
Abstract
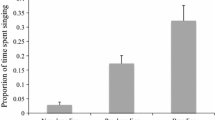
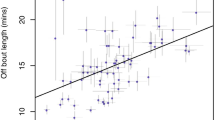
Extra-pair fertilizations are common in many socially monogamous species, and paternity studies have indicated that females may use male vocal performance and plumage ornaments as cues to assess male quality. Female off-territory forays may represent a key component of female choice and male extra-pair mating success, and female foray behaviour is expected to be strongly influenced by indictors of male quality. In this study, we examined how male song and ornamentation affect how often females left their territories, which males they visited and extra-pair paternity in a socially monogamous passerine, the hooded warbler (Wilsonia citrina). We radiotracked 17 females during the fertile period and quantified male vocal performance (song output and rate) and plumage characteristics (size of the black melanin hood and colour of the black hood, yellow cheeks and breast areas). We obtained blood samples and determined paternity at 35 nests including those of 14 females that we radiotracked. Eleven (65%) of the 17 females forayed off-territory, whilst fertile and female foray rate was positively correlated with the number of extra-pair young in the nest. Females that left their territories more frequently were paired with males that sang at a low rate. In addition, extra-pair mates had higher song rates than the social mates they cuckolded (5.3 songs/min vs. 4.4 songs/min). Female off-territory forays or extra-pair paternity were not significantly related to male plumage characteristics. Our results indicate that a high song rate influences both the foray behaviour of a male’s social mate and the likelihood that he will sire extra-pair offspring with neighbouring females.



Similar content being viewed by others

References
Albrecht T, Schnitzer J, Kreisinger J, Exnerova A, Bryja J, Munclinger P (2007) Extrapair paternity and the opportunity for sexual selection in long-distant migratory passerines. Behav Ecol 18:477–486
Ballentine B (2006) Morphological adaptation influences the evolution of a mating signal. Evolution 60:1936–1944
Byers BE (2007) Extrapair paternity in chestnut-sided warblers is correlated with consistent vocal performance. Behav Ecol 18:130–136
Chakraborty R, Meagher TR, Smouse PE (1988) Parentage analysis with genetic markers in natural populations I. The expected proportion of offspring with unambiguous paternity. Genetics 118:527–536
Dawson RJG, Gibbs HL, Hobson KA, Yezerinac SM (1997) Isolation of microsatellite DNA markers from a passerine bird, Dendroica petechia (the yellow warbler), and their use in population studies. Heredity 79:506–514
Dickinson JL (1997) Male detention affects extra-pair copulation frequency and pair behaviour in western bluebirds. Anim Behav 53:561–571
Dolan AC, Murphy MT, Redmond LJ, Sexton K, Duffield D (2007) Extrapair paternity and the opportunity for sexual selection in a socially monogamous passerine. Behav Ecol 18:985–993
Double M, Cockburn A (2000) Pre-dawn infidelity: females control extra-pair mating in superb fairy-wrens. Proc R Soc Lond, B Biol Sci 267:465–470
Dunn PO, Whittingham LA (2007) Search costs influence the spatial distribution, but not the level, of extra-pair mating in tree swallows. Behav Ecol Sociobiol 61:449–454
Endler JA (1990) On the measurement and classification of color in studies of animal color patterns. Biol J Linn Soc 41:315–352
Fedy BC, Norris DR, Stutchbury BJM (2002) Do male hooded warblers guard their mates when their paternity is most at risk? J Field Ornithol 73:420–426
Forstmeier W, Kempenaers B, Meyer A, Leisler B (2002) A novel song parameter correlates with extra-pair paternity and reflects male longevity. Proc R Soc Lond, B Biol Sci 269:1479–1485
Gibson RM, Langen TA (1996) How do animals choose their mates? Trends Ecol Evol 11:468–470
Gil D, Gahr M (2002) The honesty of bird song: multiple constraints for multiple traits. Trends Ecol Evol 17:133–141
