
Abstract
Reproductive traits at the beginning and the end of the annual reproductive season were compared between two populations of the intertidal crab Chasmagnathus granulatus living in ecologically contrasting habitats: (1) Mar Chiquita (MC) (37°45′S, 57°19′W), a highly productive estuarine coastal lagoon with strong salinity fluctuations. (2) San Antonio Bay (SA) (40°46′S, 64°50′), a physically stable but less productive coastal marine environment. Number, size, and elemental composition (CHN) of eggs and larvae differed significantly between populations. Regardless of the season, more but smaller eggs and larvae were produced in MC, while eggs and larvae from SA revealed higher dry mass and C/N ratios indicating higher lipid content. A latitudinal temperature gradient cannot explain these patterns, suggesting that other environmental factors including salinity, quality or quantity of benthic food sources and productivity may be responsible. In both populations, fecundity and biomass per egg were higher at the beginning as compared to the end of the reproductive season. As a consequence, the reproductive effort was consistently maximal at the beginning of the season. At MC, also variability was found between two successive years. Intraspecific (both interpopulational and seasonal) variations in reproductive and developmental traits may be important for the formation of physiologically different metapopulations along the wide geographic range of C. granulatus.
Similar content being viewed by others


Introduction
It is generally assumed that life-history patterns are adaptive, shaped by selection pressures maximizing the survival of offspring. In this context, reproductive strategies have been compared among species of brachyuran crabs (Hines 1986a, b; Hartnoll and Gould 1988; Anger 1995). Additionally, there is also growing evidence of intraspecific variability in nearly all aspects of the life histories of crabs and other marine invertebrates (Hines 1986a, b; Hadfield and Strathmann 1996).
This variability occurs in response to regionally varying environmental conditions and, as a consequence, at least some life-history traits may vary with the prevailing conditions along the distributional range of a species. Latitudinal and/or habitat-related intraspecific differences in the reproductive biology of different populations have been reported for several marine decapod crustaceans (e.g., Jones and Simons 1983; Simons and Jones 1981; Wehrtmann and Kattner 1998; Kyomo 2000; Lardies and Castilla 2001; Brante et al. 2003, 2004; Ouellet and Plante 2004).
Seasonal effects have been identified as another source of intraspecific variation in the reproduction of marine invertebrates (Ghiselin 1987). In a few cases, this phenomenon has been documented also for decapod crustaceans. Seasonal changes were observed, for example, in the number and size of eggs produced by Emerita talpoidea (Díaz 1980), or in the biochemical composition of the embryos of Callinectes sapidus (O’Leary Amsler and George 1984). In a shrimp, Crangon crangon, egg size, larval biomass, duration of larval development, and larval starvation resistance were found to vary between seasons (Paschke et al. 2004). Similarly, there are interannual changes in egg size and larval condition of lobsters (Ouellet and Plante 2004).
The subject of this study, Chasmagnathus granulatus Dana 1853 (Brachyura, Varunidae), is a burrowing, semiterrestrial crab. It is common in warm temperate coastal areas of the southwestern Atlantic, occurring in from Rio de Janeiro, Brazil, to the Gulf of San Matías, Argentina (Boschi 1964). Dense populations of this species are found in estuarine mudflats and salt marshes, as well as in coastal bays and lagoons with highly variable salinities, temperatures, and tidal patterns (Spivak 1997). Studies of different populations of this species have shown differences, particularly between marine and estuarine habitats, in maximum adult body size, egg size, and in the timing and duration of the reproductive season (see Botto and Irigoyen 1979; D’Incao et al. 1992; Anger et al. 1994; Ruffino et al. 1994; Spivak et al. 1994, 1996; Iribarne et al. 1997; Luppi et al. 1997, 2002; López-Greco and Rodríguez 1999; Ituarte et al. 2004; Bas et al. 2005).
The principal aim of this study was to enhance the understanding of spatial and seasonal variations in reproductive traits of C. granulatus. Specifically, this comprises seasonal variability in fecundity, reproductive effort (RE), size and elemental composition of eggs and newly hatched larvae, and biomass losses during embryonic development. These traits were compared between two populations that live in ecologically contrasting habitats on the southwestern coast of Argentina: (1) Mar Chiquita (MC), an estuarine coastal lagoon characterized by strong and unpredictable salinity fluctuations. (2) San Antonio Bay (SA), a physically stable coastal marine environment.
Methods
Study areas
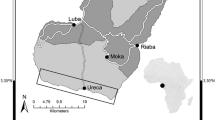
Mar Chiquita is a coastal lagoon located 35 km NE of the city of Mar del Plata, Argentina (37°45′S, 57°19′W; Fig. 1). It has an area of 46 km2, its longest axis running parallel to the coastline. It is divided into a wide northern freshwater part and a narrow southern estuarine part. Water temperature and salinity show great seasonal, daily, and local variations (Anger et al. 1994; Reta et al. 2001). Semidiurnal tides (maximum amplitude: 1.8 m), superimposed by irregular, often strong wind-driven currents, cause an irregular pattern of water exchange through the narrow and shallow mouth. Freshwater enters through creeks and rainfalls (average precipitation: 800 mm/year; Fig. 2), while seawater enters from the adjacent ocean with tidal waves or due to easterly winds. The lower intertidal zones of MC are characterized by mudflats, the higher parts by cordgrass (Spartina densiflora) marshes. Burrows of C. granulatus are found in both habitat types.

