
Abstract
The concept of autopoiesis was proposed 40 years ago as a definition of a living being, with the aim of providing a unifying concept for biology. The concept has also been extended to the theory of knowledge and to different areas of the social and behavioral sciences. Given some ambiguities of the original definitions of autopoiesis, the concept has been criticized and has been interpreted in diverse and even contradictory ways, which has prevented its integration into the biological sciences where it originated. Here I present a critical review and conceptual analysis of the definition of autopoiesis, and propose a new definition that is more precise, clear, and concise than the original ones. I argue that the difficulty in understanding the term lies in its refined conceptual subtlety and not, as has been claimed by some authors, because it is a vacuous, trivial or very complex concept. I also relate the concept of autopoiesis to the concepts of closed systems, boundaries, homeostasis, self-reproduction, causal circularity, organization and multicellularity. I show that under my proposed definition the concept of a molecular autopoietic system is a good demarcation criterion of a living being, allowing its general integration into the biological sciences and enhancing its interdisciplinary use.
Similar content being viewed by others

Introduction
The search for a definition of life or of a living being has been recurrent in biological thinking, and has acquired a renovated interest due to the appearance of research programs related to artificial life, the origin of life and astrobiology (reviewed in Popa 2004). The Chilean biologists Humberto Maturana and Francisco Varela (1973; 1980) proposed that the minimal definition of a living being may be based on only one property, which they called autopoiesis. This contrasts sharply with other definitions, which are based on the combination of several properties (e.g., Koshland 2002; Popa 2004, pp. 6–7).Footnote 1 Autopoiesis also contrasts with other definitions by being a relational property (also referred to as systemic, functionalist, formalist or universalist). Its relational character is due to the fact that it does not depend directly on the nature (type, intrinsic properties) of the components of those entities which have the property, but rather depends only upon the relations between the components. Because of this, the concept connects to more general areas of research which have attempted to describe diverse types of systems in relational terms, such as tektology (Bogdanov 1996 [1922], Gorelik 1975, 1984; Dudley 1996), general systems theory (Bertalanffy 1950, 1968; Rapoport 1972), cybernetics (Wiener 1948; Ashby 1957; Varela and Maturana 1972), relational biology (Rashevsky 1954; Rosen 1958, 1959, 1991), information theory (Shannon 1948; Shannon and Weaver 1949; Avery 2003) and the recent research areas of complex systems (Kauffman 1989, 1993; Ruthen 1993; Bak 1996; Reason and Goodwin 1999; Gisiger 2001; Wolfram 2002; Bedau 2003, 2008), systems biology and synthetic biology (Wolkenhauer 2001; Breitling 2010).
The heuristic value and the advantages of relational definitions and descriptions of living beings are prominent in the research on artificial life, astrobiology and the origin of life (Dix 1983, Fleischaker and Margulis 1986; Emmeche 1992; Kauffman 2001; Popa 2004; McMullin 2004; Luisi 2006). One reason for this is that a definition of a living being that does not depend on the type of components which constitute the system would in principle account for remote living beings close to the origin of life—which were possibly much different from current forms (Reilly 2008; Wolfe-Simon et al. 2011), and for artificial life synthesized in vitro or in silico (Langton 1989; Breitling 2010) and living beings which may potentially exist outside Earth (Popa 2004). In all these cases the living beings may be possibly composed of components which are very different from those of currently known living beings (Friedman 2002; Nealson 2002). A relational definition is also attractive from a theoretical standpoint. We normally are not willing to consider as a living being an aggregate of components which have just any type of interrelation; for example, a paste of organic material from a living being which has been ground or centrifuged would not usually be considered a living being. On the other hand, it is not clear that we would in principle refuse to consider as living beings entities which are not based on DNA, polymers or even carbon atoms, that are the basic components of current living beings (see Reilly 2008). Thus it seems reasonable to think that what lets us distinguish living beings as such is based more on the nature of the relations between their components than on the nature of the components themselves. Finally, given the amount of speculation which exists about the origin of life, the specific pathways which originated life on Earth may be less relevant than the general relational characteristics or “family of mechanisms” which generated life (Pross 2004).
The high degree of abstraction of the concept of autopoiesis and its relational character allowed its authors to use this property as the base for a theoretical development which connects autopoiesis with problems of epistemology, cognitive sciences (Maturana and Varela 1972; 1980; 1992 [1987], Varela et al. 1991; Bitbol and Luisi 2004; Barandiaran and Moreno 2006) and evolutionary biology (Maturana and Varela 1992 [1987], Maturana and Mpodozis 1992, 2000; Vargas 2005). The concept has also been used in environmental sciences (Margulis 1997; Mann 1991; Sagan and Margulis 1997; Litfin 2010) and several areas of human sciences, including business administration (Mingers 1995), psychology (Colapinto 1985; Simon 1985; Snyder 1999) and particularly sociology (Luhmann 1986, 1995 [1984], 2007 [1997], Geyer and van der Zouwen 2001; Magalhaes and Sanchez 2009). The impact of the concept has increased and currently, only in ISI Web of Knowledge, the concept appears in more than 30 articles and 300 citations by year (Fig. 1). However, even though its central objective is to understand and delimit the object of study of biology, the concept has not managed to become a definition of living being accepted among biologists, nor has it been incorporated in biology texts or integrated generally into biological disciplines. I will argue that this is due to at least two factors: first, the original definitions are neither clear nor easy to understand; and second, the concept appears difficult to apply in practice. I will show that neither of those factors are actual obstacles to take autopoiesis as a general definition of living being. To do so, I will indicate the causes of this lack of clarity, analyze the possible interpretations of the original definition of Maturana and Varela, and propose a new, clearer and more precise definition, which I propose is the natural result of the analysis of this concept.

Number of articles and citations by year published in ISI Web of Knowledge in which appear the terms ‘autopoiesis’, ‘autopoietic’, ‘autopoyesis’, or ‘autopoyetic’. a. Articles in Science Citation Index Expanded (SCI-EXPANDED), Social Sciences Citation Index (SSCI), and Arts and Humanities Citation Index (AandHCI). b. Articles only in SCI-EXPANDED. c. Citations in SCI-EXPANDED, SSCI, and AandHCI. d. Citations only in SCI-EXPANDED
Autopoiesis: A Variety of Interpretations
The interpretation of the concept of autopoiesis has been controversial (Fleischacker 1988; Zolo 1990, Swenson 1992a, 1992b; Scheper and Scheper 1996; Morán et al. 1999; Escobar 2011). To begin with, the difficult rhetoric of the authors together with their persistent allusions to self-reference and circularity have produced skepticism and have made some authors suspect that the concept could mean just about anything, have no meaning, be tautological, trivial or not very scientific, which would also apply to its derivations (Zolo 1990; Munch 1992; Scheper and Scheper 1996, Swenson 1992a, 1992b, Manriquez and Rothhammer 1997; Gallardo 1997; Nespolo 2003; Valenzuela 2007, 2010). Others have considered the concept “to be nothing but a fancy synonym for both self-organization and self-maintenance” (Mahner and Bunge 1997, p. 144) and thus immediately discountable as a definition of a living being, since self-organization is a property shared by many types of non-living systems (Mahner and Bunge 1997; Pross 2004, Swenson 1992b).
At the other extreme, autopoiesis has been treated as a highly complex property, which would explain the manifest difficulty in the attempts to formalize it and in the search for concrete examples to illustrate the concept, even in purely formal terms. The attempts to formalize the concept of autopoiesis have achieved highly sophisticated results, but it is not clear in what sense these could be considered successful. A first model based on cellular automata (Varela et al. 1974) was later corrected and invalidated by one of its authors (McMullin and Varela 1997; McMullin 2000). A first attempt at a general formalization of an autopoietic system using Spencer-Brown’s very complex calculus of indications (Varela 1979) has not produced satisfactory results (Letelier et al. 2003). Later formalizations based in the very abstract theory of categories and Rosen’s (M,R) systems (Letelier et al. 2003, 2006; Cornish-Bowden et al. 2007) only recently have found a first concrete example of (M,R) system what might be considered a formal autopoietic system (Letelier et al. 2006). This line of investigation has gone to the extreme of claiming that autopoiesis is not computable, that is that autopoietic systems cannot be calculated by Turing machines (Letelier et al. 2003). All this has managed to surround the concept of autopoiesis with an air of mystery, in which it is not clear whether it is a trivial concept (which includes everything or almost everything) or empty (which includes nothing or almost nothing), whether it is useful scientifically, whether it is an extremely sophisticated property or is really very simple.
How can we explain such a wide range of interpretations of the concept? Here I will argue that it is because autopoiesis is characterized by a delicate conceptual subtlety which has not been sufficiently clear. I will show that (i) the concept has not been subject to sufficient conceptual analysis before looking for examples or formalizations, (ii) more than one meaning can be extracted from the definition, and these alternatives have not been clearly distinguished nor has it been determined which is correct, and (iii) it has not been systematically made explicit using precise concepts and making clear their relations with other relevant concepts for the understanding of the basic operation of living beings. I will then attempt to remedy these failings, by analyzing the original definition proposed by the authors and replacing it with a new definition which I feel is clearer and more concise. With this new definition, I will analyze how the concept of autopoiesis is related to other concepts such as closed system, boundary, homeostasis, self-reproduction, circular causality and organization. This critical analysis pretends to come alive again the possibility to take autopoiesis as a general and widely acceptable minimal definition of living being for biology.
