
Abstract
The factors shaping the ways in which animals use resources are a key element of conservation biology, but ecological studies on resource use typically neglect to consider how the study’s spatial scale may have affected the outcomes. We used the dryad butterfly, inhabiting xerothermic grassland and wet meadow, to test for differences in its resource use at two scales–habitat patch and landscape. Based on records of plant species composition from random points within four habitat patches and from points in 53 patches along surveyed transects, we compared the microhabitat preferences of the butterfly on the patch scale, and species occurrence and abundance patterns on the landscape scale. We distinguished four main groups of factors related to vegetation structure which affected the butterfly’s resource use—factors having similar effects on both spatial scales, factors operating primarily on one of the scales considered, factors relevant only on a single spatial scale, and factors operating on both scales but with effects differing between the two habitat types. We suggest that invertebrates may respond on two spatial levels or on only one, and conclude that larger-scale studies can meet the challenges of a sophisticated metapopulation approach and can give insight into the habitat characteristics affecting the persistence of species in landscapes. We stress the value of large-scale studies on species’ habitat preferences when planning conservation strategies, while pointing out that small-scale studies provide useful information about species ecology and behavior, especially if conducted in multiple habitats.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Assessing the key factors shaping resource use by species is a basic task of conservation biology (WallisDeVries et al. 2002; Krauss et al. 2010; Driscoll et al. 2013). Scale is a unifying concept in ecology, and many ecological processes are scale-dependent (Steffan-Dewenter et al. 2002; Battisti and Fanelli 2015). The majority of ecological studies can be classified according to their spatial scale. Studies on resource use or habitat use by animals can be carried out on the habitat patch scale (e.g., Cobbold and Supp 2012; Strausz et al. 2012; Wajnberg et al. 2013; Ginane et al. 2015), landscape scale (Cozzi et al. 2008; Bergerot et al. 2011) or the much larger scale of the distribution range of a species (Hughes et al. 2003; Lindman et al. 2015). These different scales require the use of various methods, such as observation of individual behavior (Slamova et al. 2011), transect counts (Kalarus and Nowicki 2015), mark-recapture (Schneider et al. 2003; Akeboshi et al. 2015), GPS positioning (Calenge et al. 2009) or mathematical modelling (Wajnberg et al. 2013). In the landscape approach, many matrices reflecting landscape heterogeneity, landscape fragmentation and landscape connectivity can be employed (Símová and Gdulová 2012). The results obtained with these methods give insight into different aspects of animals’ habitat requirements, from individual preferences to the population and metapopulation levels, and further to the drivers of a species distribution range.
The factors behind resource use vary with the habitat type or studied fraction of it, and they are influenced by random parameters such as weather or the time of day (Dennis and Sparks 2006; Slamova et al. 2011; Wajnberg et al. 2013; Botham et al. 2015). The probability of detecting these drivers also depend on the statistical methods and approaches applied (Schippers et al. 2015; Boyce et al. 2016). Crucial preferences and resource use patterns of an investigated species may be not detected in studies conducted on a given scale. The larger landscape scale can elucidate metapopulation processes but tends to overlook specific habitat preferences of individuals (cf. Cozzi et al. 2008; Akeboshi et al. 2015; Schippers et al. 2015). On the other hand, patch-scale research reveals these specific individual preferences and can help in the planning of local conservation measures for endangered species in focal habitat patches, while leaving one in the dark about the drivers of metapopulation dynamics (cf. Dennis and Sparks 2006; Slamova et al. 2011). In extreme cases, even good conservation work on the patch scale may not prevent the extinction of local populations due to obstruction of dispersal processes or declining genetic variation (Driscoll et al. 2013).
In our study we used the dryad butterfly (Minois dryas) to investigate how the spatial scale of research affects findings on resource use, and we assessed the relevance of these findings for the design of conservation programmes. The dryad inhabits two contrasting habitat types: xerothermic grassland and wet meadow (Buszko and Masłowski 2008; Kalarus and Nowicki 2015). The dryad is a good model for studies of habitat use and species colonization processes, as its distribution has recently been spreading in Poland. As butterflies strongly depend on plant species, used by adults as nectar sources or as food plants by larvae, we took vegetation composition as a predictor of habitat preference in this group. We tested the effects of vegetation characteristics on resource use by the dryad on two spatial scales: habitat patch and landscape. We wanted to determine which effects can be detected on the two scales, and which are specific to a particular scale.