Geographical position of the study areas, location of sampling sites (asterisks)

Average monthly rainfall (historical) data and measurements during the study period registered at the meteorological stations of Mar del Plata (MDP) and San Antonio Oeste airports (SAO). Arrows indicate the beginning of the reproductive seasons 2002–2003 and 2003–2004, when Chasmagnathus granulatus females were sampled
San Antonio Bay is located in the northwestern part of the Gulf of San Matías, Río Negro Province, Argentina (40°46′S, 64°50′W; Fig. 1). It belongs to the dry Patagonian region, which is characterized by strong west winds, no freshwater influx from creeks or rivers, and only scarce rainfall (average 240 mm/year; Fig. 2). The tides are semidiurnal, with a maximum amplitude of 9 m (Schnack et al. 1996). The average levels of both temperature and salinity are in the Gulf of San Matías higher than in the open ocean, the latter due to low freshwater influx and excessive evaporation (Piola and Scasso 1988; Scasso and Piola 1988). Most of this bay is fringed by a large intertidal area (ca. 100 km2), covered in part by Spartina alterniflora and Sarcocornia perennis. Burrows of C. granulatus are found throughout this intertidal zone, both in vegetated and unvegetated areas.
Crab collection and maintenance
In the estuarine part of MC and near the town of San Antonio Oeste (SA), ovigerous C. granulatus were collected at the beginning and near the end of the reproductive season (differing between sites; MC: October 7, 2002; March 5, 2003; SA: November 26, 2002; January 27, 2003), respectively (Fig. 1). The variable “reproductive season” is hereafter briefly referred to as “season.” The spring in 2002 was in MC unusually rainy (Fig. 2), so that the females were exposed for extended periods to extremely low salinities (nearly freshwater). This stress situation caused uncommonly high losses of egg masses, mostly occurring within a few days after collection. Therefore, additional samples were taken from both populations at the beginning of the following season (MC: October 17, 2003; SA: November 17, 2003).
Females with eggs in an early stage of embryonic development were transported to the laboratory and placed into plastic aquaria with filtered seawater (32 salinity) and kept at 20°C and natural light conditions. Food (pellets for bottom fishes) was added every 3 days. Egg masses were inspected and staged following Bas and Spivak (2000). Only females whose eggs contained ≥90% of yolk (stages 1–3) were used in our study of fecundity and egg characteristics (egg size, biomass). When the embryos reached stage 9 (ready to hatch), the females were transferred to individual aquaria until hatching occurred (see below, study of larval characteristics).
Fecundity
For estimates of fecundity, we used 30 females from each site and time of sampling. Samples of 400 freshly laid eggs in stage 3 were removed from each of ten females, counted under a stereomicroscope, rinsed in distilled water, dried for 24 h at 60°C in pre-weighed tin cartridges, and re-weighed to the nearest 0.01 mg. All females were frozen to death, sized [maximum carapace width(CW)], and their egg masses completely removed. The females and their egg clutches were dried and weighed separately, and average values of dry weight (DW, or dry mass) of the ten egg samples were used to estimate the total number of eggs in each female.
Size and biomass of eggs in an early developmental stage
At least ten females with eggs in stage 1 or 2 of embryonic development (95–100% of yolk) were obtained from each sampling site and time and used to estimate the volume (V), DW, and elemental composition (carbon, hydrogen, nitrogen contents; collectively CHN) of eggs and larvae. Only C and N data are considered in this paper, because those of H are little used in the literature, rendering comparisons difficult.
Egg volume was calculated from size measurements in ten eggs per brood assuming an ellipsoidal form (V=d 2×D×π/6; where d is the minor, D the major diameter). The CHN content of the eggs was measured in 200 eggs per female (counted under a stereomicroscope; n = 66 females), with five replicate samples of 40 eggs each. The samples were gently rinsed with distilled water, dried on fluff-free filter paper, transferred to pre-weighed tin cartridges, and stored frozen at −20°C until elemental analyses were carried out (see below, CHN analyses).
Size and biomass of newly hatched larvae
From each female (n = 39), a sample of 20–30 recently hatched larvae was fixed in 4% formaldehyde for later measurements of larval carapace volume (CV). This was estimated using the same equation for an ellipsoidal form as in egg measurements (see above), with d = CW, measured between the bases of the lateral spines, and D = carapace length, taken from the base of the rostral spine to the posterior edge of the carapace. Initial larval biomass was measured in 200 zoeae per female, with five replicate samples of 40 larvae each, as DW and CHN content, following the same procedures as for egg samples.
Elemental analyses (CHN)
Eggs and larvae were dried for 48 h in a Finn-Aqua Lyovac GT2E vacuum dryer, weighed to the nearest 0.1 μg on a Mettler UMT2 microbalance, and analyzed in a Carlo Erba Elemental Analyser (EA 1108).