Autopoiesis: A Subtle Property
Autopoiesis literally means ‘self-production’ or ‘self-creation’. The concept of autopoiesis in the book De Máquinas y Seres Vivos (On Machines and Living Beings) written in 1972 (Maturana and Varela 1994, p. xvii) was defined explicitly, as follows:
‘An autopoietic machine is a machine organized as a system of processes of production of components concatenated in such a way that they produce components which:
-
i).
generate the processes (relations) of production which produce them through their continual interactions and transformations, and
-
ii).
constitute the machine as a unit in physical space’ (Maturana and Varela 1973)Footnote 2
A good exercise for the present purpose is to read the original definition several times (if one has sufficient patience). As immediately becomes evident, this definition is relatively difficult and ambiguous (as is the definition of 1980, see Note 1) due to the lack of a precise reference of some of the terms utilized, which will be indicated in italics in what follows: ‘that they produce components which’; ‘generate the processes (relations) of production’; ‘that produce them by means of their continual’. It is not clear whether the terms in italics refer to the “system of processes”, the “processes of production”, to the “concatenated products” that produce, or the “concatenated components” produced. This may well explain the diversity of interpretations of the concept.
Although there is ambiguity in the definition, it may be inferred that autopoiesis defines a type of system constituted by a set of types of components (which I will call X) which, by means of concatenated processes, produces a set of types of components (which I will call Y). Now, what defines Y? i.e., What conditions must Y fulfill in order that a system of production of components (X) may be considered autopoietic?
The most common interpretation (and literal in almost all the paraphrases of the original definitionFootnote 3) is that the network of production of components produces all the components of the network, that is, X = Y. As we shall see, this interpretation is inacceptable given known basic aspects of living beings.
In the first place, if we understand that the ‘components of the system’ (X) are all the reactants and products of the reactions which occur in the system (that is, including its “food” and “waste”), then if we suppose that the system generates all the components of the system (X = Y) we must conclude that autopoietic systems are closed systems. Thus, given that living beings are open systems, we must reject this interpretation.
In the second place, if we consider that the ‘components of the system’ (X) are all the reactants and products of the reactions which occur in the system except its “food” and “waste” (supposing that there is a simple way to distinguish them in relational terms), then if we suppose that the system generates all the components of the system (X = Y) we are faced with two other problems. First, many (and perhaps) all known living beings need for their maintenance elements from their surroundings which form part of their metabolic network and which may act as coenzymes or cofactors, and, above all, which could not be distinguished from any other intermediate metabolites produced internally by the system. Second, if what the system produces permanently is the same set of components of the system (X = Y); this would imply that the system cannot suffer qualitative change over time, that is, it may not have qualitative changes during ontogenesis (such as metamorphosis in insects, Valenzuela 2007). Thus, given that living beings do have ontogeny and may depend upon cofactors or metabolites from their surroundings, this second interpretation must also be rejected. As a consequence, the classical image of M. C. Escher, much used to represent the concept of autopoiesis, of two hands each drawing the other completely (including the pencil) is not an acceptable representation (Fig. 2).

Photograph of Drawing Hands by M. C. Escher, 1948, taken in Escher Museum, The Hague. The usual association between autopoiesis and the drawing of two hands drawing each other is questionable as an analogy of autopoiesis. [Photograph taken by Gabriel Razeto]
According to the above, we are left with only one possible interpretation, that is, that the autopoietic system produces only a subset of the components of the system (Y∈X). In summary, the three characteristics of living beings; (1) they are open systems, (2) they may require coenzymes or metabolites from the environment for their maintenance, and (3) they may change during ontogeny, restrict the possible interpretations of the concept of autopoiesis to one. The network does not produce all the components of the network, only some of them. That is, the network produces a subgroup of the components which characterize the network as a unit.
But now the obvious following question arises, what subgroup of a production network must the network produce in order to be considered autopoietic? Given the functional centrality which molecular biology has given to the enzymes, we might be tempted to define an autopoietic system as “a network of processes which produces all the catalysts of the network”. However, as discussed above, there are many exceptions (coenzymes and cofactors mentioned in point (2) above) and it does not appear to be theoretically necessary for a system to produce all the catalysts of the network in order to be autopoietic, since if these catalysts are available in the environment and may enter the system constantly, the system may continue to maintain the processes of production of the network without producing all the catalysts internally.Footnote 4
From the above I conclude that in part (i) of the original definition of autopoiesis (Maturana and Varela 1973, 1980) there is no specification which permits to demarcate or characterize the subgroup Y that allows a production network to be called autopoietic; thus it is necessary to consider seriously part (ii) of the original definition: ‘constitutes a machine as a unit’. That is, here is the only possible specification of the subgroup Y, viz., the subgroup of components which the system must produce to maintain the system as a unit over time. Based on the above I propose that the only possible interpretation of the concept of autopoiesis which is not empty or trivial, and which is consistent with basic aspects of known living beings, may be summarized in the following short definition of autopoiesis, which I will call D1:
D1: An autopoietic system is a network of processes which produces all the components whose internal production is necessary to maintain the network operating as a unit.
Even though this definition is more clear, precise and concise than the original, it contains some subtleties which should be clarified. Definition D1 implies that, whatever the system be that has to produce (Y) to maintain its production network operating, it will depend on what is available in the environment for it. Thus we cannot impose a priori what components or types of components the network must internally produce (to maintain its production network operating), since this will depend upon the nature of its surroundings. This may explain the difficulty in defining autopoietic organization mathematically, since as is natural traditional formalizations symbolize the system only based on the components of the system (e.g., Letelier et al. 2003, 2006; Cornish-Bowden et al. 2007). Under definition D1, it may be concluded that the autopoietic organization of a system can only be formalized mathematically if components of the environment of the system are included.
Autopoiesis and Molecular Autopoiesis
The 1973 definition of Maturana and Varela suffered a small but significant change in Varela et al. (1974), which is also included in the definition of Maturana and Varela (1980) (see Note 1). The original definition of autopoiesis makes reference to a system as a unit “in physical space”; however, the later definitions refer to a unit “in the space in which the components exist” (not using the word “physical”). This allows the authors to distinguish between the concept of autopoiesis, which is general and abstract and could in principle be applied to any “space” or “dominion” (physical or not physical); and the concept of “molecular autopoiesis”, “chemical autopoiesis”, or “autopoiesis in physical space”, which is the definition of living being as such.Footnote 5 This generated a first differentiation between the thinking of Maturana and Varela. For Maturana there is no restriction in the “space” in which an autopoietic system may exist (Maturana 1981), while for Varela autopoiesis applied to non-physical domains is only metaphoric (Varela 1981).
It is the abstract (not necessarily physical) use of the concept of autopoiesis which has been the base of its success in human sciences such as psychology and sociology, in which, for example, the “space” of production of components is constituted by communications (Luhmann 1986, 1995 [1984], 2007 [1997], Razeto-Barry and Cienfuegos 2011; Veloz et al. 2012). For this reason I maintained the abstraction of the general concept of autopoiesis in definition D1, without specifying the space or domain in which it is applied. Thus D1 may also serve to make the concept more clear and precise in human sciences. However, in what follows I will examine autopoiesis in physical space, where the concept attempts to serve as a definition of living being, and only will comment on a few implications for extra-biological applications of the concept (see Notes 8 and 18).
Traditionally, molecular autopoiesis has been understood as a minimal definition of a single cell, and ‘processes of production’ simply as chemical reactions, and thus autopoietic systems are understood as a sub-class of collectively autocatalytic chemical reaction networks (McMullin 2000). As I will discuss later in this article, this is an inadequate concept of autopoiesis for unicellular and particularly for multicellular living beings.
Autopoiesis and Boundaries
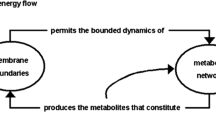
Varela followed a second line somewhat different from that of Maturana. In later works, Varela emphasizes a particular component produced by the system, the boundary; that is, the physical border which limits and separates the interior from the exterior of the system. The boundary, or its cellular parallel, the external membrane of the cell, is not mentioned in the original definitions of the concept (Maturana and Varela 1973, 1980), however, appears explicitly in the first computational examples led by Varela (Varela et al. 1974; McMullin and Varela 1997), and are incorporated explicitly in later paraphrases and explanations of the original definition (e.g., Varela 2000, also in Luisi et al. 1996).
Apparently for Varela the production of the boundary or external membrane from the interior of the system is essential for autopoiesis (see also Fleischacker 1988), that is, if the components of the membrane came from the exterior the system would not be autopoietic (i.e., for Varela one type of element necessary for Y is the boundary component). At first sight this makes sense; for one thing the membrane is effectively a particularly important component which permits the maintenance of the network of component production of the current unicellular living organisms. Also, the production of the external membrane is in reality an important factor which allows these organisms to become established and maintain themselves as units in space, and also in proto-cells generated in vitro (Luisi et al. 2006; Solé et al. 2007).
Notably, this line emphasized by Varela has recently led to the artificial production of chemical systems in the laboratory which some authors consider autopoietic. The Swiss scientist Pier Luigi Luisi and collaborators produced synthetic membranes in the form of spherical vessels which catalyze the production of the components of the membrane; they grow and reproduce spontaneously as a consequence of their self-production (Zepik et al. 2001; Luisi 2006). Although ‘self-reproduction’ and ‘autopoiesis’ (‘self-production’) have been considered definitions of life in competition, the possibility of considering some kinds of self-reproducing chemical systems as autopoietic was first suggested by Luisi and Varela (1989, Luisi 1993). This opened the possibility of considering as autopoietic both “homeostatic” systems (whose mass remains constant) and “self-reproductive” systems (whose mass grows until it spontaneously provokes reproduction) (Luisi and Varela 1989; Luisi et al. 1996; Luisi 1993, 2003, 2006; Varela 2000). From the point of view of the previous discussion, this “experimental shift” with respect to the original interpretation of the concept (although not the definition) is acceptable and completely compatible with definition D1.