Gray EM (1996) Female control of offspring paternity in a western population of red-winged blackbirds (Agelaius phoeniceus). Behav Ecol Sociobiol 38:267–278
Green DJ, Osmond HL, Double MC, Cockburn A (2000) Display rate by male fairy-wrens (Malurus cyaneus) during the fertile period of females has little influence on extra-pair mate choice. Behav Ecol Sociobiol 48:438–446
Griffith SC, Owens IPF, Thuman KA (2002) Extra pair paternity in birds: a review of interspecific variation and adaptive function. Mol Ecol 11:2195–2212
Grill CP, Rush VN (2000) Analysing spectral data: comparison and application of two techniques. Biol J Linn Soc 69:121–138
Hasselquist D, Bensch S, vonSchantz T (1996) Correlation between male song repertoire, extra-pair paternity and offspring survival in the great reed warbler. Nature 381:229–232
Hill GE (2000) Energetic constraints on expression of carotenoid-based plumage coloration. J Avian Biol 31:559–566
Hill GE (2002) A Red Bird in a Brown Bag: The Function and Evolution of Ornamental Plumage Coloration in the House Finch. Oxford University Press, Oxford
Ibarguchi G, Gissing GJ, Gaston AJ, Boag PT, Friesen VL (2004) Male-biased mutation rates and the overestimation of extrapair paternity: Problem, solution, and illustration using thick-billed murres (Uria lomvia, Alcidae). J Heredity 95:209–216
Kempenaers B, Verheyren GR, Dhondt AA (1997) Extrapair paternity in the blue tit (Parus caeruleus): female choice, male characteristics, and offspring quality. Behav Ecol 8:481–492
Lambrechts M, Dhondt AA (1988) The anti-exhaustion hypothesis—a new hypothesis to explain song performance and song switching in the great tit. Anim Behav 36:327–334
Lessells CM, Boag PT (1987) Unrepeatable repeatabilities: a common mistake. Auk 104:116–121
Lifjeld JT, Robertson RJ (1992) Female control of extra-pair fertilization in tree swallows. Behav Ecol Sociobiol 31:89–96
Marshall TC, Slate J, Kruuk LEB, Pemberton JM (1998) Statistical confidence for likelihood-based paternity inference in natural populations. Mol Ecol 7:639–655
Mays HL, Ritchison G (2004) The effect of vegetation density on male mate guarding and extra-territorial forays in the yellow-breasted chat (Icteria virens). Naturwissenschaften 91:195–198
Mays HL, McGraw KJ, Ritchison G, Cooper S, Rush V, Parker RS (2004) Sexual dichromatism in the yellow-breasted chat Icteria virens: spectrophotometric analysis and biochemical basis. J Avian Biol 35:125–134
McGraw KJ, Hill GE (2000) Differential effects of endoparasitism on the expression of carotenoid- and melanin-based ornamental coloration. Proc R Soc Lond Series B Biol Sci 267:1525–1531
McGraw KJ, Beebee MD, Hill GE, Parker RS (2003) Lutein-based plumage coloration in songbirds is a consequence of selective pigment incorporation into feathers. Comp Biochem Physiol Part B Biochem Mol Biol 135:689–696
Morton ES (1989) Female hooded warbler plumage does not become more male-like with age. Wilson Bull 101:460–462
Mota PG, Hoi-Leitner M (2003) Intense extrapair behaviour in a semicolonial passerine does not result in extrapair fertilizations. Anim Behav 66:1019–1026
Neudorf DL, Pitcher TE (1997) Radio transmitters do not affect nestling feeding rates by female hooded warblers. J Field Ornithol 68:64–68
Neudorf DL, Stutchbury BJM, Piper WH (1997) Covert extraterritorial behavior of female hooded warblers. Behav Ecol 8:595–600
Otter K, Chruszcz B, Ratcliffe L (1997) Honest advertisement and song output during the dawn chorus of black-capped chickadees. Behav Ecol 8:167–173
Pedersen MC, Dunn PO, Whittingham LA (2006) Extraterritorial forays are related to a male ornamental trait in the common yellowthroat. Anim Behav 72:479–486
Perreault S, Lemon RE, Kuhnlein U (1997) Patterns and correlates of extrapair paternity in American redstarts (Setophaga ruticilla). Behav Ecol 8:612–621
