
Summary
The occurrence of crassulacean acid metabolism (CAM) among epiphytes and related plant species from tropical and subtropical rainforests in Eastern Australia was investigated. As judged from δ13C value and the absence of Kranz anatomy, indications of CAM were found in 66 species belonging to the families, Polypodiaceae (3), Orchidaceae (55), Asclepiadaceae (6) and Rubiaceae (2).
Two thirds of orchidaceous plants examined appeared to use CAM. Those species with thicker leaves generally had less negative δ13C values, as did those species growing on more exposed sites; leaves thicker than about 1 mm in most species yielded δ13C values indicative of pronounced CAM. Two leafless species, Chiloschista phyllorhiza and Taeniophyllum malianum, which depend on chloroplast-containing, stomata-less roots for photosynthesis also showed δ13C values typical of CAM. Pseudobulbs and swollen stems, a characteristic of many orchids, were usually somewhat enriched in 13C compared to corresponding leaves.
In Polypodiaceae CAM was found in the genus Pyrrosia. While δ13C values were generally less negative with increasing frond thickness, the leaf morphology was extremely variable within species. Pyrrosia confluens plants from shaded habitats had long, relatively thin and darkgreen fronds whereas specimens from sun-exposed sites were characterized by short, thick, bleached fronds. Both types showed the capacity for nocturnal accumulation of titratable acidity and exhibited continuous net CO2 fixation during 12 h light/12 h dark cycles under laboratory conditions. Shade-fronds showed this capacity even when irradiance was lower than 2% of full sunlight during the 12 h light period.
In Asclepiadaceae CAM was found in species of two genera which usually have fleshy leaves, Hoya and Dischidia. In Rubiaceae CAM was recorded in two genera of epiphytic ant plants, Hydnophytum and Myrmecodia.
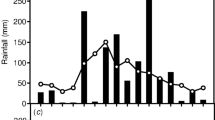
It is concluded that CAM is widespread in Australian epiphytes. It is most prevalent in species found in exposed microhabitats where the growing conditions are characterised by relatively high light intensities and short but frequent periods of water stress.
Similar content being viewed by others

References
Benzing DH (1980) The biology of the Bromeliads. Mad River Press Inc., California
Benzing DH, Ott DW (1981) Vegetative reduction in epiphytic Bromeliaceae and Orchidaceae: Its origin and significance. Biotropica 13:131–140
Coutinho LM (1969) Novas observacoes a ocorrenica do “efeito de de Sassure” e suas relacoes com a suculencia, a temperatura folhear e os movimentos estomaticos. Botanica 24:79–102
Farquhar GD, Ball MC, von Caemmerer S, Roksandic Z (1982) Effect of salinity and humidity on δ13C value of halophytes — evidence for diffusional isotope fractionation determined by the ratio of intercellular/atmospheric partial pressure of CO2 under different environnmental conditions. Oecologia (Berlin) 52:121–124
Gessner F (1956) Der Wasserhaushalt der Epiphyten und Lianen. In Ruhland W (ed) Handbuch der Pflanzenphysiologie Bd III, Pflanze und Wasser. Springer, Berlin Göttingen Heidelberg, 938–950
Goh CJ, Avadhani PN, Loh CS, Hanegraaf C, Arditti J (1977) Diurnal stomatal and acidity rhythms in orchid leaves. New Phytol 78:365–372
Hohorst HJ (1970) L-(-)-Malat, Bestimmung mit Malatdehydrogenase und NAD. In Bergmeyer HU (ed) Methoden der enzymatischen Analyse Bd II. Verlag Chemie, Weinheim, 1544–1548
Kausch W (1965) Beziehungen zwischen Wurzelwachstum. Transpiration und CO2-Gaswechsel bei einigen Kakteen. Planta 66:229–238
Medina E (1975) Dark CO2 fixation, habitat preference and evolution within the Bromeliaceae. Evolution 28:677–686
Medina E, Delgado M (1976) Photosynthesis and night CO2 fixation in Echeveria columbiana v. Poellnitz. Photosynthetica 10:155–163
Medina E, Delgado M, Troughton JH, Medina JD (1977) Physiological ecology of CO2 fixation in Bromeliaceae. Flora 166:137–152
Medina E, Minchin P (1980) Stratification of δ13C values of leaves in Amazonian rain forests. Oecologia (Berlin) 45:377–378
Medina E, Troughton JH (1974) Dark CO2 fixation and carbon isotope ratio in Bromeliaceae. Plant Sci Lett 2:357–362
Neales TF, Hew CS (1975) Two types of carbon fixation in tropical orchids. Planta 123:303–306
O'leary MH (1981) Carbon isotope fractionation in plants. Phytochem 20:553–567
Osmond CB, Nott DL, Firth PM (1979) Carbon assimilation patterns and growth of the introduced CAM plant Opuntia inermis in Eastern Australia. Oecologia (Berlin) 40:331–350
Powles SB, Osmond CB (1978) Inhibition of the capacity and efficiency of photosynthesis in bean leaflets illuminated in a CO2-free atmosphere at Low O2: a possible role for photorespiration. Aust J Plant Physiol 5:619–629
Powles SB, Osmond CB, Thorne SW (1979) Photoinhibition of intact attached leaves of C3 plants illuminated in the absence of both carbon dioxide and photorespiration. Plant Physiol 64:982–988
Richards PW (1952) The tropical rainforest. Camb Univ Press, London-New York-Melbourne
Rowley G (1978) Illustrated encyclopedia of succulents. Salamander, London
Szarek SR, Ting IP (1974) Seasonal patterns of acid metabolism and gas exchange in Opuntia basilaris. Plant Physiol 54:76–81
Teeri JA, Tonsor SJ, Turner M (1981) Leaf thickness and carbon isotope composition in the Crassulaceae. Oecologia (Berlin) 50: 367–369
Wallace BJ (1982) The Australian vascular epiphytes: flora and ecology. PhD thesis, Univ New England, Armidale, Australia
Walter H (1951) Grundlagen der Pflanzenverbreitung 1. Teil: Standortslehre. Ulmer, Stuttgart
Willis JC, Airy Shaw HK (1973) A dictionary of the flowering plants and ferns. Camb Univ Press, London-New York
Winter K, Osmon CB, Pate JS (1981) Coping with salinity. In Pate JS, McComb AJ (eds) The biology of Australian plants. University of Western Australia Press, Nedlands, 88–113
Wong SC, Hew CS (1976) Diffusive resistance, titratable acidity, and CO2 fixation in two tropical epiphytic ferns. Amer Fern J 66:121–124
Author information
Authors and Affiliations
Rights and permissions
About this article
Cite this article
Winter, K., Wallace, B.J., Stocker, G.C. et al. Crassulacean acid metabolism in australian vascular epiphytes and some related species. Oecologia 57, 129–141 (1983). https://doi.org/10.1007/BF00379570
Received:
Issue Date:
DOI: https://doi.org/10.1007/BF00379570