It is worth noting that, in spite of the compatibility of D1 with the “shift” of Varela, the generation of a layer forming a physical barrier as a kind of production of a boundary is not necessarily a universal requirement in causal terms, and thus is not a requirement of definition D1. The idea that the production of the components of a barrier is necessary for the maintenance of the system comes from the image of life as a cell with molecules in a liquid state dissolved in an aqueous solvent. However, the physical state of cellular cytoplasm is viscous (Schroder 2010) and under certain circumstances might maintain its unity (and therefore its border) by cohesion and surface tension (as does a drop of water surrounded by air). It is probable that in the origin of the first cells on Earth the production of a membrane was by the self-assembly of simple lipids synthesized in the environment, while the internal production of membrane components was a derived process evolved by natural selection (Hargreaves and Deamer 1978; Hanczyz et al. 2003; Budin and Szostak 2011).Footnote 6 On the other hand, it is possible that in the origin of life the membrane was not a “limit” which separated the interior from the exterior of the living being, but rather the membrane itself was the living being (the membrane would be the interior of the living being, Fig. 3).Footnote 7

Representation of the non-intuitive idea that the system “outside” of a living system may be spatially “inside”. In the origin of life metabolism may ocurr in the membranes rather than inside the space surrounded by the vesicles. Note that Robert Hook (1635–1703) who discovered cells, thought that the spatial inside of cells is not living matter (Mazzarello 1999)
It is also worth noting that the requirement of the internal production of the components of the membrane is not a requirement of the definition itself, but rather a practical condition imposed by the known cellular forms of living beings on Earth. According to D1, if the system did not require a membrane to maintain itself as a cohesive unit, or if the components of the membrane came from the environment and the system continued to produce the subgroup of components necessary for its maintenance as a spatially discreet unit, then the system would continue to be considered autopoietic.
It is important to mention here that the aim of the concept of autopoiesis is to characterize a minimal definition of life. That is, a definition that includes the minimal requirements (semantically) necessary to consider that a system is a living system. Even if studies suggest that on Earth the proposition: “all current autopoietic systems on earth produce the elements of their membranes” (and equally replacing ‘autopoietic systems’ by ‘living beings’) probably is true, the question about the definition of autopoiesis (and of life) implies deciding whether this proposition will be a trivial tautology guaranteed by the definition of autopoiesis (or of living being) or a synthetic empirical result due to current observations and predictions based on current life on the Earth. The minimal life project searches for a minimal definition of life, that is, ‘the definition should… be minimal but specific enough (i.e., it should include just those elements that are common to all forms of life – not being, in principle, restricted to life on Earth…).’ (Ruiz-Mirazo et al. 2004, p. 326, similarly see Emmeche 1998). In this context it is clear that it is preferable to take the mentioned proposition as an interesting synthetic property of known nature rather than an analytical tautology guaranteed by definition, maintaining open the door to include other kinds of life in the definition, together with the one in the origin of life in Earth, for which it is possible that the mentioned proposition is not true (Hargreaves and Deamer 1978; Hanczyz et al. 2003; Budin and Szostak 2011).
In this vein, as we will see below in our discussion about multicellular living beings, as a direct consequence of D1, a multicellular organism is an autopoietic system as such (i.e., it is a “first-order” autopoietic system), and in it the production of a “border” (e.g., an epidermis in vertebrate animals) is not precisely what maintains it as a cohesive unit.Footnote 8
Autopoiesis, Homeostasis and Self-Reproduction
The vesicles generated in the laboratory by Luisi and collaborators are autopoietic according to definition D1, however, they are essentially self-reproductive and cannot maintain themselves dynamically in homoeostasis (without growth or permanent reproduction). This characteristic does not agree with the usual emphasis that Maturana and Varela gave to the concept of autopoiesis, which is based on the maintenance (homeostasis) of the system by means of the regeneration of elements which must necessarily disintegrate spontaneously (Varela et al. 1974; McMullin and Varela 1997; Varela 2000; McMullin 2004). Thus from my point of view it is not necessary to include the ‘disintegration’ and ‘regeneration’ of components as an essential element in the formalization of autopoiesis, in contrast to the original emphasis of its authors and the formal models of autopoiesis (e.g., Varela et al. 1974; McMullin and Varela 1997; Letelier et al. 2003, 2006; Cornish-Bowden et al. 2007; Cornish-Bowden and Cárdenas 2007). As a consequence, the common theoretical contraposition between autopoiesis and self-reproduction as strictly competing ideas (e.g., Cornish-Bowden and Cárdenas 2007) is not necessary. Although Varela (2000; Luisi et al. 1996) admits that autopoietic systems may be in homeostasis or in growth tending to self-replication, he systematically emphasizes the regeneration of degraded elements as an essential factor of autopoiesis. By contrast, I propose that even though there may be autopoietic systems which are not self-reproductive, and self-reproductive systems which are not autopoietic, also there may perfectly well be autopoietic systems which are strictly self-reproductive (such as the special type of self-catalytic network generated in the laboratory by Luisi and collaborators). That is, there may be autopoietic systems without disintegration of their components, and thus in constant growth and sef-reproduction.
It should be clear that the definition D1 I propose is compatible both with systems whose mass is maintained in homeostasis and with systems whose mass is in constant growth (auto-catalytic and auto-reproductive networks). If the products of an auto-catalytic network generate the conditions necessary to maintain the network as a discrete unit in space, then the network is an autopoietic system. As a consequence, I reject the recently proposed idea that autopoietic systems are a Robert Rosen (M,R) type of system (Letelier et al. 2003, 2006; Cornish-Bowden et al. 2007; Cornish-Bowden and Cárdenas 2007), since it is essential that a (M,R) system have degradation and reparation.Footnote 9 It is worth noting that no one has yet generated in the laboratory an artificial system (or a concrete model for the evolution of life on Earth) which is a homeostatic autopoietic system, which without doubt is the most recurrent conception of an autopoietic system, both in the original formulation of its authors and in its formalizations (Maturana and Varela 1972; 1980; Varela et al. 1974; Varela 1979; McMullin and Varela 1997; Varela 2000; McMullin 2004; Letelier et al. 2003, 2006; Cornish-Bowden et al. 2007; Cornish-Bowden and Cárdenas 2007).
Autopoiesis and Circular Causality
Maturana and Varela often emphasized that autopoiesis implies a certain circularity. In their terms, an autopoietic system is open in structure (components enter and leave) but is closed in its organization (its relations are closed as in a circle).Footnote 10 From this idea came others which the authors used, such as ‘operational closure’Footnote 11, ‘operative closure’, ‘recursivity’, and ‘self-referential systems’. Here we address the question: How should this circularity be understood? Firstly, what seems clear is that it is a circular causality. Footnote 12 The notion of circular causality first arose with the appearance of cybernetics, around 1946 (Umpleby 2005), in which the feedback mechanism was represented graphically as a cycle (a circle with a defined direction). For the creators of cybernetics, the cyclic vision of the feedback mechanism assumes a representation of nodes and relations between nodes, which is collectively closed on itself and in which the nodes represent states of particular variables of a system, and the relations between nodes represent the control relations of these states (Fig. 4). Now, what type of causal circularity is autopoiesis?

Abstract representation of a causal cycle
Obviously all circularity is a circularity of some kind of relations. The causal circularity of feedback proposed by the cyberneticists is a circular relation among control relations. Autopoiesis is usually interpreted as a causal circularity of production relations (see the first quotation in Note 10), which is consistent with the literal interpretation of the concept as ‘self-production’. However, according to the earlier arguments, this is not possible. Circularity means that the leaving and entering relations are the same. Starting from a given point, the relations of the cycle must return to the same point (Fig. 4). If the relation is that of production, then starting from any point—all components of the system composing the unit—by means of the relations of production, all the components of the system should be produced. That is, using the earlier nomenclature, if the set X produces the set Y, (X → Y), that the production of the system is circular implies X = Y. However, we have already rejected this. If autopoiesis were a circular causality of production, then the types of components which constitute the system should (indirectly, mediated by a complicated network of relations) produce themselves; that is, produce exactly the same types of components. However, an autopoietic system only produces a subset of all the types of elements of the system (Y∈X). Then, in what sense can its organization be understood as circular (supposing that this is in fact possible)? Definition D1 may be of help.
A living being is undoubtedly a system of production (in which production refers specifically to chemical reactions). Although the components of the system do not produce all the components of the system, they do produce that subset of components ‘whose internal production is necessary to maintain the network function as a unit’ (D1). And since the system must maintain itself to meet D1, this subset of components must be capable of procuring the internal presence of the rest of the components of the system from the exterior, bringing them “in” and maintaining them in a sufficiently local proximity to compose a physical unit. That is, it is not especially relevant whether the functional components of the system (we may exclude the “waste”) come from the environment or are produced internally, what is important is that they be present.Footnote 13 If the chemical reactions of a system produce 90 % of its components and the other 10 % are taken from the environment, this system is no more autopoietic than one which produces 10 % of its components and takes the other 90 % from its environment; for this reason there cannot be “degrees” of autopoiesis.Footnote 14 This means that there is circularity, but even in a system based on production the circularity does not come from the production relations, but rather from a circularity of physical proximity. Thus the basis of the circularity is that the physical proximity of the components of the system allows the production of components whose internal production is necessary for the physical proximity of the components of the system (that which constitutes the network as a discrete unit in space). This may allow us to understand Varela’s emphasis on the production of membrane components that spontaneously form vesicles as a very practical specification of the concept of autopoiesis; not because the production of a boundary is in itself essential, but rather because the membrane is an efficient manner of maintaining a molecular system united, especially in an aquatic medium. If the system does not control molecules diffusion reactants’ dilution will lead to the cessation of chemical reactions in which they are involved.Footnote 15
This causal relation may be profitably understood as a ‘probabilistic causality’.Footnote 16 That is, what the physical proximity of the components of a system does is to increase the probability that the components of the system are constantly in physical proximity. Thus, autopoiesis would be a particular type of circular probabilistic causality. Physical proximity (or spatial-temporal cohesion) is not a scientifically unimportant quality; on the contrary, it is the base of all chemical reactions, catalysis in particular. If we want to produce a chemical reaction, what we have to do is bring the reactants into close proximity, which greatly increases the probability that the product will be produced. In fact, this is exactly what a catalyst does; an enzyme is a “molecular approximation machine”.