Methods
Study area
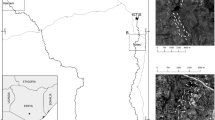
The study was carried out within an extensive grassland complex covering ca. 35 km2, about 8 km south-west of the centre of the town of Kraków in southern Poland. The area is part of the Bielańsko-Tyniecki Landscape Park and the Dębnicko-Tyniecki Obszar Łąkowy Natura 2000 Area (PLH 120,065). The grasslands form a habitat mosaic of wet meadow patches in the Vistula River valley and xerothermic calcareous grassland patches on hills. The xerothermic grasslands comprise vegetation of the Festuco-Brometea class and thermophilous sandy grasslands of the Festuco-Thymetum serpylli vegetation. The wet meadows are covered with vegetation of the Molinietalia order, mainly Molinion communities of different quality, and relatively humid lowland hay meadows (Kalarus and Nowicki 2015). The landscape-scale study was done in 27 patches of xerothermic grassland and 26 patches of wet meadow within the grassland complex (Fig. 1, Electronic Supplementary Material (ESM) S1). Patch area ranged from 0.066 to 11.749 ha for wet meadow, and from 0.024 to 3.452 ha for xerothermic grassland. The typical distance between the nearest neighboring patches ranged from 100 to 600 m. To examine the influence of vegetation on the dryad’s resource use on the patch scale we selected two xerothermic grasslands and two wet meadows (included also in the landscape-scale study) in the core fragment of the species distribution area, in the western part of the meadow complex (Fig. 1, ESM S1). These patches covered between 0.813 and 4.184 ha and were from 100 to 400 m apart.

Location and spatial structure of the study area in the Kraków region, southern Poland. Light grey xerothermic grassland patches; dark grey wet meadow patches; bolded boundaries—patches occupied by the dryad. Four patches used in the patch-scale study are outlined. Data are presented in the EPSG coordinate reference system (32,634 – WGS 84 / UTM zone 34 N)
Study species
The dryad butterfly has a Euro-Siberian distribution range from the northern part of the Iberian Peninsula to Japan (Dąbrowski 1999; Kudrna 2002). In Europe it usually inhabits two contrasting habitat types: Molinietalia wet meadow and xerothermic grassland with steppe plants typical for communities of Festuco-Brometea (Buszko and Masłowski 2008). The dryad occurs usually locally in xerothermic and wet habitats. In Poland its occurrence has been restricted to the southern part of the country in the Kraków region and the Eastern Carpathians, although recently the species is spreading in the country (Buszko and Masłowski 2008; Warecki and Sielezniew 2008). The dryad is protected under Polish law and listed as a critically endangered species in the Polish Red Data Book of Animals (Głowaciński and Nowacki 2004). The larval food plants include Poaceae grasses mainly Molinia caerulea as well as Festuca rubra, Bromus erectus, Calamagrostis epigeios and Arrhenatherum elatius. Females do not show any distinct preferences for ovipositing on specific foodplants and drop their eggs on various grasses (Settele et al. 1999; Głowaciński and Nowacki 2004; Buszko and Masłowski 2008). The flight period is from the end of July to the beginning of September. With its expanding distribution, the dryad is a good model for studies of species expansion and colonization processes, as in the Kraków region, where its distribution has recently been spreading from a former refugium—the Skołczanka Reserve. Further advantages are that the dryad is a relatively large butterfly, and its flight is not very fast and easy to follow, which makes it a useful object for habitat use studies and investigations using transect counts and mark-recapture methods.
