 29 % of analysed observations in case of males, and 22 % in case of females. Data were analysed separately for bulls (n = 729) and mixed groups (cows with calves and sub-adults) (n = 388). Data on cows was from 1992 (first observations of mixed groups out of the forest) to 2010, whilst the data concerning bulls was divided into two periods. The first period is from 1964 to 1989 (n = 167), when the main part of the population (mixed groups and the majority of bulls) used the central part of the forest, had even calf sex ratio and exhibited a slow increase (Hayward et al. 2011). In the second period from 1990 to 2010 (n = 562), the population began to rapidly increase that led to increased dispersal rates, decrease in female body condition and female-biased calf sex ratio (Hayward et al. 2011).
The number of bison observed outside of the forest in various years was estimated as the maximum number of individuals roaming in open areas. The data was recorded during a short time period in different locations. The approximate age of males (ranging from 3 to 18 years) based on horn size and shape and body characteristics (Krasińska and Krasiński 2007) was determined for 62 % of observed males (n = 455).
We used logistic regression (generalised linear model with Poisson distribution and logit link function) to determine influence of different factors on number of European bison observed annually out of the Białowieża Forest. The factors analysed were population size, sex ratio, snow depth, annual temperature and precipitation. Data on population size and bison sex ratio originated from the annual survey conducted in winter by Białowieża National Park staff (Krasińska and Krasiński 2007). For more details, see Hayward et al. (2011). The meteorological data originated from the weather stations of Białowieża National Park and Mammal Research Institute.
Distances of bison movements were determined as the shortest straight line distances between observed individuals or herds and the nearest forest edge using ESRI ArcGIS 9.3 software. The data were analysed in the periods: spring (April–June), summer (July–September), autumn (October–November) and winter (December–March). Statistical analyses were performed using STATISTICA (Stat Soft 1984–2000).
Results
The first distinct movements of males out of the Białowieża Forest area were recorded in 1964, i.e., 12 years after the re-introduction of bison, when the population numbered 84 individuals (Fig. 1). Over 46 years, there were, on average, nine males a year (0–33 individuals) observed outside the forest. The highest number of males beyond the forest was observed from 1984 to 1989 (Fig. 3). Logistic regression analysis performed for the whole study period showed that the number of males observed out of the forests was dependent on the population size and snow depth. (Table 1, Fig. 4).

Observations of European bison males out of the Polish part of the Białowieża Forest
Mixed groups (cows with calves and sub-adults) have regularly moved out of the forest since 1994 (Fig. 2). On average, 44 cows (range: 26–106) roaming in mixed groups were observed annually on agricultural lands. Once the population had reached 260 individuals (1.8 individuals/km2; Figs. 3 and 4), the number of cows observed out of the forest systematically increased (from 26 individuals in 1994 to 106 in 2010). The number of bison in mixed groups observed out of the forest was related only to population size (Table 1, Fig. 4). Proportion of the bison that moved out of the forest varied from 0 to 27 % of the population annually and gradually increased during the study (Fig. 3).

Observations of European bison females out of the Polish part of the Białowieża Forest

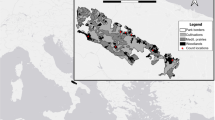
Number of European bison males and females ranging out of the Polish part of the Białowieża Forest (1964–2010)

Relationship between the number of males (closed circles and solid line) and females (open circles and broken line) observed out of the Białowieża Forest (both log transformed) and the population size
Open areas were most frequently visited by single bulls (50 % of observations) or male groups of two to three individuals (25 %). Only rarely were larger groups of six to 12 individuals recorded (Fig. 5). Observed group size distribution of bison outside of the forest significantly differed from those observed in the population inside of the forest (χ2 = 48.937, df = 4, p < 0.0001). Fewer solitary males and those in small groups, and more in larger groups roamed out of the forest in comparison to the population inside the forest (Fig. 5). Most of the observed males were 5–11 years old (54 % of males with known age) or older (42 %). Young males (3–4 years old) were only occasionally recorded (4 %) and they were always in groups with older bulls.