In summary, paraphrasing Maturana and Varela, an autopoietic machine is a homeostatic system of chemical production which has its own individual identity as a variable which it maintains constant.Footnote 17 ‘Individual identity’ is understood to be the same as ‘numerical identity’ or ‘quantitative identity (reviewed in Noonan 2009), that is, the maintenance of a discrete spatio-temporal unit (i.e., without losing spatio-temporal continuity or increasing or decreasing its ‘number’ or ‘quantity’), although its properties or qualities may change (i.e. losing its ‘qualitative identity’, see Noonan 2009). Thus the only condition on the chemical system is that it maintains constantly (by its chemical production) its components in physical proximity in such a way that it insures the spatio-temporal continuity which characterizes it as a unit.Footnote 18
It must be kept in mind that a living system is not only a physical system but notably also a chemical system. What distinguishes a living bacterium from a frozen bacterium or a crystal is that the living being is a machine characterized by chemical reactions. A crystal maintains its individual identity by itself (and the physical proximity of its constituents) trivially, due to the immobility of the relative positions of its components, but it is not a production machine. It is very easy in the laboratory to generate a crystal spontaneously; however, generating a concatenated system of chemical reactions which maintains its individual identity thanks to the products of these reactions is something that only has been achieved recently by Luisi and collaborators artificially in the laboratory, and they still have not been able to make the system homeostatic. From the chemical point of view, what is impressive about living systems is their exceptional coherence of chemical reactions which maintain stably (i.e. without disintegrating or spreading out in a continuous massFootnote 19) a discrete unit in space-time.
From the above we extract that possibly the idea of autopoiesis has seemed trivial to some because it is not commonly emphasized that chemical reactions (i.e. processes of chemical production) are not similar, in relevant aspects, to the springs of physicists or the circuits of engineers. Chemical reactions imply the constant modification of the individual identity of the components involved (reactants which become products), which in general impedes a component from maintaining its function over time (the main exceptions are the enzymes). This makes it very difficult to produce or even outline formally the conditions which permit the continued existence of an autopoietic system based on chemical reactions. This does not occur with the homeostasis of a servomechanism or the self-organization of a tornado or a Bénard cell. In these cases it is true that a tornado or a Bénard cell are units whose dynamics make probable the incorporation of components from their surroundings which maintain their individual identity. However, what makes them different from a living being is that living beings are systems which produce components in the form of chemical reactions, in which the individual identity of the components is constantly modified. The maintenance of individual identity of a tornado does not involve more variables than the position and velocity of the particles in its locality. The particles themselves are not modified in the whirlwind; they enter and leave without modifying their identity. They have the same form when they enter and when they leave, they leave being “themselves”, without substantial modification.Footnote 20
In summary, and as a specification of D1 in physical space, I propose a schematic definition of molecular autopoiesis (D2):
D2: A system is autopoietic if and only if:
-
1.
It is a network of physical and chemical processes.
-
2.
This network chemically produces a subset of the components which are parts of the network.
-
3.
This subset of components, by means of relations among its members and with the components of its surroundings, generates the conditions necessary to maintain the components of the network in physical proximity, collectively forming a spatially discrete individual unit over time.
Autopoiesis and Organization
One of the greatest sources of confusion generated around the concept of autopoiesis has been the use that Maturana and Varela gave to the concept of ‘organization’. One phrase of Maturana and Varela (that was paraphrased previously) was used recurrently to explain what an autopoietic system is: “A living system is a homeostatic system whose homeostatic organization has its own organization as the variable that it maintains constant” (Maturana and Varela 1980, p. 48). I will show that the emphasis of the authors in the maintenance of self-organization is difficult to understand due to the sense in which the word ‘organization’ is used by these authors.
At first the reader tends to interpret ‘organization’ in the usual way, that is, as the configuration of the existing relations among the components of a system; as “the relations which determine…the dynamics of the interactions and transformations of the components” (Maturana and Varela 1994, p. 67). But later in the text one understands that, according to the authors, the organization which is maintained constant in the case of an autopoietic system is the type of organization which defines living beings, that is, autopoietic organization. Thus the questioned affirmation is equivalent to saying “an autopoietic system is a system which maintains its autopoietic organization as the variable which it maintains constant”. That is like saying that “a chair is an object that maintains its chair organization as the variable which it maintains constant”. That is, the affirmation would be only a non-informative tautology (i.e., by definition of its terms it cannot be false). On the other hand, if by ‘organization’ we understand “the relations which determine…the dynamics of the interactions and transformations of the components” (as is usual), the affirmation would imply a system composed of relations which do not change over time, a system without ontogeny, without individual history (and of course an insect in metamorphosis would not be a living being, see Valenzuela 2007); that is, something very different from the idea of a living being. As we saw before, definition D1 implies that what characterizes an autopoietic system is not its capacity to oppose changes in its organization, but rather its capacity to oppose the disintegration of its spatio-temporal unity (in spite of its constant chemical reactions). This also implies that the concept of ‘organizational invariance’ (Letelier et al. 2003, 2006; Cornish-Bowden et al. 2007) is not an acceptable term as a formalization of the concept of autopoiesis.
Some of the confusion related to the concept of autopoiesis are associated with a number of the pseudo-problems and classical confusions which have been generated in philosophy and continue to the present day, due to not distinguishing between ‘individual identity’ (=‘quantitative’ or ‘numeric’ identity) and ‘qualitative’ identity (these pseudo-problems are reviewed in Tugendhat 2002 [1992] and Tugendhat and Wolf 1997 [1983]). Individual identity refers to what specifies particular individuals (or ‘tokens’ in philosophical jargon), while qualitative identity refers to what specifies the classes (or kinds) of individuals defined according to their properties (or ‘types’). This confusion arises principally because both concepts are referred to using the ambiguous terms of “identity” or “self” .Footnote 21 Maturana clearly defines ‘organization’ as that which specifies the type or class to which an individual or system pertains, distinguishing it from the ‘structure’ that specifies the characteristics of the individual as a particular system or “token” .Footnote 22 However, the authors tend to specify an autopoietic system as a system which achieves the production and maintenance of a same (constant) organization. That is, unless the authors want to fall into a simple tautology by making ‘organization’ equal ‘autopoietic organization’, they are affirming that autopoietic systems are systems which produce that which maintains their qualitative identity (their properties) constant. That is, systems which do not change their properties during their lives! In other words, the affirmation is either tautological or is false. Put another way, autopoietic systems do not maintain an organizational homeostasis (homeostasis of their qualitative identity), but rather a homeostasis of their individual unity (homeostasis of their quantitative identity). This is the reason for the paraphrase based on D1: An autopoietic machine is a homeostatic system of chemical production which has its own individual identity as the variable which it maintains constant.Footnote 23
Another of the pseudoproblems related to autopoiesis due to not distinguishing between qualitative identity and individual (or quantitative) identity is the so-called problem of ‘infinite regression’ stated by Rosen (1959) in the causal circularity of living beings (Letelier et al. 2003, 2006; Cornish-Bowden and Cárdenas 2008). Robert Rosen believed that there was a kind of paradox in the regeneration of components, in which the components of a living being are produced by other components of the being, which in turn are produced by other components of the being, and so on. However, the error lies in not considering that the regeneration is a replacement of elements of the same type. There is nothing paradoxical in a series of events among particular elements (tokens) producing reactions which generate other particular elements. What makes the system stable over time is that the elements produced are approximately of the same type (approximately, of course, unless there is no change of qualities during ontogeny). In fact, in autopoiesis there is regeneration of both the types of elements and the types of relations (which, as an “epiphenomenon”, produce the regeneration of the process of regeneration). Thus there is no “infinite regression” whatsoever in living beings which is different from any other habitual causal process in nature, including any cybernetic machine.Footnote 24
Autopoiesis and Multicelularity
The essentiality that biology gives of chemical reactions to understand the life is well represented in Helmont’s (1648) claim: ‘All life is chemistry’ (from Pross 2004). Certainly, the main sources of control of processes in living systems are chemical reactions. However, even in unicellular organisms and minimal cell prototypes many necessary processes to the system maintenance are different from chemical reactions or processes catalyzed by chemical reactions of the system. For example, in the model of Varela et al. (1974, McMullin and Varela 1997) there are reactions of concatenation and disintegration that are not catalyzed by products of the system (McMullin 2000). Furthermore, processes of molecules diffusion and vesicles formation are physical spontaneous processes in which there are not changes of chemical substances. Thus, in living beings not all processes (relations) necessary to maintain the system are, or are catalyzed by, chemical reactions. In addition, multicellular living beings depend essentially on many physical (non chemical) processes to its maintenance. For example, circulatory, lymphatic, and respiratory systems in animals are based on fluids flowing within a network of vessels, valves and chambers, in which changes of pressures, heat release and many other physical phenomena are fundamental to organism’s survival. A single mechanical obstruction of a cerebral artery may cause the death and disintegration of a multicellular organism (see also Goldenfeld and Woese 2011).
Maturana and Varela (1973, 1980) briefly characterized multicellular organisms as “second-order autopoietic systems”, which means that they are made up of (first-order) autopoietic systems.Footnote 25 However, they do not affirm that multicellular organisms are first-order autopoietic systems. Maturana and Varela (1992 [1987]) faced directly this problem: ‘What is the organization of metacellulars?… Are some metacellulars autopoietic unities? That is, are second-order autopoietic systems also first-order autopoietic systems?’ (p. 87). However, authors finally avoided giving a response: ‘For the purposes of this book, we shall leave open the question of whether or not metacellular systems are first-order autopoietic systems.’ Because ‘in the case of metacellulars, we are still ignorant of the molecular processes that would constitute those metacellulars as autopoietic unities comparable to cells.’ (pp. 88–89) and characterized multicellular organisms only as “lineages” of cells. This point becomes critical to autopoiesis becomes a unifying concept for biology and a good definition of living being. If multicellular organisms (the paradigmatic living beings, Wilson 2000) are not autopoietic then multicellular organisms are not living beings! In fact, according to Weber and Varela (2002), multicellular organisms are not themselves autopoietic systems: ‘Autopoiesis has been explicitly formulated for the minimal living system, the cell. (…) A multicellular organism (and this includes all vertebrates usually taken as prototypical organisms) is not in itself an autopoietic unit of second order, since its organization does not follow the same selfcontructing principles.’ (p. 115).