Field study
Data collection was done in four patches in August of 2009 and 2010 for the patch-scale study (Fig. 1), and between July and early September of 2013 for the landscape-scale study spanning the entire meadow complex (Fig. 1). Butterfly observations were carried out on sunny and windless days. Habitat patch quality and vegetation composition were characterized at random points in each investigated habitat patch. For each random point, five measurements of vegetation height were taken with a measuring tape to 1 cm accuracy: one measurement at the point and the other four 1.5 m to the north, south, east and west of it. Distance to the nearest shrub was measured to 10 cm accuracy in order to investigate the butterfly preferences for places near to shrubs for patch-scale study and to test effect of shrub density for landscape-scale study. The shrub density was approximated as the inverse distance to the nearest shrub and expressed in m−1. Finally, all flowering nectar plant species within a 1.5 m radius of the point were counted and their ground cover was recorded. Cover was scored on a 10-point scale: 1 = < 10%; 2 = 11–20%, ..., 9 = 81–90% and 10 = 91–100%. Special attention was given to invasive goldenrod cover, which was adopted as a measure of vegetation succession and habitat deterioration. The goldenrods Solidago gigantea and S. canadensis are alien species overgrowing abandoned meadows in Europe (Scharfy et al. 2010). For further details of the studies see Kalarus et al. (2013) and Kalarus and Nowicki (2015).
Landscape scale field sampling
For the landscape-scale study the vegetation was surveyed at 100 random points for xerothermic grassland patches and 110 points for wet meadow patches, once at each point. At each patch 3–10 points were located at a random distance along transects, the length of which varied between 50 and 500 m, depending on patch size. In all habitat patches we recorded the presence and number of dryads along 5 m wide transects, using the standard Pollard walk method (Pollard and Yates 1993). Detectability of the dryad butterfly was estimated using the occupancy model of MacKenzie et al. (2002); detectability was very close to 1, so we can safely assume that the recorded presence-absence pattern represents the true pattern of patch occupancy by the species (see Kalarus and Nowicki 2015 for more details). We made three counts of the dryads along transects in each meadow and recorded dryad abundance at occupied patches between July 28 and August 24. The dryad abundance was expressed as the total number of individuals for three counts calculated per 1 ha of transect.
Patch scale field sampling
In the patch-scale study the points within patches were selected through random generation of their geographic coordinates. The vegetation was investigated at 32 random points for xerothermic grasslands and 31 points for wet meadows. Apart from this, at all the investigated patches we selected the dryad occurrence points for studying their microhabitat preferences by following randomly spotted flying imagoes between August 1 and August 21. The butterfly observations were only conducted on the days with appropriate weather, namely sunny and windless conditions. In xerothermic grassland 39 individuals and in wet meadows 34 individuals were followed. In both habitat types relatively balanced numbers of females and males were recorded, with only a slight majority of females. The places where they landed on plants were marked with a bamboo pole. Subsequently, we measured and recorded the vegetation within a 1.5 m radius of those places in the same way as for the random points. It should be noted that for the subsequent analysis, the data for males and females were pooled together to increase the sample size, as there was no difference in the microhabitat use by both sexes (Kalarus et al. 2013).
Statistical analysis
To describe the diversity of plant species composition within habitat patches, and to detect the most important gradients of this diversity, we applied non-metric multidimensional scaling (NMS) based on Bray-Curtis distances using Canoco for Windows 4.52 (Lepš and Šmilauer 2003; Kindt and Coe 2005), separately for xerothermic grassland and wet meadow patches. The NMS allows seeking for a configuration of sample points that best reflect the rank of inter-sample distances, using other distance measure than Euclidean distances. This approach is highly useful if differences between sample sizes occur, because it avoids the deficiency of Euclidean distances that overestimate the importance of the most abundant plant species (Lepš and Šmilauer 2003; Kindt and Coe 2005). Finally, the NMS method enables to find subtle differences in plant species assemblages at distinguished gradients. The NMS analysis was performed in four dimensions. For testing the differences between groups of points we performed ANOSIM analysis (see Results). The analysis was performed in PAST 3.01 (Hammer et al. 2001).
