
Abstract
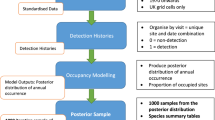
There is limited information available on changes in biodiversity at the European scale, because there is a lack of data from standardised monitoring for most species groups. However, a great number of observations made without a standardised field protocol is available in many countries for many species. Such opportunistic data offer an alternative source of information, but unfortunately such data suffer from non-standardised observation effort and geographical bias. Here we describe a new approach to compiling supranational trends using opportunistic data which adjusts for these two major imperfections. The non-standardised observation effort is dealt with by occupancy modelling, and the unequal geographical distribution of sites by a weighting procedure. The damselfly Calopteryx splendens was chosen as our test species. The data were collected from five countries (Ireland, Great Britain, the Netherlands, Belgium and France), covering the period 1990–2008. We used occupancy models to estimate the annual number of occupied 1 × 1 km sites per country. Occupancy models use presence-absence data, account for imperfect detection of species, and thereby correct for between-year variability in observation effort. The occupancy models were run per country in a Bayesian mode of inference using JAGS. The occupancy estimates per country were then aggregated to assess the supranational trend in the number of occupied 1 × 1 km2. To adjust for the unequal geographical distribution of surveyed sites, we weighted the countries according to the number of sites surveyed and the range of the species per country. The distribution of C. splendens has increased significantly in the combined five countries. Our trial demonstrated that a supranational trend in distribution can be derived from opportunistic data, while adjusting for observation effort and geographical bias. This opens new perspectives for international monitoring of biodiversity.




Similar content being viewed by others

References
Butchart SHM, Walpole M, Collen B, van Strien A, Scharleman JPW et al (2010) Global biodiversity: indicators of recent declines. Science 328:1164–1168
Clausnitzer V, Kalkman VJ, Ram M, Collen B, Baillie JEM, Bedjanic M, Darwall WRT, Dijkstra KB, Dowf R, Hawking J, Karube H, Malikova E, Paulson D, Schutte K, Suhling F, Villanuevam RJ, von Ellenrieder N, Wilson K (2009) Odonata enter the biodiversity crisis debate: the first global assessment of an insect group. Biol Conserv 142:1864–1869
De Heer M, Kapos V, ten Brink BJE (2005) Biodiversity trends in Europe: development and testing of a species trend indicator for evaluating progress towards the 2010 target. Philos Trans R Soc B 360:297–308
De Knijf G, Anselin A, Goffart P, Tailly M (eds) (2006) De libellen (Odonata) van België: verspreiding—evolutie—habitats [Dragonflies (Odonata) of Belgium: distribution—evolution—habitats]. Libellenwerkgroep Gomphus & Instituut voor Natuur- en Bosonderzoek, Brussel [Dutch with English abstract]
Dennis RLH, Thomas CD (2000) Bias in butterfly distribution maps: the influence of hot spots and recorders home range. J Insect Conserv 4:73–77
Dennis RLH, Sparks TH, Hardy BP (1999) Bias in butterfly distribution maps: the effect of sampling effort. J Insect Conserv 3:33–42
Devictor V, Whittaker RJ, Beltrame C (2010) Beyond scarcity: citizen science programmes as useful tools for conservation biogeography. Divers Distrib 16:354–362
Devictor V, van Swaay C, Brereton T, Brotons L, Chamberlain D, Heliölä J, Herrando S, Julliard R, Kuussaari M, Lindström Å, Reif J, Roy DB, Schweiger O, Settele J, Stefanescu C, van Strien A, van Turnhout C, Vermouzek Z, WallisDeVries M, Wynhoff I, Jiguet F (2012) Differences in the climatic debts of birds and butterflies at a continental scale. Nat Clim Chang. doi:10.1038/nclimate1347:1-4
Dingemans N, Kalkman VJ (2008) Changing temperature regimes have advanced the phenology of Odonata in the Netherlands. Ecol Entomol 33:394–402
Dommanget JL (2002) Protocole de l’inventaire cartographique des Odonates de France (Programme INVOD). Muséum National d’Histoire Naturelle, Société française d’odonatologie, Paris
Dommanget JL (2010) Complément à l’inventaire des libellules de France (Programme national de collecte des données odonatologique (Programme CILIF), Actualisation janvier 2010. Société française d’odonatologie http://www.libellules.org/protocole/cilif.html/. Accessed 15 June 2012
European Environment Agency (2007) Halting the loss of biodiversity by 2010: proposal for a first set of indicators to monitor progress in Europe. EEA, Copenhagen
European Union (2011) Communication from the Commission to the European Parliament, the Council, the Economic and Social Committee and the Committee of the Regions. Our life insurance, our natural capital: an EU biodiversity strategy to 2020. COM(2011) 244 final, Brussels
Gregory RD, van Strien A, Vorisek P, Gmelig Meyling AW, Noble DG, Foppen RPB, Gibbons DW (2005) Developing indicators for European birds. Philos Trans R Soc B 360:269–288
Hassall C (2012) Predicting the distributions of under-recorded Odonata using species distribution models. Insect Conserv Divers 5:192–201