It is possible that Maturana and Varela did not include multicellular organisms as autopoietic systems because of their misguided emphasis in chemical reaction networks and in the production of a boundary, which as I have analyzed previously, are unnecessary for the definition of autopoiesis. According to definitions D1 and D2, multicellular organisms are clearly (first-order) autopoietic systems. The ontogeny of mammals develops from a unicellular (zygote) state to a morula, a blastula and so on, as a consequence of the chemical production of a subgroup of the components which maintain it as a discrete multicellular unit. It is important to note that multicellular organisms such as adult humans produce their own boundary (e.g., the skin in vertebrates, shells and cuticles in invertebrates and plants, respectively); however, in contrast with unicellular organisms, the production of the border is not what physically maintains the system as a unit. We can think of multicellular formations of Myxobacteria in Prokarya or Dictyostelium in Eukarya, but also of human beings tortured by ancient indigenous Moche people in South America whom took the skin off of their prisoners while maintaining them alive (Castillo 2000).Footnote 26 Essential components of multicellular organisms are chemical products that maintain cells joined. Probably the first step in the evolution of multicellularity was the chemical production of components that prevent the daughter cells from separating after the replication (Bonner 1998). It seems that this kind of processes is not improbable because multicellularity has evolved independently many times in each domain of life (Bonner 1998; Carroll 2001). Thus the network of interconnected components of multicellular organisms, chemically produce the components that maintain the components of the network in physical proximity, collectively forming a spatially discrete individual unit over time.
Autopoiesis and Life
Now that the definition of autopoiesis has been re-examined and redefined, it may be seen that the definition of ‘living being’ as a ‘molecular autopoietic system’ is a good definition which serves as a demarcation criterion of living beings. That is, autopoiesis understood in its subtle formulation (D1 and D2) is a singular relational property which effectively allows the differentiation of the main groups of systems which we recognize as living beings from the rest of the objects and non-living systems. As we have seen, the ‘self-production’ characteristic of autopoiesis is not the production of the totality of the components which the system materializes, nor is it the production of the same organization of components and relations which give rise to the products of the network, but rather the production of the subgroup of components which actively conserve the continuous discrete existence of the system (that is, which conserve the spatio-temporal individual identity of the system in spite of the chemical reactions it undergoes). This definition excludes not only the machines generated so far by humans (such as automobiles) and typical inanimate natural objects (such as crystals), but also excludes self-organized dynamic systems such as whirlwinds, Bénard cells, and other natural physical systems as dissipative structures.
A definition of a living being should also serve as a demarcation criterion for “borderline beings”, those at the edge of life. Autopoiesis excludes several such beings such as viruses, viroids, transposons and prions, since these are not systems which have chemical reactions whose products maintain the discreet individual unity in space. Other borderline systems are the apparently “superorganisms” such as those of social insects or colonies of coelenterates; in this case the epithet autopoietic is less clear, since the spatial unity is somewhat less clear. In spite of this, the individual identity of a colony is usually quite clear and may be followed in space and time; these must be included as living beings. This may be unintuitive conclusion is consistent with recent approaches that understand “organismality” as a continuous variable characterized by a high level of cooperation and low level of conflict among parts. These approaches consider that colonies of insects are “organisms” in the same degree than an individual animal or human being (Queller and Strassman 2009; West and Kiers 2009).Footnote 27
One of the permanent difficulties in the definitions of living beings is that they must satisfy two conditions which are in constant tension; they must be compatible with the theories of the origin of life on Earth, in which reproduction appears to be essential, and they must not fall into the “mule paradox” (Szathmáry 2002), that is, conclude that non-reproductive systems such as mules are not living beings. One classical dispute in the theories of the origin of life is whether self-maintained metabolism preceded self-replication or whether self-replication occurred first (Pross 2004). In any case, it appears that the basic characteristic of the most primitive living beings (pre-cellular) is the self-reproduction (of singular molecules or complete metabolic systems) (Pross 2004; Razeto-Barry 2012), which is a key condition for their propagation and evolution. However, any definition which requires self-reproduction as a definitive condition for life falls into the “mule paradox”, that is, it excludes individuals which are clearly living beings but which are not self-reproducing (such as mules and some castes of eusocial insects). The subtle definitions D1 and D2 resolve this tension, because both the growth which tends to self-reproduction and the homeostatic maintenance (not self-reproductive) of a system are special cases which comply with the definition of autopoiesis. Thus both self-reproductive vesicles (e.g., Zepik et al. 2001; Luisi 2006) and non-reproductive organisms (mules, naked mole rats, worker bees, plants without seeds) are autopoietic systems as long as some subset of the products of their internal chemical reactions provides the maintenance of the individual identity of the system. It seems evident that self-reproductive autopoiesis is the ancestral condition, but non self-reproducing autopoiesis has arisen many times in the past, and re-occurs with every organism born infertile (see also Razeto-Barry 2012).
The above is highly relevant for the compatibility of autopoiesis with the theories of the origin of life. The fact that definitions D1 and D2 make autopoiesis compatible with the non-homeostatic self-reproduction without internal disintegration of components is what allows autopoiesis to be compatible with any currently plausible theory of the origin of life on Earth (e.g., see Pross 2004). This is because in general the half life (or spontaneous decay) of the components of living beings is much greater than the life span of the organisms. In fact, the decay of the polymers of currently living beings is the result of complex degradation networks (such as the systems of protein degradation). However, it is hard to imagine that in the origin of life there were sophisticated molecular systems (Pross 2004) such as those required for the degradation of enzymes and other products of metabolism. Thus homeostatic autopoietic systems with internal degradation of components are derived living beings and not primitive ones.Footnote 28 Definitions D1 and D2 not only resolve the mule paradox, but also allow a definition of living being compatible with origin of life theories and serves as a good criterion for the demarcation of uni- and multicellular living beings.
Probably one of the greatest defects of the concept of autopoiesis as a definition of a living being is that it is only a descriptive definition and not mechanical-causal. That is, the property in question does not refer to causes or forces which may be analyzed within physical, chemical or thermodynamic theories. One consequence of this is that the autopoietic property also does not explain other properties of living beings in causal terms. A good definition not only should give the conditions for recognition of a living being, but also the conditions for the production and manipulation of living systems (Escobar 2011). For example, one might require that every reaction of an autopoietic system be spontaneous, but autopoiesis does not specify the thermodynamic conditions which allow the complete system to function as an autonomous machine, nor does it define its energetic relation with the environment. The specification of the directive forces may be essential to the understanding of the nature of life (Pross 2004). However, in spite of this shortcoming, it appears that autopoiesis (under D1 and D2) is the only simple property actually available which allows a satisfactory definition of living being, satisfying the majority of the theoretical and practical criteria of a good definition.
Final Conclusions
The analysis of the concepts of autopoiesis (D1) and molecular autopoiesis (D2) have led me to conclusions which contrast strongly with an important part of the literature which deals with autopoiesis. Here I provide a summary of the points in which this work opposes other publications.
First, I conclude that the autopoiesis of a system does not imply the production of all the components of the system, in contrast to the opinions of Capra 1996; Letelier et al. 2003 and Luisi 2003 (see note 3); thus the circularity of autopoiesis is not a causal circularity of production relations, and therefore I reject the concept of “metabolic closure” (Letelier et al. 2003, 2006) or “closure of the reaction network” (McMullin 2000, p. 167) as a characterization of autopoiesis.
Second, I conclude that the concept of autopoiesis is not tautological, trivial or unscientific, and neither are its derivations, in contrast to the conclusion of a number of authors (Zolo 1990; Munch 1992; Scheper and Scheper 1996, Swenson 1992a, 1992b, Manríquez and Rothammer 1997; Gallardo 1997; Nespolo 2003; Valenzuela 2007, 2010),
Third, the concept of autopoiesis is not highly complex, nor does it imply an undecidable or un-computable process, as suggested by Letelier et al. (2003, 2006) (see also the seventh conclusion below).
Fourth, the internal production of the border or external membrane is not essential to the concept of autopoiesis, in contrast to the analyses and formalizations of Varela et al. (1974), Fleischaker (1988), Luisi et al. (1996), McMullin and Varela (1997), and Varela (2000).
Fifth, multicellular organisms are first-order autopoietic systems (in contrast to Weber and Varela 2002).
Sixth, the disintegration and regeneration of disintegrated components is not essential to autopoiesis, in contrast to Varela et al. (1974), McMullin and Varela (1997), Varela (2000), Letelier et al. (2003, 2006), McMullin (2004), Cornish-Bowden et al. (2007), Cornish-Bowden and Cárdenas (2007).
Seventh, and as a consequence of the previous, I conclude that autopoietic systems are not a type of Robert Rosen’s (M,R) system, in contrast to the conclusions of Letelier et al. (2003, 2006), Cornish-Bowden et al. (2007) and Cornish-Bowden and Cárdenas (2007). One consequence of this is the third conclusion above.
Eighth, I conclude that the concept of autopoiesis is not in contraposition to that of self-reproduction (in contrast, for example, to Cornish-Bowden and Cárdenas 2007) and that the homeostatic autopoietic systems described by the majority of the formalizations of the concept (Maturana and Varela 1972; 1980; Varela et al. 1974; Varela 1979; McMullin and Varela 1997; Varela 2000; McMullin 2004; Letelier et al. 2003, 2006; Cornish-Bowden et al. 2007; Cornish-Bowden and Cárdenas 2007) are only particular cases of an autopoietic system, which do not correspond to the systems which characterized the origin of life, which in turn implies that homeostatic autopoiesis is an evolutionary derived, not primitive property.