Landscape scale
At the landscape scale the random points established along the transects on both occupied and vacant patches were used for NMS analysis. Final stress reached 0.129 for xerothermic grassland patches and 0.151 for wet meadow patches, which reflects the high reliability of the analyses. The data concerning butterflies occupancy and abundance patterns were analysed using multimodel selection and inference procedures in the MuMIn package of R software (Burnham and Anderson 2002; R Core Team 2013). In the present work we only considered the effects of the following independent variables: vegetation height, shrub density, goldenrod cover, nectar plants cover and habitat type. Nevertheless, the analysis also included further factors in order to control for their effects even if they are not the subject of interest here (see ESM S1 and Kalarus and Nowicki 2015). Logistic regression modeling with binomial distributions of the dependent variable was used to build the models of dryad occupancy patterns; to build the models of butterfly abundance patterns we used the general linear model (GLM) approach and Gaussian distributions of the dependent variable. The latter analysis was performed using data obtained for all 18 occupied patches. The dependent variable was cubic-rooted to achieve normality. To find the set of the most supported models among all possible models we applied the Akaike Information Criterion corrected for small sample size (AICc) (Hurvich and Tsai 1989). Next, the model results were averaged across all supported models i.e., those with ΔAICc < 7 (ESM S1), using their Akaike weights, which reflect the probability that a given model is the best one (Burnham and Anderson 2002). All non-significant interaction terms were removed from the final model. As the occupied patches were located in one part of the study area forming a cluster around the former species refugium, an additional analysis of occupancy pattern was performed for a restricted data set of patches located within the cluster as these patches may have distinctively higher chance to be occupied. For more details of the analyses see ESM S1 as well as Kalarus and Nowicki (2015).
Patch scale
At the patch scale the random points as well as butterfly resting and nectaring points were used in NMS as grouping factors for vegetation structure. Final stress was 0.150 for xerothermic grassland patches and 0.153 for wet meadow patches, which again reflects the high reliability of the analyses. To test the effects of shrub density (assessed as the distance to the nearest shrub), vegetation height and the cover of invasive goldenrods on butterfly habitat use, we performed hierarchical ANOVA for the patch-scale data, with random points vs butterfly nectaring and resting places as the main groups. The analyses was conducted using Statistica 9.0. For wet meadow we also calculated the availability of invasive alien goldenrods as well as of native composite plants constituting nectar sources, using the overall cover of all the plants in a given group pooled across all the random points within the habitat type. The proportion of observations of nectaring on plants in both groups (representing their use by the dryad) were compared with their availability, using Bailey’s use-availability test performed in MS Excel 2007 (Bailey 1980).
Results
Both xerothermic grassland and wet meadow had relatively homogeneous plant communities. On the landscape scale plant species composition differed between points located within occupied and vacant patches in both habitat types (ANOSIM: xerothermic grasslands: r = 0.12, P < 0.001; wet meadows: r = 0.04, P = 0.047), whereas on the patch scale butterfly occurrence places differed from random points only for wet meadows (xerothermic: r = 0.03, P = 0.196; wet: r = 0.25, P < 0.001). On both investigated scales, NMS analysis revealed three main ecological gradients of plant species diversity at the random points in the investigated patches. The gradients were identical in both habitat types but differed between the patch and landscape scales. The gradients were identified based on plant species composition in investigated points and on the plant species characteristics, such as preferred environmental conditions, syntaxonomical memberships and invasive traits (ESM S2).
On the landscape scale the gradient related to the first ordination axis explained 35.8% of plant community variation for xerothermic patches (score 0.970), and 38.6% for wet meadow patches (score 0.988). This gradient reflected a shift of the general quality of the plant community from one having a typical species composition to a degraded one, characterized by a larger share of ruderal or invasive plants. Importantly, degraded xerothermic grassland also included plants typically associated with wet meadows or even the garden plants (ESM S2, Fig. 2a, c). The gradient related to the second ordination axis explained 29.0% of plant community variation for xerothermic grassland (score −0.952), and 25.2% for wet meadow (score 0.962). In xerothermic grassland it was linked partly with increasing habitat impoverishment, as suggested by the presence of species deterioration such as Rumex spp. and Agrostis capillaris, and potential host plant of the dryad—Calamagrostis epigeios, that tend to overgrow xerothermic grasslands. In wet meadows it was associated with increasing humidity (ESM S2). The third ordination axis explained 20.1% of the variation of plant species composition for xerothermic grassland (score 0.980), and 19.5% for wet meadow (score 0.980). The gradient associated with this axis reflected increasing grass cover and vegetation height (ESM S2, Fig. 2 a, c).