Group size distribution of males observed out of the Białowieża Forest (black bars) in comparison to those observed inside the forest (grey bars). (Krasińska and Krasiński 1995)
Movements of bison beyond the forest were seasonal. The majority of bison observations outside of the forest (78 % for males and 88 % for females) were recorded from November to April (Fig. 6). There were significant differences in the proportion of observations out of the forest in growing season between males in 1964–1989 and 1990–2010 (χ2 = 25.092, df = 11, p = 0.009) and between males in 1990–2010 and females (χ2 = 29.391, df = 11, p = 0.002). In the growing season (May–October), males in 1990–2010 were more often observed out of the forest (25 % of observations) than females and males in the previous period (12 % and 15 %, respectively).

Proportion of male and female observations out of the Białowieża Forest in different months
The average distance of bull observations from the forest edge was 1.8 ± 0.17 km (Table 2). Between 1964 and 1989, the distance was significantly smaller (mean 0.9 km) than in the years 1990–2010 (mean 2.2 km) (Table 2). In the first period, the maximum distance from the forest edge was 13.8 km and in the second, it was 33.9 km. The average distance of cow observations from the forest boundaries was 2.1 ± 0.18 km, which was not significantly different from that of bulls (Table 2). Distances of males and cows from the forest in the same season did not differ significantly (ANOVA and post hoc Tukey test, p > 0.1). Bison were recorded the farthest from the forest in summer (2.75 ± 0.6510.2 km and 5.9 ± 6.7 km, respectively) (Fig. 7). Summer distances differed from those in other seasons (ANOVA and post hoc Tukey test, p < 0.0001). When long-distance migrations of bison were excluded from the analysis, seasonal differences were no longer significant (ANOVA and post hoc Tukey test p > 0.5) (Fig. 7).

Average distances (mean ± SE) of male and female observations from the edge of the forest in different seasons in the Białowieża Forest
Both male and female bison that moved outside of the forest were most often observed at between 1 and 5 km away from the forest boundary (50 % of relocations in bulls and 59 % in cows). Whilst bulls moved to 1 km from the forest more frequently than cows did (44 % and 34 %, respectively), this difference was not statistically significant (χ2 = 4.514, p = 0.2). European bison of both sexes rarely roamed further than 5 km (6–7 %) from the forest edge (Table 3). There was a significant difference between the direction of movement of males and females out of the forest, and this also significantly differed from the expected direction of movement (Table 4). Males preferred to move to the west (78 % of observations, Jacobs index D = 0.68) until 1989 and southwest (49 % of records, D = 0.69) in following years, whilst females strongly preferred the north direction (81 % of observations, D = 0.68) (Table 4, Figs. 1 and 2).
Discussion
European bison in the Białowieża Forest showed increasing utilisation of open areas outside of the forest. This was probably caused by range expansion of the bison population that led to greater access to attractive foraging areas, especially in winter time. Range expansion to include open areas is observed in 70 % of free-ranging bison populations (Kerley et al. 2012). In Białowieża Forest, bison introduced to the centre of the forest were supplementarily fed in winter to mitigate migrations and limit expansion to neighbouring open areas. Increasing density initiated the pulsed expansion, which is typical for a species recolonising its historical range (Reynolds 1998; Larter et al. 2000). The range expansion of the population and the increasing number of bison observed out of the forest was density dependent as it was influenced by population size. Patterns of density dependence in dispersal rates may differ markedly between sexes in highly polygynous species. Females are usually food limited, whilst males are limited by access to mates. When local density increases, forage availability declines, whilst mate access for males may increase due to an increasingly female-biased sex ratio (Loe et al. 2009). We have not found evidence on the influence of sex ratio on the number of bison moving out of the forest; however, until the late 1980s, the sex ratio of bison calves was male biased (Hayward et al. 2011). This indicates stronger mate competition between males that might force their dispersal (Handley and Perrin 2007; Pérez-González and Carranza 2009). In the closely related American wood bison (B. bison athabascae), about 40 % of mature males were reproductively successful (Wilson et al. 2002). This suggests that bison with a lower