Ninth, I conclude that an autopoietic system is not a system which maintains the properties of its organization (type) constant, in contrast to Maturana and Varela (1973, 1980), due to which I reject the term ‘organizational invariance’ of Letelier et al. (2003, 2006) and Cornish-Bowden et al. (2007) as a formalization of the concept of autopoiesis.
Tenth, I conclude that the so-called problem of ‘infinite regression’ of Rosen (1959), Letelier et al. (2003, 2006) and Cornish-Bowden and Cárdenas (2008) is a pseudo-problem produced by not distinguishing between the concepts of ‘individual identity’ (quantitative) and ‘qualitative identity’.
Notes
See Ruiz-Mirazo et al. 2004 for a criticism of this kind of descriptive, “check-out list” definition
In Maturana and Varela (1980, p. 79) the definition was: ‘An autopoietic machine is a machine organized (defined as a unity) as a network of processes of production (transformation and destruction) of components that produces the components which:
-
(i).
through their interactions and transformations continuously regenerate and realize the network of processes (relations) that produced them; and
-
(ii).
constitute it (the machine) as a concrete unity in the space in which they (the components) exist by specifying the topological domain of its realization as such a network’.
-
(i).
To give a few examples (italics mine): ‘All components of an autopoietic network are produced by other components in the network’ (Capra 1996, p. 167). ‘In this way the entire network continually “makes itself”. It is produced by its components and in turn produces those components’ (Capra 1996, p. 98). ‘In an Autopoietic system, the result of any given process is the production of components that eventually would be transformed by other processes in the network into the components of the first process.’ (Letelier et al. 2003, p. 266). ‘An autopoietic unit is a system that is capable of self-sustaining owing to an inner network of reactions that re-generate all the system’s components’ (Luisi 2003, p. 51). Note that, unfortunately, these definitions are not clearly differentiated from the descriptions of closed systems. There are also more ambiguous notions as: ‘autopoiesis… means “self-renewing”’ (Skyttner 2005, p. 60)
To my knowledge, no organisms have been reported to date that directly obtain a functional enzyme from its environment, but the requirement of cofactors and coenzymes have been widely described, which are necessary to catalysis processes. Anyway, it seems evident that if we discover that a kind of bacteria directly obtains some functional enzymes from its environment, that would not negate that they are living beings. Indeed, the bacterial production of enzymes was discovered much later than the discovering of bacteria, which were considered living beings from the very beginning (Porter 1976). Note that in the model of Varela et al. (1974) there are catalysts that are not produced by any reaction in the network. Note also that if consider membranes as catalytic structures in the sense that they ‘emerge unchanged from the catalyzed reaction, while drastically increasing the reaction rate’ (McMullin 2000) and that the membrane of the first living beings probably self-assembled from simple lipids synthesized in the environment (Hargreaves and Deamer 1978; Hanczyz et al. 2003; Budin and Szostak 2011), then the first living beings on Earth would be examples of living systems with catalysts coming from the environment.
Maturana and Varela usually differentiated between autopoiesis understood as the ‘organizing principle’ of living beings, and living beings as the ‘physical realization’ of autopoiesis (see Fleischacker 1988).
It is perhaps worth mentioning that the border of viruses is formed by proteins; however, since there is no dynamic processes of production (metabolism) within this border, viruses cannot be considered to be autopoietic systems.
I owe this last observation to Carlos Medina.
Another advantage of not including the production of a boundary in the definition of autopoiesis is that non-molecular autopoietic systems, as society, may be more clearly characterized as autopoietic systems. In fact, one of the major objections to consider society autopoietic is the absence of a production of a boundary which makes sense: ‘These ideas are based, in my opinion, on an abuse of language. In autopoiesis, the notion of boundary has a more or less precise meaning. When, however, the net of processes is transformed into an “interaction among people”, and the cellular membrane is transformed into the limit of a human group, one falls into an abuse, as I expressly said.’ (Varela 2000, see also Luisi 2003). Note that medieval cities were defined by their boundaries (a closed defensive wall), however this kind of definition is not applicable in modern societies.
The symbol (M,R) comes from ‘metabolism-repair systems’, also called ‘metabolism-replacement systems’ by Letelier et al. (2006). Essential to any materialization of a (M,R) system is the decay of the catalysts, which is counterbalanced by a group of replacement reactions which re-synthesizes the catalysts (Cornish-Bowden and Cárdenas 2007, 2008). By contrast, in an autocatalytic network the catalysts do not necessarily decay (or their half-life is much greater than the life of the organisms). I thank Carlos Medina for bringing this point to my attention.
The authors make reference to the circularity of autopoiesis in most of their works. Here I take only two examples (from Maturana and Varela 1980): ‘…living systems as systems defined as unities through the basic circularity of their production of their components’ (xiv); ‘Living systems… are…all organized in a closed causal circular process’ (p. 9, italics are mine).
According to Fleischacker (1988), the term ‘organizational closure’ was replaced by ‘operational closure’ by Varela since 1982.
Note that contemporary organizational approaches to function (e.g., Moreno and Ruiz-Mirazo 2009) do not ascribe functionality in any component of a living system created externally, which contrasts with the wide intuition that vitamins and cofactors are functional molecules which are not internally produced but obtained from the environment. Consistent with the present analysis the function of these molecules is guaranteed because they have the effect of allowing the maintainance of the autopoiesis, not matter whether they come from external or internal sources. Furthermore, many functional parts of multicellular living beings are microorganisms of other species, kingdoms and domains which are not internally produced but obtained from the environment (Pepper and Rosenfeld 2012; Costello et al. 2012).
It was recently argued that in a network of biochemical processes the distinction between ‘enzyme’ and ‘metabolite’ is arbitrary and unnecessary (Cornish-Bowden and Cárdenas 2007). This reinforces the fact that it is not necessary for the understanding of autopoietic organization to know what types of elements enter into the system or what role they play in the network of chemical reactions (e.g., whether they participate in ‘peripheral’, ‘intermediate’ or ‘central’ reactions of the network).
Any autocatalytic set ‘must be confined to a sufficiently small volume’ (Kauffman 1993, p.298).
A review of the notion of probabilistic causality and its application to evolutionary biology may be found in Razeto-Barry and Frick (2011).
According to Maturana and Varela (1973) “An autopoietic machine is a homeostatic system which has its own organization as the variable which it maintains constant” ([1994], p. 69, in this article all quotations from publications in Spanish are my translations). I criticize this affirmation in the next section.
Note that this definition of ‘individual identity’ (or ‘quantitative’ or ‘numerical’) is only applicable in the context of systems in physical space. To apply this analysis to other spaces or domains (social, linguistic, psychological, etc.), the concept of ‘individual identity’ (and its differentiation from ‘qualitative identity’) must be clearly specified.
This property may be related to thermodynamics as the maintenance of low entropy (see note 20).
From this viewpoint, I dare to propose that the thermodynamic characteristic recognized in living beings as being systems permanently away from thermodynamic equilibrium (Schneider and Kay 1994) may be considered as a consequence of the definition of autopoiesis applied to systems of concatenated chemical reactions. That is, the mere requirement that a system maintain its individual identity in spite of consisting of concatenated chemical reactions requires that the system be in permanent thermodynamic disequilibrium. This affirmation is not immediately obvious and must be demonstrated. However, it is plausible from the fact that systems far from equilibrium may tend to form local zones of order (Schneider and Kay 1994), and that the spatio-temporal congruence proper to the components of a unit with a continuous existence in space and time constitutes a source of order in itself (as happens with tornadoes).
A classical example of this is the woman who says “you’re not the same man I fell in love with”. Obviously she is not denying the “individual identity” of the man, she says it to him (e.g., Peter) and not to someone else. In fact, if she were talking to another person and not to Peter, we would either think she was crazy or had confused Peter with someone else. She means that he (the same individual, Peter) doesn´t have the same qualities he had when she fell in love with him; that is, she claims that his “qualitative identity” changed (see Tugendhat 2002 [1992]). The individual identity is not lost when properties change; what is lost is the qualitative identity.
According to the author: ‘The relations between components that define a composite unity (system) as a composite unity of a particular kind, constitute its organization.’ He defines ‘structure’ as ‘The actual components (all their properties included) and the actual relations holding between them that concretely realize a system as a particular member of the class (kind) of composite unities to which it belongs by its organization, constitute its structure.’ (Maturana 1980, “Introduction”, pp. xix-xxx, in Maturana and Varela 1980). Maturana (1978, p. 32) affirms that organization ‘refers to the relations between components that define and specify a system as a composite unity of a particular class, and determine its properties as such a unity’, while structure ‘refers to the actual components and the actual relations that these must satisfy in their participation in the constitution of a given composite unity’. I thank Aldo Mascareño for the last reference.
This also runs counter to studies of autopoiesis such as that of Cornish-Bowden and Cárdenas (2008), in which the maintenance of individuality is an accident, an abstract imposition which does not result from the dynamics of the system. The same can be said about the majority of computational models of autocatalytic reaction networks (see McMullin 2000).
It is possible that what made so attractive the introduction of the notion of ‘circularity’ in cybernetics due to the ‘circular causality’ (of feedback systems), is that we erroneously tend to think that if the causality of tokens is never backward (that is, there is never ‘backward causation’, see Faye 2010), then neither is the causality of types. The causal circularity of feedback is precisely a circularity of types of events, not of particular events.