Non-metric multidimensional scaling (NMS) ordination of points within habitat patches, indicating plant species composition on the landscape scale (a, c) and patch scale (b, d) for xerothermic grassland (a, b), and wet meadow (c, d). The ordination diagram represents the first and third ordination axes. “GR” (with a number) denotes a gradient related with respective axis. Landscape scale: filled squares represent points within occupied patches, empty squares represent points within unoccupied patches. Patch scale: filled squares represent butterfly resting places, gray circles represent butterfly nectaring places, and empty squares represent random points within habitat patches. See text for explanation of ordination axes
At patch scale the gradient related to the first ordination axis explained 34.3% of plant community variation for xerothermic grassland (score 0.996), and 35.1% for wet meadow (score 0.994). It reflected the diversity and abundance of the nectar plants preferred by the dryad. (ESM S2, Fig. 2 b, d). The second gradient accounted for 26.9% of plant community variation for xerothermic grassland (score −0.984), and 25.9% for wet meadow (score 0.976). For xerothermic grassland this ordination axis was positively related to higher microhabitat temperature and negatively related to humidity (ESM S2). For wet meadow it was associated with habitat deterioration, reflected in higher abundance of Solidago gigantea and lower abundance of various nectar plants and grasses other than Molinia caerulea (ESM S2). The third ordination axis explained 20.5% of the variation of plant species composition for xerothermic grassland (score −0.901), and 20.1% for wet meadow patches (score −0.708). It reflected the decreasing quality of the microhabitats’ plant community, from a more typical to degraded species composition (ESM S2, Fig. 2 b, d).
Shrub density was a crucial factor influencing habitat use by the dryad at patch scale, while at landscape scale it was of less importance (Fig. 3 a, b) and its effect on the species occurrence was detected only for a few patches in the west part of the meadow complex for the dryad occurrence pattern. At patch scale, vegetation height was marginally significant in xerothermic grassland, where butterflies tended to prefer places with higher vegetation. At landscape scale this factor proved non-significant (Fig. 3 c, d). The negative influence of the invasive goldenrod was significant at both scales. Butterflies avoided nectaring on the goldenrod at patch scale, while at landscape scale the negative effect of this alien plant was detected for the dryad abundance pattern (Fig. 4).

Standardized model estimates with 95% confidence intervals based on multi-model inference (for landscape scale) and hierarchical ANOVA (for patch scale) for the key variables explaining the dryad’s habitat use: density of shrubs (a, b) [approximated as the inverse distance to the nearest shrub and expressed in m−1 (a), distance to shrub with reference to random points in m (b)] and vegetation height (cm) (c, d). For patch scale, the estimates for random points group are shown. Occupancy Asterisk denotes the result for data excluding vacant patches distant from a cluster of occupied ones. A particular factor is significant if its estimated confidence intervals do not overlap zero. For plates (a, b) and c positive values denote positive effects e.g., preference of butterfly for places close to shrubs, while for plate d negative values denote positive effects e.g., preference of butterflies for places with higher vegetation on xerothermic grassland

The impact of the invasive goldenrod on the habitat use by the dryad at landscape (a) and patch scale (b). For the landscape scale, the diamonds show standardized model estimates for the goldenrod cover with their 95% confidence intervals based on multi-model inference, and the effect of goldenrod should be considered significant if the confidence intervals do not overlap zero. For the patch scale, the diamond indicates proportional use of goldenrods by the dryad, presented with its Bailey’s 95% confidence interval. The goldenrod availability (broken line) shown for comparison is well above this 95% confidence interval, which denotes statistically significant avoidance
Discussion
In this study the patterns of resource use by the dryad were mostly scale-dependent. The data permitted us to distinguish four groups of environmental factors affecting those patterns. The first group consists of factors of great importance on one of the spatial scales but unimportant on the other scale as revealed predominantly by the first gradient of the NMS ordination. At patch scale this gradient reflected diversity and abundance of nectar plants, which suggests that the share of nectar plants mainly drives the dryad’s microhabitat use at patch scale. Plant community quality affects the dryad mainly at landscape scale. The second group comprises factors whose effects are highly significant on both scales and which operate in the same direction, such as invasive goldenrod cover (with a negative effect) and density of shrubs (with a positive effect). The third group includes factors of some importance only at patch scale, whose effects differ between the two habitat types; in the case of vegetation height, for example, the dryad showed a preference for microhabitats with higher vegetation, but only in xerothermic grassland. The fourth group contains factors of lower importance at both scales and whose effects differ between the two habitat types, such as habitat deterioration and humidity. We found no factor whose effects on the dryad’s resource use differed in direction between scales.