position in the dominance hierarchy (usually younger sexually mature males) may have limited access to mating and may increase their movements to search for mates. We observed higher proportion of males moving out of the forest in groups in comparison to the population inside the forest. As group size is negatively related to age, this indicates that younger male bison are moving out of the forest (Krasińska and Krasiński 1995). Increased movements of males in the 1980s, in comparison to other decades, may also have been exploratory in nature and may indicate preference to open habitats as predicted by refugee species concept (Kerley et al. 2012). It was additionally driven by an increase in the area of rape cultivation on the fields west of the Białowieża Forest, which has created attractive winter foraging area for bison (Hofman-Kamińska and Kowalczyk 2012). Those factors probably caused marked increases in the number of extra-limital males observed out of the forest in the early 1980s. In adult American wood bison, single males or small male groups move to examine new habitat patches and then return to core breeding areas of females (Komers et al. 1992, 1993). Gates and Larter (1990) indicated that distant movements of American wood bison adult male may be a result of innate behaviour. Increasing density of bison in Białowieża initiated the pulse range expansion of mixed groups, as observed in early 1990s (Krasińska and Krasiński 2007), resulted in increasing utilisation of open habitats by females in the vicinity of the forest. It seems that the process was density-dependent and driven by intra-specific competition for food (Larter et al. 2000). If bison are forest specialists, the range expansion should lead to an even larger distribution of the population within the Białowieża Forest. However, some suitable parts of the forest are still not occupied or rarely utilised by bison and an increasing proportion of the population is observed in open areas adjacent to the forest. This may suggest withdrawal of bison from the refugee habitat (Kerley et al. 2012). If migration is adaptive, movement between seasonal ranges should involve a positive change in some aspect of the habitat (Ball et al. 2001). Forest habitats in the growing season offer relatively high biomass of easily digestible herbaceous plants and tree leaves (Jędrzejewska and Jędrzejewski 1998). In autumn, most of the vegetation on the forest floor is disappearing. To compensate for the declining plant biomass, bison may increase their browsing (Kowalczyk et al. 2011), but more often, they will aggregate at feeding sites or migrate more to agriculture areas where winter cultivations of rape and cereals or hay, which are stored in meadows by farmers, offer attractive foraging opportunities (Hofman-Kamińska and Kowalczyk 2012). Thus, the seasonal partial migrations to open habitats observed in Białowieża is an adaptation to seasonal change in food resources. Similar patterns are observed in other Polish populations in Knyszyn Forest (50 km north of Białowieża Forest) and in the West Pomeranian herd (western Poland), where bison migrate to agricultural areas in autumn and winter (Krasiński et al. 1994; Bereszyński 2005; Krasińska and Krasiński 2007; Hofman-Kamińska and Kowalczyk 2012).
Weather conditions may influence the rate of range expansion and seasonal migrations (Reynolds 1998; Ball et al. 2001). In our study, snow depth negatively influenced movements of male bison out of the forest. As showed by Potapov et al. (2011), snow depth may significantly reduce ungulate movements. During deep snow, ungulates prefer wooded habitats and avoid open fields (Potapov et al. 2011). This is probably related to snow depth, which is significantly reduced by canopy cover (D'Eon 2004). Radio-tracking of bison conducted in Białowieża Forest in previous years showed that long-lasting snow cover and low temperatures significantly limited mobility of bison in the forest (Krasińska et al. 2000). The high influence of snow cover on bison males is probably related to their larger ranges and greater mobility (Krasińska et al. 2000; Potapov et al. 2011).
Supplementary feeding is much less intensive on the edges of the forest than it is in the centre, but this did not stop the expansion of the bison to open areas that offer attractive foraging opportunities in winter (Hofman-Kamińska and Kowalczyk 2012). Increasing the selection of agriculture fields instead of winter feeding sites at fixed locations within the forest may also be a strategy to avoid negative effects of winter aggregation, such as risk of disease and parasite transmission (Wołk and Krasińska 2004; Radwan et al. 2010; Pyziel et al. 2011), and aggressive interactions and culling.