‘What is common to all metacellulars in the five kingdoms is that they include cells as components of their structure. That is why we say that metacellulars are second-order autopoietic systems.’ (Maturana and Varela 1992, p. 87)
Note that the formation of external and internal boundaries in multicellular organisms is a derived evolved property, posibly increasing autonomy (Rosslenbroich 2009), analogously to the formation of external and internal membranes in the first cells (Hargreaves and Deamer 1978; Hanczyz et al. 2003; Budin and Szostak 2011) and in firsts unicellular eukaria (Kutschera and Niklas 2008), respectively. The idea of a living being as a “bag” containing liquid should be thus given up.
‘These [colonies of social insects] are sometimes referred to as superorganisms, but why not just call them organisms? From an adaptive perspective, a colony of social insects is analogous to the colony of cells that comprise a human’ (West and Kiers 2009). Following Queller and Strassman (2009), we may say that an autopoietic system is a system with high level of cooperation among parts based on “strong reciprocity” (see West et al. 2011 for a critical review), such that a typical homeostatic autopoietic system is a system with high “indirect reciprocity” among types of molecules (autocatalytic network) and some level of “punishment” (degradation of molecules).
I owe this point to Carlos Medina (see note 7).
References
Ashby WR (1957) An Introduction to Cybernetics. Chapman and Hall LTD
Avery J (2003) Information theory and evolution. World Scientific Publishing, London.
Bak P (1996) How Nature Works: the Science of Self-Organized Criticality. Springer-Verlag
Barandiaran X, Moreno A (2006) On what makes certain dynamical systems cognitive: a minimally cognitive organization program. Adapt Behav 14(2):171–185
Bedau M (2003) Artificial life: organization, adaptation and complexity from the bottom up. Trends in Cogn Sci 7(11):506–512
Bedau M (2008) What is life? In Sarkar, S. and Plutynski, A. (ed.) A Companion to the Philosophy of Biology. pp. 455–603. Wiley-Blackwell
Bertalanffy L (1950) The theory of open systems in physics and biology. In: Emery FE (ed) Systems thinking. Penguin, Harmondsworth, pp 70–85
Bertalanffy L (1968) General system theory. Penguin, Harmondsworth
Bitbol M, Luisi PL (2004) Autopoiesis with or without cognition: defining life at its edge. J R Soc Interface 1:99–107
Bognadonov A 1996 [1922]. Bogdanov’s Tektology. Dudley, P. (ed.) 1996, Centre for Systems Studies, Hull, U.K
Bonner JT (1998) The origins of multicellularity. Integeative Biol 1:28–36
Breitling R (2010) What is systems biology? Front Physiol 1(9):1–5
Budin I, Szostak JW (2011) Physical effects underlying the transition from primitive to modern cell membranes. PNAS USA 108(13):5249–5254
Capra F (1996) The Web of life. Anchor Books, New York
Carroll SB (2001) Chance and necessity: the evolution of morphological complexity and diversity. Nature 409:1102–1109
Castillo LJ (2000) Los rituales mochicas de la muerte. In Makowski, K. (ed.), Los Dioses del Antiguo Perú. Colección Arte y Tesoros del Perú. Lima, Banco de Crédito del Perú, pp., 103–135
Colapinto J (1985) Maturana and the ideology of conformity. The Family Therapy Networker (May/June): 29–30
Cornish-Bowden A, Cárdenas M-L (2007) Organizational invariance in (M, R)-systems. Chem Biodivers 4:2396–2406
Cornish-Bowden A, Cárdenas M-L (2008) Self-organization at the origin of life. J Theor Biol 252:411–418
Cornish-Bowden A, Cárdenas M-L, Letelier J-C, Soto-Andrade J (2007) Beyond reductionism: metabolic circularity as a guiding vision for a real biology of systems. Proteomics 7:839–845
Costello EK, Stagaman K, Dethlefsen L, Bohannan BJM, Relman DA (2012) The application of ecological theory toward an understanding of the human microbiome. Science 336:1255–1262
Dix D (1983) Toward a definition of life – semantic and thermodynamic considerations. J Theor Biol 102:337–340
Dudley P (1996) Back to basics? tektology and general system theory (GST). Syst Pract Action Res 9(3):273–284
Emmeche C (1992) Life as an abstract phenomenon: is artificial life possible? In: Towards a Practice of Autonomous Systems, ed. by F.J. Varen, F. Bourgine, Proc. 1st European Conference on Artificial Life. pp. 466–474
Escobar JM (2011) Autopoiesis and Darwinism. Synthese (in press). doi:10.1007/s11229-011-9875-y
Faye J (2010) Backward causation. Stanford Encyclopedia of Philosophy, Edward N. Zalta (ed.). Stanford
Fleischacker G (1988) Autopoiesis: the status of its system logic. Biosystems 22:37–49
Fleischacker G, Margulis L (1986) Autopoiesis and the origin of bacteria. Adv Space Res 6(11):53–55
Friedman I (2002) Short definitions of life. In: P’alyi G, Zucchi C, Caglioti L (eds) Fundamentals of life. Elsevier, New York, p 30
Gallardo MH (1997) Determinismo estructural: ¿teoría o dogma? Rev Chil Hist Nat 70:315–319
Geyer F, van der Zouwen J (2001) Sociocybernetics. Complexity, autopoiesis and observation of social systems. Greenwood Press, London
Gisiger T (2001) Scale invariance in biology: coincidence or footprint of a universal mechanism? Biol Rev 76:161–209
Goldenfeld N, Woese C (2011) Life is physics: evolution as a collective phenomenon far from equilibrium. Annu Rev of Condens Matter Phys 2:299–375
Gorelik G (1975) Principle ideas of Bogdanov’s ‘tektology: the universal science of Organization’. Gen Syst 20:145
Gorelik G (1984) Essays from tektology, 2nd edn. Intersystems, Seaside, Calif
Hanczyz MM, Fujikawa SM, Szostak JW (2003) Experimental models of primitive cellular compartmentalization: encapsulation, growth, and division. Science 302:618–622
Hargreaves WR, Deamer DW (1978) Liposomes from ionic, single-chain amphiphiles. Biochemistry 17:3759–3768
Kauffman SA (1989) Principles of adaptation in complex systems. Lectures in the Sciences of Complexity. The Santa Fe Institute Series (Addison Wesley), pp. 619–712
Kauffman SA (1993) The origins of order: self-organization and selection in evolution. Oxford University Press, Oxford
Kauffman SA (2001) Prolegomenon to a general biology, unity of knowledge: the convergence of natural and human science. Ann N Y Acad Sci 935:18–36
Koshland DE (2002) The seven pillars of life. Science 295:2215–2216
Kutschera U, Niklas KJ (2008) Macroevolution via secondary endosymbiosis: a Neo-goldschmidtian view of unicellular hopeful monsters and Darwin’s primordial intermediate form. Theory Biosci 127:277–289
Langton C (ed.) (1989) Artificial Life. Addison-Wesley
Letelier J-C, Marín G, Mpodozis J (2003) Autopoietic and (M, R) systems. J Theor Biol 222(2):261–272
Letelier J-C, Soto-Andrade J, Guiñez Abarzúa F, Cornish-Bowden A, Cárdenas M-L (2006) Organizational invariance and metabolic closure: analysis in terms of (M, R) systems. J Theor Biol 238(4):949–961
Litfin K (2010) Principles of gaian governance: a rough sketch. In: Christ E, Rinker HB (eds) Gaia in turmoil: climate change, biodepletion, and earth ethics in an age of crisis. The MIT Press, Cambridge
Luhmann N (1986) The autopoiesis of social systems. In: Geyer F, van der Zouwen J (eds) Sociocybernetic paradoxes. Sage, London
Luhmann N 1995 [1984]. Social Systems. Palo Alto, CA: Stanford University Press
Luhmann N 2007 [1997]. La Sociedad de la Sociedad. Universidad Iberoamericana, México
Luisi PL (1993) The chemical implementation of autopoiesis. In: Fleischaker GR, Stephano C, Pier Luigi L (eds) Self-production of supramolecular structures: from synthetic structures to models of minimal living systems. Kluwer, Dordrecht, pp 179–197
Luisi PL (2003) Autopoiesis: a review and a reappraisal. Naturwissenschaften 90:49–59
Luisi PL (2006) The emergence of life. From chemical origins to synthetic biology. Cambridge University Press, Cambridge
Luisi PL, Varela F (1989) Self-replicating micelles: a chemical version of minimal autopoietic systems. Orig Life Evol Biosph 19:633–643
Luisi PL, Lazcano A, Varela F (1996) Autopoiesis: the very idea, in: defining life. In: Rizzotti M (ed) The central problem in theoretical biology. Universitá de Padova, Padova, pp 146–167
Luisi PL, Ferri F, Stano P (2006) Approaches to semi-synthetic minimal cells: a review. Naturwissenschaften 93(1):1–13
Magalhaes R, Sanchez R (2009) Autopoiesis in organization theory and practice. Emerald, U. K
Mahner M, Bunge M (1997) Foundations of biophilosophy. Springer, Berlin
Mann C (1991) Lynn margulis: Science’s unruly earth mother. Science 252:378–381
Manriquez G, Rothhammer F (1997) Teoría moderna de la evolución. Amphora Editores, Santiago
Margulis L (1997) Kingdom animalia. In: Margulis L, Sagan D (eds) Slanted truths: essays on Gaia symbiosis and evolution. Springer, New York, pp 91–111
Maturana H (1978) Biology of language: the epistemology of reality. In: Millar G, Lennenberg E (eds) Psychology and biology of language and thought: essays in honour of Eric lennenberg. Academic, New York, pp 29–49
Maturana H (1981) Autopoiesis. In: Zeleny M (ed) Autopoiesis: A theory of living organization. North Holland, New York, pp 21–33