Statistical analyses
Standard statistical analyses were based on Zar (1996) and Underwood (1997). Normality was checked with normal plots and homoscedasticity with Cochran’s test. Fecundity was compared between sites (MC, SA), times (beginning, end of the reproductive season), and years (2002 vs. 2003), computing linear regression lines of egg number (log-transformed) plotted on CW.
Since fecundity observed at the beginning of 2002 was not significantly correlated with CW (neither at MC nor SA), scaled fecundity values (size-specific fecundity, FecR = number of eggs/mm CW) were compared with two-way ANOVA as variable, and site (MC, SA) and time (beginning 2002, beginning 2003, and end 2002) as factors. RE was estimated as DW ratio (total egg dry mass per female dry mass) expressed as percentage and compared in the same way.
The fecundity and the RE of females measured at the beginning of the reproductive seasons 2002–2003 and 2003–2004 differed significantly between years and were therefore analyzed separately. All other data (i.e., biomass and volume of eggs and larvae) obtained at the beginning of the two successive seasons did not differ and, consequently, were pooled.
DW, C, and N content (μg/individual and % of DW), energy content (estimated from C content; Salonen et al. 1976) and the C/N ratio were compared between eggs or larvae from the beginning of the two successive seasons (2002 and 2003), with one-way ANOVA for each variable, with year as factor, for MC and SA separately. Since no differences between years were found in the variables analyzed, data from the beginning of 2002 and 2003 were pooled.
Differences in DW, V, C, N, energy content and the C/N ratio of eggs and larvae were compared with two-way ANOVA, with site (MC, SA) and time (beginning, end of season) as factors. When variances were heterogeneous and no transformations were possible, an approximate test of equality of means with heterogeneous variances was performed in pairs of means using the Games and Howell method (Sokal and Rohlf 1981). Significant differences among treatments after ANOVA were tested with a Student–Newman–Keuls (SNK) test. Linear regressions were calculated to evaluate the relationships between DW, CHN, and energy contents of eggs or larvae from the same female, from both sites, pooling data of beginning and end of season for each one. The homogeneity of slopes and Y-intercepts between populations were analyzed with a test of parallelism and ANCOVA, respectively.
Results
Fecundity and reproductive effort (RE)
At both sites (MC, SA), the number of eggs carried per female correlated at the end of the reproductive season 2002–2003 and at the beginning of the season 2003–2004 significantly with female body size (CW, Fig. 3). Conversely, no significant relationship between these variables was found at the beginning of the season 2002–2003 (linear regressions for materials from MC: R 2 = 0.109, P = 0.08; SA: R 2 = 0.08, and P = 0.126). The differences in fecundity between sites, among the three sampling times (beginning and end of one season and beginning of the next year), and the interaction between these factors, were statistically significant (P < 0.0001, in all cases). Size-specific fecundity (FecR) was in both populations highest at the beginning of the season 2003–2004 (MC: 1,560 eggs/mm CW; SA: 770 eggs/mm CW). The lowest FecR-values were observed at the beginning (MC: 981 eggs/mm) and at the end (SA: 616 eggs/mm) of the season 2002–2003. Intermediate fecundity values were registered at the end (MC: 1,147 eggs/mm) and the beginning (SA: 749 eggs/mm) of the season 2002–2003. On average, fecundity was consistently lower in SA than in MC (Fig. 3).

Chasmagnathus granulatus. Relationship between carapace width (CW) and number of eggs (log-transformed) carried at the beginning of the reproductive seasons 2002–2003 (beginning 2002) and 2003–2004 (beginning 2003), and at the end of the reproductive season 2002–2003 (end 2002). Linear regression for beginning 2002 not shown (statistically not significant)
The RE of females from MC varied significantly among the three sampling times, showing the same pattern as in fecundity. The RE of females from SA, by contrast, was significantly lower at the end of the season 2002–2003 compared to the beginning of the seasons 2002–2003 and 2003–2004 (Fig. 4). When RE was compared between populations, it was at the beginning of the season 2002–2003 significantly lower in MC (P < 0.0001), but higher at the end of the same season (P = 0.022), while it did not differ at the beginning of the season 2003–2004 (P = 0.86).

Chasmagnathus granulatus. Reproductive effort (RE; mean ± SD) of females from Mar Chiquita (MC) and San Antonio (SA) collected at the beginning and end of the reproductive season 2002–2003 (beginning 2002 and end 2002) and at the beginning of the reproductive season 2003–2004 (beginning 2003). Different letters on bars: statistically significant differences
Egg biomass and volume
The individual dry mass (or dry weight, DW) of newly laid eggs varied greatly among conspecific females, ranging between 5.5 and 12.0 μg, while egg volume (V) ranged between 0.0102 and 0.0215 mm3 (Table 1). Both variables differed significantly between sampling times (beginning versus end of season) and between populations. The interaction of these two factors was also significant (Table 2). The same patterns were found in the CHN and energy data (Table 3). In general, the largest eggs had also the highest DW (Table 1). Eggs from the beginning of the season had in both populations, a larger size and higher biomass values compared to eggs produced at the end of the season, with consistently higher values in SA than in MC (Fig. 5).