Patch-scale studies have a smaller selection of research approaches to choose from. An obvious limitation is the impossibility of testing the effects of spatial characteristics of habitat patches (e.g., their size and shape) or the effects of the surrounding matrix on resource use by focal species (e.g., Strausz et al. 2012). Broader landscape-scale research can elaborate sophisticated metapopulation models to account for matrix impacts (Shreeve and Dennis 2011; Driscoll et al. 2013). Patch-scale research can only reveal the preferences of taxa under the delimited conditions of particular habitat patches (e.g., Slamova et al. 2011). Consequently, it cannot shed light on the general patterns of resource use and may draw attention away from its most important drivers. The failure of some conservation strategies in the past can be attributed to excessive reliance on strong conclusions drawn from patch-scale research (cf. Godet et al. 2007; Battisti and Fanelli 2015).
On a larger scale it is more likely that patterns difficult or even impossible to detect on patch scale, such as the effects of plant community quality and grass cover, will be detected. A recent study by Botham et al. (2015) showed that the same habitat composition differently affects specialist and generalist butterfly species. Landscape-scale research also provides information on whether a given species exists in a single unfragmented population, a patchy population, or a meta-population system (Battisti and Fanelli 2015). This cannot be determined on a local scale. The results obtained from landscape-scale studies are much more useful for predicting the long-term persistence of a species (Godet et al. 2007; Battisti and Fanelli 2015).
On the other hand, a recent study by Gonthier et al. (2014) indicated that various taxonomic groups respond differently to the factors operating on small and landscape scales. Species with low mobility turned out to be affected mainly on the local scale, while species with good dispersion ability are affected mainly at the landscape level. Certain factors affecting the dryad’s resource use turned out to have similar effects on both investigated scales. Some invertebrates respond similarly to factors operating on both spatial scales, and some respond more on one scale (cf. Steffan-Dewenter et al. 2002).
To improve the explanatory power of patch-scale studies, we suggest that a multi-model approach should be used to determine the relative importance of the assessed variables (Burnham and Anderson 2002), and that these studies should include sites with different habitat conditions (cf. Pellet et al. 2012; Boyce et al. 2016). In practical terms, if the relative importance of a significant factor is very high in patches of different habitat types, that factor can be expected to drive species preferences on the landscape scale as well. In our study such factors were alien goldenrod cover and shrub density.
Although we suggest that conservation planning for target species should be based primarily on the results of landscape-scale studies, some specific measures based on the results of patch-scale studies may be worth implementing in given habitat patches if they are particularly important for metapopulation functioning, for example through improvement of connectivity (Hanski 1999; Krauss et al. 2010). Generally, we strongly recommend a multiscale approach. Results obtained simultaneously from local patch-scale and landscape-scale studies are the most robust. The conclusions drawn from them can serve as a sound basis for the design of measures aimed at conserving target species and biodiversity.