Hassall C, Thompson DJ (2010) Accounting for recorder effort in the detection of range shifts from historical data. Methods Ecol Evol 1:343–350
Hickling R, Roy DB, Hill JK, Thomas CD (2005) A northward shift of range margins in British Odonata. Glob Chang Biol 11:502–506
Kalkman VJ, Boudot JP, Bernard R, Conze KJ, de Knijf G, Dyatlova E, Ferreira S, Jović M, Ott J, Riservato E, Sahlén G (2010) European red list of dragonflies. Publications Office of the European Union, Luxembourg
Kendall WL, White GC (2009) A cautionary note on substituting spatial subunits for repeated temporal sampling in studies of site occupancy. J Appl Ecol 46:1182–1188
Kéry M (2010) Introduction to WinBUGS for ecologists: a Bayesian approach to regression, ANOVA, mixed models and related analyses. Academic Press, Burlington
Kéry M (2011) Towards the modelling of true species distributions. J Biogeogr 38:617–618
Kéry M, Royle JA, Schmid H, Schaub M, Volet B, Häfliger G, Zbinden N (2010) Site-occupancy distribution modeling to correct population-trend estimates derived from opportunistic observations. Conserv Biol 24:1388–1397
MacKenzie DI, Nichols JD, Royle JA, Pollock KH, Hines JE, Bailey LL (2006) Occupancy estimation and modeling: inferring patterns and dynamics of species occurrence. Elsevier, San Diego
Nelson B, Ronayne C, Thompson R (2011) Ireland Red List No. 6. Damselflies & Dragonflies (Odonata). National Parks and Wildlife Service, Department of the Environment, Heritage and Local Government, Dublin
Plummer M (2009) JAGS Version 1.0.3 manual. http://sourceforge.net/projects/mcmc-jags/. Accessed 15 June 2012
Robertson MP, Cumming GS, Erasmus BFN (2010) Getting the most out of atlas data. Divers Distrib 16:363–375
Rota CT, Fletcher RJ, Dorazio RM, Betts MG (2009) Occupancy estimation and the closure assumption. J Appl Ecol 46:1173–1181
Royle JA, Dorazio RM (2008) Hierarchical modeling and inference in ecology. Academic Press, Amsterdam
Royle JA, Kéry M (2007) A Bayesian state-space formulation of dynamic occupancy models. Ecol 88:1813–1823
Schmeller DS, Henry PY, Julliard R, Gruber B, Clobert J, Dziock F, Lengyel S, Nowicki P, Déri E, Budrys E, Kull T, Tali K, Bauch B, Settele J, van Swaay C, Kobler A, Babij V, Papastergiadou E, Henle K (2009) Advantages of volunteer-based biodiversity monitoring in Europe. Conserv Biol 23:307–316
Service de l’observation et des statistiques (2009) Évolution de la qualité des cours d’eau: volet macropolluants. Études et documents, SOeS et Commissariat général au développement durable, Orléans
Service de l’observation et des statistiques (2010) L’environnement en France—edition 2010. Références, SOeS et Commissariat général au développement durable, Orléans
Spiegelhalter D, Thomas A, Best NG (2003) WinBUGS user manual, version 1.4. MCR Biostatistics Unit, Cambridge
Szabo JK, Vesk PA, Baxter PWJ, Possingham HP (2010) Regional avian species declines estimated from volunteer-collected long-term data using list length analysis. Ecol Appl 20:2157–2169
Van Strien AJ, Termaat T, Groenendijk D, Mensing V, Kéry M (2010) Site-occupancy models offer new opportunities for dragonfly monitoring based on daily species lists. Basic Appl Ecol 11:495–503
Van Strien AJ, van Swaay CAM, Kéry M (2011) Metapopulation dynamics in the butterfly Hipparchia semele changed decades before occupancy declined in The Netherlands. Ecol Appl 21:2510–2520
Van Swaay CAM, Plate CL, van Strien A (2002) Monitoring butterflies in the Netherlands: how to get unbiased indices. Proc Exper Appl Entomol NEV Amsterdam 13:21–27
Van Swaay CAM, Nowicki P, Settele J, van Strien AJ (2008) Butterfly monitoring in Europe: methods, applications. Biodivers Conserv 17:3455–3469
Ward L, Mill PJ (2004) Distribution of the Banded Demoiselle Calopteryx splendens (Harris) in northern England: an example of range expansion. J Br Dragonfly Soc 20:61–69
Yoccoz NG, Nichols JD, Boulinier T (2001) Monitoring of biodiversity in space and time. TREE 16:446–453
Acknowledgments
The work of AJvS is facilitated by the BiG Grid infrastructure for eScience (www.biggrid.nl). The DragonflyIreland project (2000–2003) was supported financially by National Parks and Wildlife Service, Northern Ireland Environment Agency and the National Museums Northern Ireland. SP’s post is funded by Countryside Council for Wales, Environment Agency, Esmeé Fairbairn Foundation, Natural England and Scottish Natural Heritage. The work of CV is supported by ARCir BioImpact (Arrêté 11000922). We are grateful to Grégory Motte for organising field work in Wallonia and to Willy van Strien for critically reading the draft and to Rita Gircour for improving the English. Marcel Straver helped in preparing Fig. 2. Finally, we thank all contributors to the dragonfly recording databases in the each country.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
van Strien, A.J., Termaat, T., Kalkman, V. et al. Occupancy modelling as a new approach to assess supranational trends using opportunistic data: a pilot study for the damselfly Calopteryx splendens . Biodivers Conserv 22, 673–686 (2013). https://doi.org/10.1007/s10531-013-0436-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10531-013-0436-1