Chasmagnathus granulatus. Dry mass (DW), carbon (C), nitrogen (N), and energy content of eggs and larvae (mean ± SD), at the beginning (2002 and 2003, data pooled) and end of the reproductive season. Numbers above columns: percentage of gain (+) or loss (−) of each variable between eggs and larvae (see Table 6 for ANOVA)
When CHN and energy values were expressed as fractions (%) of DW, as elemental mass ratio (C/N), or as DW-specific energy content (in J/mg DW), these data (except for the percentage N-values) differed significantly between the beginning and end of the season, being consistently higher at the beginning (Tables 4, 5). No significant differences, however, were found between populations.
Larval biomass and volume
The dry mass of freshly hatched zoea-I larvae did not differ significantly from that of newly laid eggs (on average by only 1%; see Fig. 5, Table 6), although larval CV was almost twice as large as the initial egg volume, V (Table 1). As in the eggs, larval DW and CV varied also significantly between the beginning and the end of the season and between populations, but the interaction between these factors was insignificant (Table 2). Larvae hatching at the beginning of the season were consistently heavier and larger than those produced at the end, and larvae from SA had always-higher values of DW and V than those from MC (Fig. 5).
When larval biomass was expressed as CHN or energy content per individual, these values were always lower than in the eggs from which they hatched (i.e., from the same brood; Fig. 5). The proportion of C lost during the embryonic development and at hatching differed significantly between populations and times within season (beginning versus end; Table 6). These embryonic C losses were higher in larvae from MC than in those from SA, and they were in both populations higher at the end of the reproductive season (Fig. 5). The losses of energy, however, differed significantly only between populations, but not between the beginning and end of the season, and those in N were generally insignificant (Table 6, Fig. 5).
The values of larval DW, CHN, and energy content per individual measured at hatching showed significant positive correlations with the initial values found in the eggs at an early stage of embryonic development (Fig. 6). The slopes of these regressions did not differ between the two populations, while the Y-intercept differed significantly in DW, but not in other biomass parameters (Fig. 6, Table 7). Also, larval CV was in materials from MC, but not from SA, significantly correlated with egg V (R 2 = 0.33, P = 0.021 vs. R 2 = 0.22, P = 0.073).

Chasmagnathus granulatus. Relationship between dry mass (DW), carbon (C), nitrogen (N), and energy content of newly laid eggs and of newly hatched larvae (Z I) from the same brood. SA San Antonio, MC Mar Chiquita Tables
In the proportions of CHN within larval DW (%) and the DW-specific energy values of the larvae, significant seasonal variation was detected, with higher values at the beginning of the season. Also, the C/N mass ratio was significantly higher at the beginning of the season, and it was higher in larvae from SA compared to those from MC. The percentage N-values, by contrast, did not significantly vary between populations or times within season. The interaction between these factors was significant only in the C/N ratio (Tables 4, 8).
Discussion
Intraspecific variability in the number and size of eggs produced by invertebrate species under varying environmental conditions may aid to optimizing the relative allocation of female energy resources to growth and reproduction, as well as survival, dispersal, and recruitment of the offspring (Hadfield and Strathmann 1996). In decapod crustaceans, fecundity (defined as number of eggs per batch produced by a female) and the average size of the eggs may also vary among conspecific populations, tending either to many small or to less but larger eggs. Such differences have been related to latitudinal temperature gradients (e.g., Jones and Simons 1983; Clarke et al. 1991; Lardies and Castilla 2001; Lardies and Wehrtmann 2001; Wehrtmann and López 2003), interannual temperature variation (Simons and Jones 1981; Díaz 1980; Boddeke 1982; Ouellet and Allard 2002; Ouellet and Plante 2004; Brillon et al. 2005), regional variation in salinity, or other environmental factors (Mashiko 1982, 1990; Hancock et al. 1998). Besides the number and size, also the chemical composition of the eggs may vary among populations (O’Leary Amsler and George 1984; Wehrtmann and Kattner 1998). Generalizations of causes and consequences of intraspecific variation in reproductive traits, however, remain difficult, mostly due to scarce knowledge of carry-over effects, which occur between successive ontogenetic phases such as embryonic and larval development (Hadfield and Strathmann 1996; Marshall et al. 2003).
In our study, we observed significant differences in the number, size, dry mass, and elemental composition (CHN) of eggs produced by two spatially separated populations of an intertidal crab, C. granulatus. Moreover, we found in both populations significant seasonal variation. Regardless of the season, on average more but smaller eggs were produced in MC, or fewer but larger eggs in SA. Due to this trade-off, the RE did not vary between these populations at the beginning of the reproductive season, except when unusual environmental conditions occurred (extremely low salinities in MC in 2002). However, considering that the RE was lower in SA at the end of the season, and that the season is 50% longer in MC (6 months vs. 4 months in SA), the RE in a whole reproductive season will be lower in SA.
The pattern of variation in reproductive traits (trade-off between number and size of eggs) observed between a northern (MC, 37°45′S) and a southern population (SA, 40°46′S) resembles latitudinal differences found in other decapod species, where larger egg size and lower fecundity occurred in colder regions within the distributional range (see Clarke et al. 1991; Lardies and Castilla 2001; Lardies and Wehrtmann 2001; Wehrtmann and López 2003; Brante et al. 2003, 2004). In this particular case, however, a latitudinal temperature gradient cannot explain the observed patterns, because peculiar regional oceanographic processes cause a reversal of the commonly known pattern of temperature distribution, with higher water temperatures at SA compared to MC (Piola and Scasso 1988; Scasso and Piola 1988).