Maturana H, Mpodozis J (1992) Origen de las especies por medio de la deriva natural o la diversificaion de los linajes a traves de la conservacion y cambio de los fenotipos ontogenicos. Museo Nacional de Historia Natural, Publicación Ocasional 46:1–48
Maturana H, Mpodozis J (2000) The origin of species by means of natural drift. Revista Chilena de Historia Natural 73:261–310
Maturana H, Varela F (1973) De máquinas y seres vivos. Editorial Universitaria, Santiago, Chile
Maturana H, Varela F (1980) Autopoiesis and cognition. The Realization of the Living, Reidel, Dordrecht
Maturana H and Varela F 1992 [1987]. The Three of Knowledge, Shimbhala, Boston
Maturana H, Varela F (1994) De máquinas y seres vivos. Autopoiesis: La organización de lo viviente, 2nd edn. Editorial Universitaria, Santiago
Mazzarello P (1999) A unifying concept: the history of cell theory. Nat Cell Biol 1:E13–E15
McMullin B (2000) Remarks on autocatalysis and autopoiesis. Ann N Y Acad Sci 901:163–174
McMullin B (2004) Thirty years of computational autopoiesis: a review. Artif Life 10:277–295
McMullin B and Varela FJ (1997) Rediscovering computational autopoiesis. In P. Husbands and I. Harvey (Eds.), Proceedings of the Fourth European Conference on Artificial Life (pp. 38–47). Cambridge, MA: MIT Press. http://www.eeng.dcu.ie/˜alife/bmcm-ecal97/
Mingers J (1995) Self-producing systems: implications and applications of autopoiesis. PlenumPress, New York
Morán F, Moreno A, Minch E, Montero F (1999) In: Artificial Life V, Langton CG, Shimonara K (eds) Further steps toward a realistic description of the essence of life. MIT Press, London, pp 255–263
Moreno A, Ruiz-Mirazo K (2009) The problem of the emergence of functional diversity in prebiotic evolution. Biol Philos 24:585–605
Munch R (1992) Autopoiesis by definition. Cardozo Law Rev 13:1463–1471
Nealson K (2002) Short definitions of life. In: P’alyi G, Zucchi C, Caglioti L (eds) Fundamentals of life. Elsevier, New York, p 45
Nespolo RF (2003) Evolution by natural selection: more evidence than ever before. Rev Chil Hist Nat 76:699–716
Noonan H (2009) Identity. The Stanford Encyclopedia of Philosophy. Edward N. Zalta (ed.). Stanford
Pepper JW, Rosenfeld S (2012) The emerging medical ecology of the human gut microbiome. Trends Ecol Evol 27(7):381–384
Popa R (2004) Between necessity and probability: searching for the definition and origin of life. Springer, Berlin
Porter JR (1976) Antony van Leeuwenhoek: tercentenary of his discovery of bacteria. Bacteriol Rev 40(2):260–269
Pross A (2004) Causation and the origin of life, metabolism or replication first? Orig Life Evol Biosph 34:307–321
Queller DC, Strassman JE (2009) Beyond sociality: the evolution of organismality. Phil Trans R Soc Lond B 364:3143–3155
Rapoport A (1972) The uses of mathematical isomorphism in general system theory. Trends in General Systems Theory, Bertalanffy et al. (eds.). John Wiley and Sons
Rashevsky N (1954) Topology and life: in search of general mathematical principles in biology and sociology. Bull Math Biophys 16:317–348
Razeto-Barry, P. 2012. What should be the first law of biology? (forthcoming)
Razeto-Barry P, Cienfuegos J (2011) La paradoja de la probabilidad de lo improbable y el pensamiento evolutivo de Niklas Luhmann. Convergencia 57:13–38
Razeto-Barry P, Frick R (2011) Probabilistic causation and the explanatory role of natural selection. Stud Hist Phil Biol Biomed Sci 42(3):344–355
Reason P, Goodwin BC (1999) Toward a science of qualities in organizations: lessons from complexity theory and postmodern biology. Concepts and Transform 4(3):281–317
Reilly M (2008) Early life could have been based on arsenic. The New Scientist 198(2653):10
Rosen R (1958) A relational theory of biological systems. Bull Math Biophys 20:245–341
Rosen R (1959) A relational theory of biological systems II. Bull Math Biophys 21:109–128
Rosen R (1991) Life Itself. Columbia University Press, New York
Rosslenbroich B (2009) The theory of increasing autonomy in evolution: a proposal for understanding macroevolutionary innovations. Biol Philos 24:623–644
Ruiz-Mirazo K, Paretó J, Moreno A (2004) A universal definition of life: autonomy and open-ended evolution. Orig Life Evol Biosph 34:323–346
Ruthen R (1993) Adapting to complexity. Sci Am 268:130–140
Sagan D, Margulis L (1997) The uncut self. In: Margulis L, Sagan D (eds) Slanted truths: essays on Gaia symbiosis and evolution. Springer, New York, pp 59–73
Scheper WJ, Scheper GC (1996) Autopsies on autopoiesis. Behav Sci 41:1–12
Schneider ED, Kay JJ (1994) Life as a manifestation of the second law of thermodynamics. Math Comput Model 19(6–8):25–48
Schroder SM (2010) Toward a conceptual framework for biology. Q Rev Biol 85(3):293–318
Shannon CE (1948) A mathematical theory of communication. Bell Syst Tech J 27(379–423):623–656
Shannon CE, Weaver W (1949) The mathematical theory of communication. University of Illinois Press, Urbana-Champaign, IL
Simon R (1985) Structure is destiny – A frog’s view of the world. The Family Therapy Networker (May/June): 32–43
Skyttner L (2005) General systems theory. Problems, perspectives, practice, 2nd edn. World Scientific, London
Snyder E (1999) Is freud’s model of the mind autopoietic? Ger Rev 74:67–78
Solé RV, Munteanu A, Rodriguez-Caso C, Macía J (2007) Synthetic protocellbiology: from reproduction to computation. Philos Trans R Soc Lond B Biol Sci 362:1727–1739
Swenson R (1992a) Autocatakinetics, yes-autopoiesis, no: steps towards a unified theory of evolutionary ordering. Int J Gen Syst 21(2):207–228
Swenson R (1992b) Galileo, Babel, and autopoiesis (It's turtles all the way down). Int J Gen Syst 21(2):267–269
Szathmáry E (2002) “Short definitions of life” and “redefining life: an ecological, thermodynamic, and bioinformatic approach”. In: Pályi G, Zucchi C, Caglioti L (eds) Fundamentals of life. Elsevier, New York, pp 50, 181–195
Tugendhat E 2002 [1992]. Identidad personal, particular y universal. In Problemas, Gedisa, Barcelona
Tugendhat E and Wolf U 1997 [1983]. Propedéutica Lógicosemántica, Anthropos, Barcelona. [Original: Logisch-semantische Propädeutik, Philipp Reclam jun., Stuttgard]
Umpleby SA (2005) A history of the cybernetics movement in the United States. J Wash Acad Sci 91(2)
Valenzuela CY (2007) Within selection. Rev Chil Hist Nat 80:109–116
Valenzuela CY (2010) Neutral illusions, selective nightmares and autopoietic madness. Rev Chil Hist Nat 83:497–500
Varela FJ (1979) Principles of biological autonomy. Elsevier, New York
Varela FJ (1981) Describing the logic of the living – the adequacy and limitations of the idea of autopoiesis. In: Zeleny M (ed) Autopoiesis: A theory of living organization. North Holland, New York, pp 36–48
Varela FJ (2000) El fenómeno de la Vida. Dolmen Ensayo, Santiago, Chile
Varela FJ, Maturana HR (1972) Mechanism and biological explanation. Philos Sci 39(3):378–382
Varela FJ, Maturana HR, Uribe R (1974) Autopoiesis: the organization of living systems, its characterization and a model. Biosystems 5:187–196
Varela FJ, Thompson E, Rosch E (1991) The embodied mind. Cognitive science and human experience. MIT Press, Cambridge, MA
Vargas A (2005) Beyond selection. Revista Chilena de Historia Natural 78(4):739–752
Veloz T, Razeto-Barry P, Dittrich P & Fajardo A (2012) Reaction networks and evolutionary game theory. Journal of Mathematical Biology (forthcoming)
Weber A, Varela F (2002) Life after Kant: natural purposes and the autopoietic foundations of biological individuality. Phenomenol Cogn Sci 1:97–125
West SA, Kiers ET (2009) Evolution: what is an organism? Curr Biol 19(23):R1080
West SA, Mouden CE, Gardner A (2011) Sixteen common misconceptions about the evolution of cooperation in humans. Evol Hum Behav 32:231–262
Wiener N (1948) Cybernetics. Wiley, New York
Wilson JA (2000) Ontological butchery: organism concepts and biological generalizations. Philos Sci 67:S301–S311
Wolfe-Simon F, Jodi Switzer B, Kulp TR, Gordon GW, Hoeft SE, Pett-Ridge J, Stolz JF, Webb SM, Weber PK, Davies PCW, Anbar AD, Oremland RS (2011) A bacterium that can grow by using arsenic instead of phosphorus. Science 332:1163–1166
Wolfram S (2002) A new kind of science. Wolfram Media Inc., Champaign
Wolkenhauer O (2001) Systems biology: the reincarnation of systems theory applied in biology? Brief Bioinform 2(3):258–270
Zepik HH, Bloechliger E, Luisi PL (2001) A chemical model of homeostasis. Angew Chem 40:199–202
Zolo D (1990) Autopoiesis: critique of a postmodern paradigm. Telos 23:61–80
Acknowledgements
I thank Carlos Medina, Roberto Torretti, Luis Razeto, Lafayette Eaton, Gabriel Razeto and Leonardo Sepúlveda for useful comments on previous versions of this manuscript. I especially thank Carlos Medina for a number of ideas in this article (see notes 7, 9, and 28). I thank an anonymous reviewer for useful suggestions.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Razeto-Barry, P. Autopoiesis 40 years Later. A Review and a Reformulation. Orig Life Evol Biosph 42, 543–567 (2012). https://doi.org/10.1007/s11084-012-9297-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11084-012-9297-y