References
Akeboshi A, Takagi S, Murakami M, Hasegawa M, Miyashita T (2015) A forest–grassland boundary enhances patch quality for a grassland-dwelling butterfly as revealed by dispersal processes. J Insect Conserv 19:15–24
Bailey BJR (1980) Large sample simultaneous confidence intervals for the multinomial probabilities based on transformations of the cell frequencies. Technometrics 22:583–589
Battisti C, Fanelli G (2015) Don’t think local! scale in conservation parochialism dogmatic bureaucracy and the implementing of the European Directives. J Nat Conserv 24:24–30
Bergerot B, Fontaine B, Julliard R, Baguette M (2011) Landscape variables impact the structure and composition of butterfly assemblages along an urbanization gradient. Landsc Ecol 26:83–94
Botham MS, Fernandez-Ploquin EC, Brereton T, Harrower CA, Roy DB, Heard MS (2015) Lepidoptera communities across an agricultural gradient: how important are habitat area and habitat diversity in supporting high diversity? J Insect Conserv 19:403–420
Boyce MS, Johnson CJ, Merrill EH, Nielsen SE, Solberg EJ, Van Moorter B (2016) Can habitat selection predict abundance? J Anim Ecol 85:11–20
Burnham KP, Anderson DR (2002) Model selection and multimodel inference. Springer, New York
Buszko J, Masłowski J (2008) Motyle dzienne Polski. Koliber, Nowy Sącz (in Polish)
Calenge C, Dray S, Royer-Carenzi M (2009) The concept of animals’ trajectories from a data analysis perspective. Ecol Inform 4:34–41
Cobbold SM, Supp SR (2012) Patch shape alters spider community structure: links between microhabitat choice and sensitivity to increased edge habitat. J Insect Conserv 16:581–589
Cozzi G, Müller CB, Krauss J (2008) How do local habitat management and landscape structure at different spatial scales affect fritillary butterfly distribution on fragmented wetlands? Landsc Ecol 23:269–283
Dąbrowski JS (1999) Skalnik driada Minois dryas (Scop.), (Lepidoptera: Satyridae) – gatunek zagrożony wyginięciem na ostatnich znanych stanowiskach w Polsce. Chrońmy Przyrodę Ojczystą 55:91–94 (in Polish)
Dennis RLH, Sparks TH (2006) When is a habitat not a habitat? Dramatic resource use changes under differing weather conditions for the butterfly Plebejus argus. Biol Conserv 129:291–301
Driscoll DA, Banks SC, Barton PS, Lindenmayer DB, Smith AL (2013) Conceptual domain of the matrix in fragmented landscapes. Trends Ecol Evol 28:605–613
Ginane C, Manuelian CL, Dumont B (2015) Sheep herbivory within grassland patches: the potential cost of food item discrimination. Basic Appl Ecol 16:347–353
Głowaciński Z, Nowacki J (eds) (2004) Polska czerwona księga zwierząt. Instytut Ochrony Przyrody PAN, Kraków, Akademia Rolnicza, Poznań (in Polish with English abstract)
Godet L, Devictor V, Jiguet F (2007) Estimating relative population size included within protected areas. Biodivers Conserv 16:2587–2598
Gonthier DJ, Ennis KK, Farinas S, Hsieh H, Iverson AL, Batáry P, Rudolphi J, Tscharntke T, Cardinale BJ, Perfecto I (2014) Biodiversity conservation in agriculture requires a multi-scale approach. Proc R Soc Lond B 281:20141358
Hammer Ø, Harper DAT, Ryan PD (2001) PAST: Paleontological statistics software package for education and data analysis. Palaeontologia Electronica 4(1):9. http://palaeoelectronicaorg/2001_1/past/issue1_01htm
Hanski I (1999) Metapopulation ecology. Oxford University Press, New York
Hughes CL, Hill JK, Dytham C (2003) Evolutionary trade-offs between reproduction and dispersal in populations at expanding range boundaries. Proc R Soc Lond B 270:147–150
Hurvich CM, Tsai C (1989) Regression and time series model selection in small samples. Biometrika 76:297–307
Kalarus K, Nowicki P (2015) How do landscape structure, management and habitat quality drive the colonization of habitat patches by the dryad butterfly (Lepidoptera: Satyrinae) in fragmented grassland? PLoS One 10:e0138557
Kalarus K, Skórka P, Nowicki P (2013) Resource use in two contrasting habitat types raises different conservation challenges for the conservation of the dryad butterfly Minois dryas. J Insect Conserv 17:777–786