As another environmental factor, different average quality and/or quantity of available benthic food sources may cause differences in the reproductive traits of separate populations (Harrison 1990). This can affect the condition of reproducing females and, through carry-over effects, that of eggs and larvae (see Cahu et al. 1995; Cavalli et al. 1999; Palacios et al. 1998, 1999; Lardies et al. 2004). C. granulatus feeds on both Spartina plants and soft sediments (Iribarne et al. 1997), which may vary qualitatively among habitats. However, there is no comparative information available on possibly differential quality or quantity of plants at MC and SA, and differences in the food quality of sediments cannot account for the patterns observed in the size and biomass of eggs and early larvae (larger in SA compared to MC), as the sediments in MC tend to be richer in organic matter than in SA (Bas et al. 2005). Future comparative ecological field studies are thus necessary to support or exclude variation in nutritional conditions in the benthos as a possible explanation for interpopulational differences in the reproduction of C. granulatus.
Exceptionally low fecundity observed only at MC at the beginning of the season 2002–2003 was most probably a consequence of unusual environmental conditions with extremely low salinities after heavy rainfalls, thus representing a case of interannual rather than seasonal or regional variability. In spite of dramatic variation in fecundity, the average size and biomass (V, DW, and CHN) per egg was not affected by these temporarily and locally occurring adverse conditions. This indicates variation in the overall energy investment into female reproduction (i.e., per egg batch), but constant investments per offspring (or per egg).
In both populations fecundity varied seasonally, with consistently higher egg numbers produced in early versus late season. Since the average biomass per egg followed the same seasonal pattern as the number of eggs per female, the RE was consistently maximal at the beginning of the season. A similar pattern was observed in a population of the mole crab, Emerita talpoida (Díaz 1980). As in C. granulatus, the seasonal decrease in egg mass coincided with rising water temperature. In our study, water temperature increased in both populations by ca. 4°C from November to January (ca. 16–20°C in MC, 18–22°C in SA). Among the principal mortality factors affecting decapod larvae in the field, predation and starvation may be most important (Morgan 1995). Large eggs and larvae with particularly high C/N ratios (indicating high lipid contents; Anger 2001) observed at the beginning of the reproductive season suggest that larvae produced early in the season should be especially well adapted to food limitation, which may occur during a long period of larval development at relatively low water temperatures (Boddeke 1982; Anger 2001; Paschke et al. 2004).
Since C. granulatus produces several successive clutches during a season, smaller clutch size, and egg biomass near the end of the reproductive season could also be a consequence of reproductive exhaustion of the females (see Palacios et al. 1999).
Giménez and Anger (2003) found a significant positive correlation between the initial biomass (measured as dry mass or μg C/larva) of recently hatched zoea-I larvae of C. granulatus and subsequent larval survival through metamorphosis to the first juvenile stage. Variation in egg biomass occurred between broods from different females that had been kept at the same condition, but also as a response to reduced salinity (15) during egg development. Such differences in egg size persisted at least to the zoea IV stage, with a tendency of increased growth rate in larvae hatched from larger eggs (Giménez and Torres 2002). Higher egg biomass observed at the beginning of the reproductive season in MC could also be caused by low salinities, which generally occur during this period due to strong rainfalls and low evaporation during the preceding autumn and winter. On the other hand, the same pattern (larger eggs at the beginning of the season) was also observed in SA, where salinity is never lower than seawater (see below).
Regardless of the season, eggs and zoea-I larvae of C. granulatus from SA were consistently larger, contained more biomass (DW, CHN), and showed higher C/N ratios than those from MC. This may indicate different patterns of yolk provisioning during oogenesis or differential utilization of yolk during embryogenesis in the two populations compared here. In estuaries, fronts and currents may produce local patches with high productivity, aiding also in the avoidance of an excessive larval dispersal toward the open sea, and providing physical and chemical cues for the reinvasion of megalopae (Young 1995; Forward et al. 2001). In MC, advanced developmental stages (zoeal stages II–IV) of C. granulatus are found only nearshore, north of the mouth of the lagoon (Bas, personal observation). Productivity in the area of MC is high due to nutrients supplied by agricultural activities in the surrounding region, which are leached from soils by freshwater runoff (Marcovecchio et al. 2005) and then transported northward by the coastal drift (Bas, personal observation).
In SA Bay, by contrast, productivity is low. The waters of the inner parts of the San Matías Gulf enter and leave this bay twice a day. There is no freshwater input except for occasional rainfalls in San Matías Gulf, and there is no agricultural use of the land in this region. As a consequence, the productivity of the waters of the Gulf of San Matías is generally low with 0.03–0.18 mg/m3 chlorophyll a in the inner parts, and 0.16–1.28 mg/m3 in the more productive outer parts, which receive water coming from the south (Carreto et al. 1974). By comparison, Marcovecchio et al. (2005) measured chlorophyll a concentrations of 40–60 mg/m3 in MC. The pattern of larval dispersal of C. granulatus in SA is not known, but it is likely that larval development takes place inside the San Matías Gulf. This is due to a particular pattern of circulation, which largely retains the water masses and plankton, especially during the reproductive season of C. granulatus in spring and summer (Lucas et al. 2005). Compared to larvae from MC, the relatively large zoeae from SA may be more efficient in obtaining food (Morgan 1995). Alternatively or in addition, they may tolerate starvation periods in waters with low productivity better, as they reveal higher average C and C/N-values, indicating larger lipid reserves (Anger 2001). Moreover, the larvae of C. granulatus are generally well adapted to survive in patchy environments with temporally or spatially limited availability of planktonic food (Giménez and Anger 2005).