We found significant differences in seasonal movement pattern of male and females. Observations of females ranging out of the forest were more concentrated in the winter period. Differences in the observed pattern were probably due to differential sex-specific risks and benefits of foraging in human disturbed areas (Clutton-Brock et al. 1988; Lykkja et al. 2009). Males usually exhibit lower flight and weaker anti-predator responses than females and are rather indiscriminate with respect to human activity and land cover (Winnie and Creel 2007; Ramsrud 2007; Stankowich 2008). Females with calves in Białowieża utilised agricultural areas almost exclusively in winter, probably to avoid human activity and disturbance risk as these are at the lowest levels in this period. The same factors were probably responsible for the observed direction of bison movements out of the forest. Both males and females differed from each other and from expected in this respect. Females utilised open areas mainly north of the forest, which is dominated by extensively utilised (mowed usually once a year) meadows with very low density of human settlements and numerous woodland patches, areas of low disturbance risk and good cover. Behaviour of mixed groups utilising open areas out of the forest, mainly dawn–dusk activity far away from human settlements and hiding in woodland patches during the day, was adjusted to avoid contact with human. Males moved more randomly, utilising intensively managed farmland with attractive farm crops often visiting areas close to human settlements (such as orchards and gardens).
Northward and west–southward movements of bison in Białowieża result in low risk of conflict and are desired to some extent. Expansion to the north will allow for connection of Białowieża population with the neighbouring herd in Knyszyn Forest and create a compact distribution range with free migrations (Pucek et al. 2004). It may help to maintain genetic variability of the population, which is very low due to the bottleneck effect the bison experienced during their extermination in the wild and restoration from a small founder population (Tokarska et al. 2009, 2011; Wójcik et al. 2009). Grounds southwest of the Białowieża Forest offer a mosaic of woodlands and meadows and may serve as a suitable area for conflict-free population expansion.
The amount of farm crop depredation by bison is increasing (Hofman-Kamińska and Kowalczyk 2012). Although bison increase an area's attractiveness to tourists, increasing use of open areas by these large ungulates may very soon reach the level of social acceptance. We found that bison rarely moved further than 5 km from the forest edge. Creation of a buffer zone around woodlands that are inhabited by bison populations may be the way to secure winter foraging grounds for bison and reduce human–wildlife conflicts. In order to plan effective conservation strategies, future re-introduction programs should take into account the range expansion of increasing bison populations and the seasonal migrations to open habitats. Areas selected for re-introduction should include both forest and open habitats and be large enough to support viable bison population and any potential expansion. Knowledge on the spatial pattern and direction of European bison movements may guide adaptive management to reduce human–bison conflict and control expansion of the population in Białowieża Forest and other areas. Natural expansion of the bison population is a long-term process and may be driven by various factors such as population growth, presence and intensity of supplementary feeding and habitat structure (Reynolds 1998).
In conclusion, the range expansion of bison in the Białowieża Forest and access to open areas was probably driven by habitat preferences for grasslands/pastures. These patterns seem to be typical for many bison populations, which suggest that the European bison is not necessarily a forest specialist as previously thought, as indicated by Kerley et al. (2012). The strong seasonal pattern of these movements indicates that the partial seasonal migrations were initiated in Białowieża population. Range and spatial characteristics of these migrations need further investigation. We hope that intensive VHF and GPS tracking conducted in Białowieża population in previous years will allow investigation of this pattern in more detail.