Podos J (1996) Motor constraints on vocal development in a songbird. Anim Behav 51:1061–1070
Podos J (1997) A performance constraint on the evolution of trilled vocalizations in a songbird family (Passeriformes:Emberizidae). Evolution 51:537–551
Poesel A, Foerster K, Kempenaers B (2001) The dawn song of the blue tit Parus caeruleus and its role in sexual selection. Ethology 107:521–531
Pyle P (1997) Identification guide to North American birds. Part I. Bolinas. Slate Creek Press, California
Seutin G, White BN, Boag PT (1991) Preservation of avian blood and tissue samples for DNA analyses. Can J Zool -Revue Canadienne De Zoologie 69:82–90
Stutchbury BJM (1998) Extra-pair mating effort of male hooded warblers, Wilsonia citrina. Anim Behav 55:553–561
Stutchbury BJ, Neudorf DL (1997) Female control, breeding synchrony and the evolution of extra-pair mating systems. Ornithol Monogr 49:103–129
Stutchbury BJM, Piper WH, Neudorf DL, Tarof SA, Rhymer JM, Fuller G, Fleischer RC (1997) Correlates of extra-pair fertilization success in hooded warblers. Behav Ecol Sociobiol 40:119–126
Stutchbury BJM, Pitcher TE, Norris DR, Tuttle EM, Gonser RA (2005) Does male extra-territory foray effort affect fertilization success in hooded warblers Wilsonia citrine? J Avian Biol 36:471–477
Tarof SA, Ratcliffe LM (2000) Pair formation and copulation behavior in least flycatcher clusters. Condor 102:832–837
Tarof SA, Dunn PO, Whittingham LA (2005) Dual functions of a melanin-based ornament in the common yellowthroat. Proc R Soc B-Biol Sci 272:1121–1127
Thusius KJ, Peterson KA, Dunn PO, Whittingham LA (2001) Male mask size is correlated with mating success in the common yellowthroat. Anim Behav 62:435–446
Venier LA, Dunn PO, Lifjeld JT, Robertson RJ (1993) Behavioral-patterns of extra-pair copulation in tree swallows. Anim Behav 45:412–415
Webster MS, PruettJones S, Westneat DF, Arnold SJ (1995) Measuring the effects of pairing success, extra-pair copulations and mate quality on the opportunity for sexual selection. Evolution 49:1147–1157
Webster MS, Tarvin KA, Tuttle EM, Pruett-Jones S (2004) Reproductive promiscuity in the splendid fairy-wren: effects of group size and auxiliary reproduction. Behav Ecol 15:907–915
Webster MS, Tarvin KA, Tuttle EM, Pruett-Jones S (2007) Promiscuity drives sexual selection in a socially monogamous bird. Evolution 61:2205–2211
Westneat DF, Stewart IRK (2003) Extra-pair paternity in birds: causes, correlates, and conflict. Annu Rev Ecolog Evol Syst 34:365–396
Whittingham LA, Dunn PO (2005) Effects of extra-pair and within-pair reproductive success on the opportunity for selection in birds. Behav Ecol 16:138–144
Wiley RH, Godard R, Thompson AD (1994) Use of 2 singing modes by hooded warblers as adaptations for signaling. Behaviour 129:243–278
Woolfenden BE, Stutchbury BJM, Morton ES (2005) Male acadian flycatchers, Empidonax virescens, obtain extrapair fertilizations with distant females. Anim Behav 69:921–929
Yezerinac SM, Weatherhead PJ (1997) Extra-pair mating, male plumage coloration and sexual selection in yellow warblers (Dendroica petechia). Proc R Soc Lond, B Biol Sci 264:527–532
Acknowledgment
We would like to thank Stefanie LaZerte and Elizabeth Gow for their enthusiastic help in the field as well as Bonnie Woolfenden for much help with the genetic analysis. This research was supported by a Natural Science and Engineering Research Council of Canada (NSERC) scholarship and York University Faculty of Graduate Studies grants to IC as well as NSERC grants to BJMS.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by J. Podos
Rights and permissions
About this article
Cite this article
Chiver, I., Stutchbury, B.J.M. & Morton, E.S. Do male plumage and song characteristics influence female off-territory forays and paternity in the hooded warbler?. Behav Ecol Sociobiol 62, 1981–1990 (2008). https://doi.org/10.1007/s00265-008-0629-x
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00265-008-0629-x