Taking into account the phenotypic plasticity of some reproductive traits of C. granulatus, our study suggests that intraspecific (both interpopulational and seasonal) variation in these traits may be important for the formation of physiologically different metapopulations across the wide geographic range of this species, ranging from Patagonia, Argentina, to the tropical region of Rio de Janeiro, Brazil (Giménez 2003). More studies will be necessary to separate plastic (i.e., reversible) from non-plastic (irreversible) differences between populations. The maximal degree of differentiation should be defined by the actual connectivity between populations, which has hardly been studied in the field.
References
Anger K (1995) The conquest of freshwater and land by marine crabs: adaptations in life-history patterns and larval bioenergetics. J Exp Mar Biol Ecol 193:119–145
Anger K (2001) The biology of decapod crustacean larvae. In: Vonk R (ed) Crustacean issues 14. A A Balkema Publishers, Lisse, The Netherlands
Anger K, Bas C, Spivak E, Ismael D, Luppi T (1994) Hatching rhythms and dispersion of decapod crustacean larvae in a brackish coastal lagoon in Argentina. Helgoländ Meeresun 48:445–466
Bas C, Spivak E (2000) Effect of salinity on embryos of two Southwestern Atlantic estuarine grapsid crab species cultured in vitro. J Crustacean Biol 20:647–656
Bas C, Luppi T, Spivak E (2005) Population structure of the South American estuarine crab, Chasmagnathus granulatus (Brachyura: Varunidae) near the southern limit of its geographical distribution: comparison with northern populations. Hydrobiologia 537:217–228
Boddeke R (1982) The occurrence of winter and summer eggs in the brown shrimp (Crangon crangon) and the pattern of recruitment. Neth J Sea Res 16:151–162
Boschi E (1964) Los crustáceos decápodos Brachyura del litoral bonaerense (R Argentina). Bol Inst Biol Mar, Mar del Plata 6:1–99
Botto J, Irigoyen H (1979) Bioecología de la comunidad del cangrejal. 1: contribución al conocimiento biológico del cangrejo de estuario, Chasmagnathus granulata Dana (Crustacea, Decapoda, Grapsidae) en la desembocadura del Río Salado, provincia de Buenos Aires. Sem Lat Ecol Bent Sed Plat Cont Atl Sur. UNESCO, Montevideo, Uruguay, pp 161–169
Brante A, Fernández M, Eckerle L, Mark F, Pörtner H, Arntz W (2003) Reproductive investment in the crab Cancer setosus along a latitudinal cline: egg production, embryo losses and embryo ventilation. Mar Ecol Prog Ser 251:221–232
Brante A, Cifuentes S, Pörtner H, Arntz W, Fernández M (2004) Latitudinal comparison of reproductive traits in five Brachyuran species along the Chilean coast. Rev Chil Hist Nat 77:15–27
Brillon S, Lambert Y, Dodson J (2005) Egg survival, embryonic development, and larval characteristics of northern shrimp (Pandalus borealis) females subject to different temperature and feeding conditions. Mar Biol 147:895–911
Cahu C, Cuzon G, Quazuguel P (1995) Effect of highly unsaturated fatty acids, a-tocopherol and ascorbic acid in broodstock diet on egg composition and development of Penaeus indicus. Comp Biochem Physiol 112A:417–424
Carreto J, Verona C, Casal A, Laborde M (1974) Fitoplancton, pigmentos y condiciones ecológicas del golfo San Matías. III. An Com Invest Cient Prov Bs As, Informe 10:49–76
Cavalli R, Lavens P, Sorgeloos P (1999) Performance of Macrobrachium rosenbergii broodstock fed diets with different fatty acid composition. Aquaculture 179:387–402
Clarke A, Hopkins C, Nilssen E (1991) Egg size and reproductive output in the deep water prawn Pandalus borealis Krøyer, 1838. Funct Ecol 5:724–730
D’Incao F, Ruffino M, da Silva K, da Costa Braga A (1992) Responses of Chasmagnathus granulata Dana (Decapoda: Grapsidae) to salt-marsh environmental variations. J Exp Mar Biol Ecol 161:179–188
Díaz H (1980) The mole crab Emerita talpoida (Say): a case of changing life history pattern. Ecol Monogr 50:437–456
Forward R, Tankersley R, Rittschof D (2001) Cues for metamorphosis of brachyuran crabs: an Overview. Am Zool 41:1108–1122
Ghiselin M (1987) Evolutionary aspects of marine invertebrate reproduction. In: Giese A, Pearse J, Pearse V (eds) Reproduction of marine invertebrates, vol IX. Blackwell, CA, pp 609–665
Giménez L (2003) Potential effects of physiological plastic responses to salinity on population networks of the estuarine crab Chasmagnathus granulata. Helgoland Mar Res 56:265–273