References
Ball JP, Nordengren C, Wallin K (2001) Partial migration by large ungulates: characteristics of seasonal moose Alces alces ranges in northern Sweden. Wildl Biol 7:39–47
Bereszyński A (2005) [Restoration of European bison in West Poland—conception, status and protection]. In: Materials of conference “Protection of West Pomeranian bison” Ińsko, 15–16 September 2005: 5–11 [In Polish]
Bleier N, Lehoczki N, Újváry D, Szemethy L, Csány S (2012) Relationships between wild ungulates density and crop damage in Hungary. Acta Theriol 57:351–359. doi:10.1007/s13364-012-0082-0
Bunevich AN (2003) [Dynamics and spatial structure of European bison population in the Białowieża Forest]. Belovezhskaya Pushcha. Issledovaniya 11:160–177 [In Russian]
Bunevich AN (2004) Spatial structure and movements of European bison in the Belarussian part of Białowieża Forest. In: Krasińska M, Daleszczyk K (eds) Proceedings of the conference “European Bison Conservation”. Mammal Research Institute, Białowieża, Poland, pp 23–26
Caughley G (1970) Eruption of ungulate populations, with emphasis on Himalayan tahr in New Zealand. Ecology 51:53–72. doi:10.2307/1933599
Clutton-Brock TH, Albon SD, Guinnes FE (1988) Reproductive success in male and female deer. In: Clutton-Brock TH (ed) Reproductive success: studies of individual variation in contrasting breeding systems. University of Chicago Press, Chicago, pp 325–343
Cromsigt JPGM, Kerley GIH, Kowalczyk R (2012) The difficulty of using species distribution modeling for the conservation of refugee species—the example of European bison. Divers Distrib 18:1253–1257. doi:10.1111/j.1472-4642.2012.00927.x
Daszkiewicz P, Samojlik T (2009) [French pioneer of prehistorical research in Białowieża]. Matecznik Białowieski 2:16–17 [In Polish]
Demographic Yearbook of Poland (2009) Central Statistical Office, Warszawa. http://www.stat.gov.pl/gus/5840_3697_ENG_HTML.htm
D’Eon RG (2004) Snow depth as a function of canopy cover and other site attributes in a forested ungulate winter range in southeast British Columbia. B C J Ecosys Manage 3:1–9
Garretson MS (1938) The American bison: the story of its extermination as a wild species and its restoration under federal protection. New York Zoological Society, New York
Gates CC, Larter NC (1990) Growth and dispersal of an erupting large herbivore population in Northern Canada: the Mackenzie wood bison (Bison bison athabascae). Arctic 43:231–238, http://arctic.synergiesprairies.ca/arctic/index.php/arctic/article/view/1616/1595
Handley LJL, Perrin N (2007) Advances in our understanding of mammalian sex-biased dispersal. Mol Ecology 16:1559–1578
Hayward MW, Kowalczyk R, Krasiński ZA, Krasińska M, Dackiewicz J, Cornulier T (2011) Restoration and intensive management have no effect on evolutionary strategies. Endangered Species Research 15:53–61. doi:10.3354/esr00371
Hofman-Kamińska E, Kowalczyk R (2012) Farm crops depredation by European bison (Bison bonasus) in the vicinity of forest habitats in northeastern Poland. Environ Manag 50:530–541. doi:10.1007/s00267-012-9913-7
Jacobs J (1974) Quantitative measurement of food selection A modification of the forage ratio and Ivlev's electivity index. Oecologia 14:413–417. http://www.jstor.org/stable/4214938
Jędrzejewska B, Jędrzejewski W (1998) Predation in vertebrate communities. The Białowieża Primeval Forest as a case study. Springer, Berlin
Kerley GIH, Kowalczyk R, Cromsigt JPGM (2012) Conservation implications of the refugee species concept and the European bison: king of the forest or refugee in a marginal habitat? Ecography 35:519–529. doi:101111/j1600-0587201107146x
Komers PE, Messier F, Gates CC (1992) Search or relax: the case of bachelor wood bison. Behav Ecol Sociobiol 31:195–203, http://www.jstor.org/stable/4600740
Komers PE, Messier F, Gates CC (1993) Group structure in wood bison: nutritional and reproductive determinations. Can J Zool 71:1367–1371. doi:10.1139/z93-188