Kindt R, Coe R (2005) Tree diversity analysis, a manual and software for common statistical methods for ecological and biodiversity studies. World Agroforestry Centre (ICRAF), Nairobi
Krauss J, Bommarco R, Guardiola M, Heikkinen RK, Helm A, Kuussaari M, Lindborg R, Öckinger E, Pärtel M, Pino J, Pöyry J, Raatikainen KM, Sang A, Stefanescu C, Teder T, Zobel M, Steffan-Dewenter I (2010) Habitat fragmentation causes immediate and time-delayed biodiversity loss at different trophic levels. Ecol Lett 13:597–605
Kudrna O (2002) The distribution atlas of European butterflies. Oedippus 20:1–343
Lepš J, Šmilauer P (2003) Multivariate analysis of ecological data using CANOCO. Cambridge University Press, New York
Lindman L, Remm J, Saksing K, Sõber V, Õunap E, Tammaru T (2015) Lycaena dispar on its northern distribution limit: an expansive generalist. Insect Conserv Divers 8:3–16
MacKenzie DI, Nichols JD, Lachman GB, Droege S, Royle JA, Langtimm CA (2002) Estimating site occupancy rates when detection probabilities are less than one. Ecology 83:2248–2255
Pellet J, Bried JT, Parietti D, Gander A, Heer PO, Cherix D, Arlettaz R (2012) Monitoring butterfly abundance: beyond Pollard Walks. PLoS One 7:e41396
Pollard E, Yates TJ (1993) Monitoring butterflies for ecology and conservation. Chapman & Hall, London
R Core Team (2013) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.r-project.org/
Scharfy D, Güsewell S, Gessner MO, Venterink HO (2010) Invasion of Solidago gigantea in contrasting experimental plant communities: effects on soil microbes nutrients and plant–soil feedbacks. J Ecol 98:1379–1388
Schippers P, Hemerik L, Baveco JM, Verboom J (2015) Rapid diversity loss of competing animal species in well-connected landscapes. PLoS One 10:e0132383
Schneider C, Dover J, Fry GLA (2003) Movement of two grassland butterflies in the same habitat network: the role of adult resources and size of the study area. Ecol Entomol 28:219–227
Settele J, Feldmann R, Reinhardt R (eds) (1999) Die tagfalter deutschlands. ulmer, Stuttgart
Shreeve TG, Dennis RLH (2011) Landscape scale conservation: resources behaviour the matrix and opportunities. J Insect Conserv 15:179–188
Símová P, Gdulová K (2012) Landscape indices behavior: a review of scale effects. Appl Geogr 34:385–394
Slamova I, Klecka J, Konvicka M (2011) Diurnal behavior and habitat preferences of Erebia aethiops an aberrant lowland species of a mountain butterfly clade. J Insect Behav 24:230–246
Steffan-Dewenter I, Münzenberg U, Bürger C, Thies C, Tscharntke T (2002) Scale-dependent effects of landscape context on three pollinator guilds. Ecology 83:1421–1432
Strausz M, Fiedler K, Franzén M, Wiemers M (2012) Habitat and host plant use of the large copper butterfly Lycaena dispar in an urban environment. J Insect Conserv 16:709–721
Wajnberg E, Hoffmeister TS, Coquillard P (2013) Optimal within-patch movement strategies for optimizing patch residence time: an agent-based modelling approach. Behav Ecol Sociobiol 67:2053–2063
WallisDeVries MF, Poschlod P, Willems JH (2002) Challenges for the conservation of calcareous grasslands in northwestern Europe: integrating the requirements of flora and fauna. Biol Conserv 104:265–273
Warecki A, Sielezniew M (2008) Dryad Minois dryas (Lepidoptera, Nymphalidae) in south-eastern Poland: a recent range expansion or oversight of an endangered species? Pol J Entomol 77:191–198
Acknowledgements
The study was funded by a Polish National Science Centre grant DEC-2013/11/B/NZ8/00912. The fieldwork was additionally supported by the Jagiellonian University through its statutory funds (DS/WBINOZ/INOŚ/761/12–13) and by the Bratniak Foundation. Michael Jacobs line-edited the paper for submission.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Kalarus, K., Nowicki, P. Resource use by the dryad butterfly is scale-dependent. Popul Ecol 59, 179–187 (2017). https://doi.org/10.1007/s10144-017-0579-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10144-017-0579-0