Giménez L, Anger K (2003) Larval performance in an estuarine crab, Chasmagnathus granulata, is a consequence of both larval and embryonic experience. Mar Ecol Prog Ser 249:251–264
Gimenez L, Anger K (2005) Effects of temporary food limitation on survival and development of brachyuran crab larvae. J Plankton Res 27:485–494
Giménez L, Torres G (2002) Larval growth in the estuarine crab Chasmagnathus granulata: the importance of salinity experienced during embryonic development, and the initial larval biomass. Mar Biol 141:877–885
Hadfield M, Strathmann M (1996) Variability, flexibility and plasticity in life histories of marine invertebrates. Oceanol Acta 19:323–334
Hancock M, Hughes J, Bunn S (1998) Influence of genetic and environmental factors on egg and clutch sizes among populations of Paratya australiensis Kemp (Decapoda: Atyidae) in upland rainforest streams, south-east Queensland. Oecologia 115:483–491
Harrison K (1990) The role of nutrition in maturation, reproduction and embryonic development of decapod crustaceans: a review. J Shelf Res 9:1–28
Hartnoll R, Gould P (1988) Brachyuran life history strategies and the optimization on egg production. Symp Zool Soc Lond 59:1–9
Hines A (1986a) Larval patterns in the life histories of brachyuran crabs (Crustacea, Decapoda, Brachyura). Bull Mar Sci 39:444–466
Hines A (1986b) Larval problems and perspectives in life histories of marine invertebrates. Bull Mar Sci 39:506–525
Iribarne O, Bortolus A, Botto F (1997) Between-habitat differences in burrow charcteristics and trophic modes in the southwestern Atlantic burrowing crab Chasmagnathus granulata. Mar Ecol Prog Ser 155:137–145
Ituarte R, Spivak E, Luppi T (2004) Female reproductive cycle of the Southwestern Atlantic estuarine crab Chasmagnatus granulatus (Brachyura: Grapsoidea: Varunidae). Sci Mar 68:127–137
Jones M, Simons M (1983) Latitudinal variation in reproductive characteristics of a mud crab, Helice crassa (Grapsidae). Bull Mar Sci 33:656–670
Kyomo J (2000) Intraspecific variation of reproductive strategies of the crab Sesarma intermedia: a consequence of habitat variations. Bull Mar Sci 66:157–171
Lardies M, Castilla J (2001) Latitudinal variation in the reproductive biology of the commensal crab Pinnaxodes chilensis (Decapoda: Pinnotheridae) along the Chilean coast. Mar Biol 139:1125–1133
Lardies M, Wehrtmann I (2001) Latitudinal variation in the reproductive biology of Betaeus truncatus (Decapoda: Alpheidae) along the Chilean coast. Ophelia 55:55–67
Lardies M, Carter M, Bozinovic F (2004) Dietary effects on life history traits in a terrestrial isopod: the importance of evaluating maternal effects and trade-offs. Oecologia 138:387–395
López-Greco L, Rodríguez E (1999) Annual reproduction and growth of adult crabs Chasmagnathus granulata (Crustacea, Brachyura, Grapsidae). Cah Biol Mar 40:155–164
Lucas A, Guerrero R, Mianzán H, Acha M, Lasta C (2005) Coastal oceanographic regimes of the Northern Argentine Continental Shelf (34–43 °S). Estuar Coast Shelf Sci 65:405–420
Luppi T, Bas C, Spivak E, Anger K (1997) Fecundity of two grapsid crab species in the Laguna Mar Chiquita, Argentina. Arch Fish Mar Res 45:149–166
Luppi T, Spivak E, Anger K (2002) Postsettlement growth of two estuarine crab species, Chasmagnathus granulata and Cyrtograpsus angulatus (Crustacea, Decapoda, Grapsidae): laboratory and field investigations. Helgoland Mar Res 55:293–305
Marcovecchio J, Freije H, De Marco S, Ferrer L, Andrade S, Beltrame O, Asteasuain R (2005) Seasonality of hydrographic variables in a coastal lagoon: Mar Chiquita, Argentina. Aquatic Conserv Mar Freshw Ecosyst 15:180–191
Marshall D, Pechenik J, Keough M (2003) Larval activity levels and delayed metamorphosis affect post-larval performance in the colonial ascidian Diplosoma listerianum. Mar Ecol Prog Ser 246:153–162
Mashiko K (1982) Differences in both the egg size and the clutch size of the fershwater prawn Palaemon paucidens De Haan in the Sagami river. Jpn J Ecol 32:445–451
Mashiko K (1990) Diversified egg and clutch sizes among local populations of the freshwater prawn Macrobrachium nipponense (De Haan). J Crustacean Biol 10:306–314
Morgan S (1995) Life and death in the plankton: larval mortality and adaptation. In: McEdwards L (ed) Larval ecology of marine invertebrates. CRC, Boca Ratón, FL, pp 279–321
O’Leary Amsler M, George R (1984) Seasonal variation in the biochemical composition of the embryos of Callinectes sapidus Rathbun. J Crustacean Biol 4:546–553