Kowalczyk R, Schmidt K, Jedrzejewski W (2012) Do fences or humans inhibit the movements of large mammals in Białowieża Primeval Forest? In: Somers MJ, Hayward MW (eds) Fencing for conservation. Springer, New York, pp 235–244
Kowalczyk R, Taberlet P, Coissac E, Valentini A, Miquel C, Kamiński T, Wójcik JM (2011) Influence of management practices on large herbivore diet—case of European bison in Białowieża Primeval Forest (Poland). Forest Ecol Manage 261:821–828
Kozlo PG, Bunevich AN (2009) [European bison in Belarus]. Beloruskaya Novuka, Minsk [In Russian]
Krasińska M, Krasiński ZA (1995) Composition, group size, and spatial distribution of European bison bulls in Białowieża Forest. Acta Theriol 40:1–21
Krasińska M, Krasiński ZA (2007) European bison. The Nature Monograph. Mammal Research Institute Polish Academy of Science, Białowieża
Krasińska M, Krasiński ZA, Bunevich AN (2000) Factors affecting the variability in home range size and distribution in the Polish and Belarussian part of the Białowieża Forest. Acta Theriol 45(3):321–334. http://www.zbs.bialowieza.pl/g2/pdf/1303.pdf
Krasiński ZA (1978) Dynamic and structure of the European bison population in the Białowieża Primeval Forest. Acta Theriol 23:3–48
Krasiński ZA, Krasińska M, Leniec H (1994) The European bison in the Knyszyn Forest. Parki Nar Rez Przyr 13:107–114 [In Polish with English summary]
Larter NC (1988) Diet and habitat selection of an erupting wood bison population. Dissertation University of British Columbia, Vancouver
Larter NC, Gates CC (1990) Home ranges of wood bison in an expanding population. J Mammal 71:604–607, http://www.jstor.org/stable/1381800
Larter NC, Sinclair ARE, Ellsworth T, Nishi J, Gates CC (2000) Dynamics of reintroduction in an indigenous large ungulate: the wood bison of northern Canada. Anim Conserv 3:299–309. doi:10.1111/j.1469-1795.2000.tb00115.x
Loe LE, Mysterud A, Veiberg V, Langvatn R (2009) Negative density-dependent emigration of males in an increasing red deer population. Proc Biol Sci 276:2581–2587. doi:10.1098/rspb.2009.0224
Lykkja ON, Solberg EJ, Herfindal I, Wright J, Rolandsen CM, Hanssen MG (2009) The effects of human activity on summer habitat use by moose. Alces 45:109–124
McHugh T (1958) Social behaviour of the American buffalo (Bison bison bison). Zoologica 43(1):1–40
Meagher M (1973) The bison of Yellowstone National Park. Science Monograph 1. National Park Service, Government Printing Office, Washington, DC
Meagher M (1998) Range expansion by bison of Yellowstone National Park. J Mammal 70:670–675, http://www.jstor.org/stable/pdfplus/1381449
Nemtsev AS, Rautian GS, Puzachenko AYu, Sipko TP, Kalabushkin BA, Mironenko LV (2003) [Wisent in Caucasus] Kachestvo, Moskva-Maikop. [In Russian]
Paszkiewicz R (2004) The use of tree stands by the population of the European bison of Lowland-Caucasian line western Bieszczady. Parki Nar Rez Przyr 23:647–656 [In Polish with English summary]
Pérez-González J, Carranza J (2009) Female-biased dispersal under conditions of low male mating competition in a polygynous mammal. Mol Ecol 18:4617–4630. doi:10.1111/j.1365-294X.2009.04386.x
Perzanowski K, Januszczak M (2004) Preliminary assessment of the home range dynamic of the European bison Bison bonasus in Bieszczady Mountains. Parki Nar Rez Przyr 23:639–646 [In Polish with English summary]
Plumb GE, White PJ, Coughenour MB, Wallen RL (2009) Carrying capacity, migration, and dispersal in Yellowstone bison. Biol Conserv 142:2377–2387. doi:10.1016/j.biocon.2009.05.019
Potapov E, Bedford A, Bryntesson F, Cooper S, Brown B (2011) Impact of snow cover on movements and habitat choice by suburban white-tailed deer (Odocoileus virginianus). Bull N J Acad Sci 56:5–8
Pucek Z, (ed) Belousova IP, Krasińska M, Krasiński ZA, Olech W (2004) European bison. Status Survey and Conservation Action Plan. IUCN/SSC Bison Specialist Group, IUCN. Gland, Switzerland and Cambridge, UK