Ouellet P, Allard J (2002) Seasonal and interannual variability in larval lobster Homarus americanus size, growth and condition in the Magdalen Islands, southern Gulf of St. Lawrence. Mar Ecol Prog Ser 230:241–251
Ouellet P, Plante F (2004) An investigation of the sources of variability in American lobster (Homarus americanus) eggs and larvae: female size and reproductive status, and interannual and interpopulation comparisons. J Crustacean Biol 24:481–495
Palacios E, Ibarra A, Ramırez J, Portillo G, Racotta I (1998) Biochemical composition of egg and nauplii in White Pacific shrimp Penaeus Íannamei Boone, in relation to the physiological condition of spawners in a commercial hatchery. Aquacult Res 29:183–189
Palacios E, Perez-Rostro C, Ramirez J, Ibarra A, Racotta I (1999) Reproductive exhaustion in shrimp Penaeus vannamei reflected in larval biochemical composition, survival and growth. Aquaculture 171:309–321
Paschke KA, Gebauer P, Buchholz F, Anger K (2004) Seasonal variation in starvation resistance of early larval North Sea shrimp Crangon crangon (Decapoda: Crangonidae). Mar Ecol Prog Ser 279:183–191
Piola A, Scasso L (1988) Circulación en el Golfo San Matias. Geoacta 15:33–51
Reta R, Martos P, Perillo G, Piccolo C, Ferrante A (2001) Características hidrográficas del estuario de la laguna Mar Chiquita. In: Iribarne O (ed) Reserva de biósfera mar chiquita: características físicas, biológicas y ecológicas. Ed. Martín, Mar del Plata, pp 31–41
Ruffino M, Telles M, D’Incao F (1994) Reproductive aspects of Chasmagnathus granulata Dana, 1851 (Decapoda, Grapsidae) in the Patos lagoon estuary, Brazil. Nauplius 2:43–52
Salonen K, Sarvala J, Hakala I, Vilijanen M (1976) The relation of energy and organic carbon in aquatic invertebrates. Limnol Oceanogr 21:724–730
Scasso L, Piola A (1988) Intercambio neto de agua entre el mar y la atmósfera en el Golfo San Matias. Geoacta 15:13–31
Schnack E, Aliotta S, Isla F, Lizasoain G (1996) Formas de fondo asociadas a un delta de reflujo macromareal, San Antonio Oeste, Provincia de Rio Negro, Argentina. Thalassas 12:121–135
Simons M, Jones M (1981) Population and reproductive biology of the mud crab, Macrophtalmus hirtipes (Jaquinot, 1853) (Ocypodidae), from marine and estuarine habitats. J Nat Hist 15:981–994
Sokal R, Rohlf J (1981) Biometry. The principles and practice of statistics in biological research, 2nd edn. W. H. Freeman, NY, USA
Spivak E (1997) Cangrejos estuariales del Atlántico sudoccidental (25°–41° S) (Crustacea: Decapoda: Brachyura). Invest Mar, Valparaíso 25:105–120
Spivak E, Anger K, Bas C, Luppi T, Ismael D (1996) Size structure, sex ratio, and breeding season in two intertidal grapsid crab species from Mar Chiquita Lagoon, Argentina. Nerítica 10:7–26
Spivak E, Anger K, Luppi T, Bas C, Ismael D (1994) Distribution and habitat preferences of two grapsid crab species in Mar Chiquita Lagoon (Province of Buenos Aires, Argentina). Helgolän Meeresun 48:59–78
Underwood A (1997) Experiments in ecology. Cambridge University Press, Cambridge, UK
Wehrtmann I, Kattner G (1998) Changes in volume, biomass, and fatty acids of developing eggs in Nauticaris magellanica (Decapoda: Caridea): a latitudinal comparison. J Crustacean Biol 18:413–422
Wehrtmann I, López G (2003) Effects of temperature on the embryonic development and hatchling size of Betaeus emarginatus (Decapoda: Caridea: Alpheidae). J Nat Hist 37:2165–2178
Young C (1995) Behavior and locomotion during the dispersal phase of larval life. In: McEdward L (ed) Ecology of marine invertebrate larvae. CRC, Boca Raton, FL, pp 279–321
Zar J (1996) Biostatistical analysis, 3rd edn. Prentice Hall, Upper Saddle River, NJ, USA
Acknowledgments
This study was funded by grants from the Universidad de Mar del Plata (15/E149) and the Consejo Nacional de Investigaciones Científicas y Técnicas de la República Argentina (CONICET; PIP 0838/98) conferred to EDS. CCB acknowledges a postdoctoral fellowship from CONICET. Our sincere thanks are also due to Mrs. Karin Bickmeyer (Helgoland) for carrying on innumerable CHN analyses.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by H.-D. Franke.
Rights and permissions
About this article
Cite this article
Bas, C.C., Spivak, E.D. & Anger, K. Seasonal and interpopulational variability in fecundity, egg size, and elemental composition (CHN) of eggs and larvae in a grapsoid crab, Chasmagnathus granulatus . Helgol Mar Res 61, 225–237 (2007). https://doi.org/10.1007/s10152-007-0070-y
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10152-007-0070-y