Pyziel AM, Kowalczyk R, Demiaszkiewicz AW (2011) The annual cycle of shedding Eimeria oocysts by European Bison (Bison bonasus) in the Białowieża Primeval Forest, Poland. J Parasitol 97:737–739
Raczyński J (2011) European bison Pedigree Book. Białowieża National Park, Białowieża
Radwan J, Demiaszkiewicz AW, Kowalczyk R, Lachowicz J, Kawałko A, Wójcik JM, Pyziel A, Babik W (2010) An evaluation of two potential risk factors, MHC diversity and host density, for infection by an invasive nematode Ashworthius sidemi in endangered European bison (Bison bonasus). Biol Conserv 143:2049–2053
Ramsrud J (2007) Calving site selection by moose: anti-predation versus feeding conditions, M. Sc. Thesis. Norwegian University of Science and Technology, Trondheim, Norway
Reynolds PE (1998) Dynamics and range expansion of a reestablished muskox population. J Wildl Manage 62:734–744, http://www.jstor.org/stable/3802350
Roe EF (1970) The North American buffalo. University of Toronto Press, Toronto, Canada
Shaw JH, Carter TS (1990) Bison movements in relation to fire and seasonality. Wildl Soc Bull 18:424–430, http://www.jstor.org/stable/3782742
Sokołowski AW (2004) [Woods of the Białowieża Forest]. State Forests Information Centre, Warszawa [In Polish]
Stankowich T (2008) Ungulate flight responses to human disturbance: a review and meta-analysis. Biol Conserv 141:2159–2173
Statistical Yearbook of Agriculture (2009) Central Statistical Office. Warszawa. www.stat.gov.pl
Sztolcman J (1926) [European bison, history, behaviour and future]. The Central Union of Polish Associations of Hunting, Warszawa [In Polish]
Tokarska M, Pertoldi C, Kowalczyk R, Perzanowski K (2011) Genetic status of the European bison Bison bonasus after extinction in the wild and subsequent recovery. Mammal Review 41:151–162
Tokarska M, Marshall T, Kowalczyk R, Wójcik JM, Pertoldi C, Kristensen TN, Loeschcke V, Gregersen VR, Bendixen C (2009) Effectiveness of microsatellite and SNP markers for parentage and identity analysis in species with low genetic diversity: the case of European bison. Heredity 103:326–332
Wilson GA, Olson W, Strobeck C (2002) Reproductive success in wood bison (Bison bison athabascae), established using molecular techniques. Can J Zool 80:1537–1548
Winnie J Jr, Creel S (2007) Sex-specific behavioural responses of elk to spatial and temporal variation in the threat of wolf predation. Anim Behav 73:215–225
Wołk E, Krasińska M (2004) Has the condition of European bison deteriorated over last twenty years? Acta Theriol 49:405–418
Wójcik JM, Kawałko A, Tokarska M, Jaarola M, Valenback P, Pertoldi C (2009) Post-bottleneck mtDNA diversity in a free-living population of European bison: implications for conservation. Journal of Zoology 277:81–8
Acknowledgments
The study was financed by the Polish Ministry of Science and High Education (grant 2P04F 011 26) and by budgets from the Mammal Research Institute PAS and the Białowieża National Park. We would like to thank Drs. Matt Hayward and Joris Cromsigt for revisions and valuable comments to earlier versions of the manuscript. We are grateful to Ms. Elizabeth Finch who corrected the English.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by: Jan M. Wójcik
Electronic supplementary material
Below is the link to the electronic supplementary material.
ESM 1
(DOC 27 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution License which permits any use, distribution, and reproduction in any medium, provided the original author(s) and the source are credited.
About this article
Cite this article
Kowalczyk, R., Krasińska, M., Kamiński, T. et al. Movements of European bison (Bison bonasus) beyond the Białowieża Forest (NE Poland): range expansion or partial migrations?. Acta Theriol 58, 391–401 (2013). https://doi.org/10.1007/s13364-013-0136-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13364-013-0136-y