
Abstract
The purpose of this paper was to investigate patterns of demosponge distribution along gradients of environmental conditions in the biogeographical subzones of the eastern Mediterranean (Aegean and Levantine Sea). The Aegean Sea was divided into six major areas on the basis of its geomorphology and bathymetry. Two areas of the Levantine Sea were additionally considered. All available data on demosponge species numbers and abundance in each area, as well as their vertical and general geographical distribution were taken from the literature. Multivariate analysis revealed a NW–SE faunal gradient, showing an apparent dissimilarity among the North Aegean, the South Aegean and the Levantine Sea, which agrees with the differences in the geographical, physicochemical and biological characteristics of the three areas. The majority of demosponge species has been recorded in the North Aegean, while the South Aegean is closer, in terms of demosponge diversity, to the oligotrophic Levantine Sea. The number of studies in the Aegean and Levantine subareas was positively correlated with the number of species recorded within each Aegean subarea. Demosponge species with an Altanto-Mediterranean distribution prevailed in the Aegean and the Levantine. The reduced contribution of the endemic component, as compared to the western Mediterranean, is consistent with the general NW–SE decrease in the number of endemic species in the Mediterranean. Demosponge distribution at the order level showed also a NW–SE gradient, similar to that observed in the entire Atlantic–Mediterranean region, suggesting a warm water affinity. Sublittoral, circalittoral and bathyal zones were clearly distinguishable in the Aegean Sea on the basis of their sponge fauna. The total number of species was an exponential negative function of depth.
Similar content being viewed by others

Introduction
It is generally accepted that the Mediterranean Sea is one of the world’s most oligotrophic seas. Conspicuously, it harbors somewhat between 4% and 18% of the known world marine species, while representing only 0.82% in surface area and 0.32% in volume of the world ocean (Bianchi and Morri 2000). The eastern Mediterranean, and especially the Levantine basin, is considered as the most oligotrophic Mediterranean region, having a much lower trophic potential than that of the western Mediterranean (Azov 1991). The diversity of the Mediterranean fauna exhibits a west-to-east decline (e.g., Fredj et al. 1992) with the western Mediterranean supporting about 87.1%, the Adriatic Sea 48.9% and the eastern Mediterranean 43.1% of the total Mediterranean species. This gradient has been more or less confirmed by several studies on certain benthic groups such as decapods (Koukouras et al. 1992), prosobranch gastropods (Koutsoubas et al. 1997), polychaetes (Arvanitidis et al. 2002) and anthozoans (Vafidis 2002). The Aegean Sea, however, being the second major sea of the eastern Mediterranean after the Levantine, does not seem to follow this general trend since it supports higher species numbers than the latter and the Adriatic Sea. This strengthens the necessity of a clear subdivision of the eastern Mediterranean, separating the Aegean from the Levantine Sea as pointed out by Por (1978).
Until recently, our knowledge about species composition, abundance and diversity of the sponge fauna of the Aegean Sea was scattered in a number of publications. Voultsiadou (2005) reviewed the existing information on the Porifera of this area and provided a checklist of the Aegean sponge fauna including 200 demosponge species. Levantine demosponges are known from a restricted number of studies carried out on the coasts of Egypt, Israel and Lebanon. About 43% of the total demosponge species known from the Mediterranean are represented in the eastern basin (the Aegean and Levantine Sea) (Voultsiadou 2005).
Sponge diversity in the Aegean Sea seems to exhibit spatial differences. Kefalas et al. (2003) found a relatively higher demosponge species number and abundance in the circalittoral of some islands of the North Aegean in comparison with southern Aegean areas. Voultsiadou (2005) indicated that the different degrees of exploration in the south and the north part of the Aegean might play a role in the difference in species numbers between the two areas, thus suggesting a thorough analysis of sponge distribution in the various subareas of the Aegean.
Based on the above-mentioned information, the main goal of the present paper is to investigate the pattern of demosponge species distribution in the various subareas of the Aegean and the Levantine Sea in order to test whether there are any differences along the NW–SE gradient. In addition, an attempt was made to evaluate the bathymetric and zoogeographical distribution of sponges in the study area.
Methods
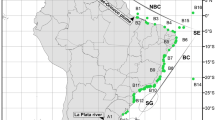
For the purposes of this study, the Aegean Sea was divided into six subareas corresponding to its main geographical divisions (Fig. 1), on the basis of its geomorphology and bathymetry, according to the data given by various authors (e.g., Valioulis and Krestenitis 1994; Stergiou et al. 1997): the northern area (A) separated from the rest of the Aegean by the N. Aegean trough, 1,600 m deep; the central-western area (B) including the Aegean coast of central Greece; the central-eastern area (C) including the Turkish coasts, the Lemnos plateau and the Chios Basin; the south-western area (D) where the Aegean joins the Ionian Sea, including the Cyclades plateau; the area of the Dodecanese Islands (E) where the Aegean joins the Levantine Basin; and the Cretan Sea (F), the deepest Aegean area with a maximum depth of 2,500 m. Areas A, B and C, located north of the 38° constitute the North Aegean, and areas D, E and F constitute the South Aegean. The only two areas of the Levantine Sea studied for their demosponge fauna were additionally considered: the coast of Egypt (G) and the coasts of Israel and Lebanon (H), the former separated from the latter by the Nile Delta.

Maps of the Aegean and the Levantine Sea indicating studied subareas
A variety of papers published on the Aegean and Levantine sponges (taxonomic, faunal and ecological) or general works including relevant information, listed in detail in Voultsiadou (2005), were studied. Based on this, a species presence/absence matrix was constructed. In addition, a matrix comprising semiquantitative values was prepared, by transforming the original information on the number of collected specimens per species and area (see Maldonado and Uriz 1995). Semiquantitative values were assigned according to the following criteria: 0=absent species, 1=species recorded once in an area, 2=species recorded with 2–5 specimens, 3=species recorded with 6–10 specimens, 4=species recorded with 11–20 specimens, and 5=species recorded with more than 20 specimens. These two matrices were transformed into triangle matrices using the Bray and Curtis (1957) semimetric distance and were subjected to multivariate analysis (i.e., cluster and multidimensional scaling MDS) (Field et al. 1982). SIMPER analysis was applied in order to identify the percentage contribution of each species to the overall similarity within areas and the dissimilarity among areas (Clarke and Warwick 1994). The above analyses were performed using PRIMER (Clark and Warwick 1994).
For the evaluation of the vertical distribution of sponges, depths were recorded for each species in each area of the Aegean Sea according to the literature. Depth distribution of the Levantine species was not analyzed due to the restricted information available. Linear regression analysis (Zar 1999) was used to identify relations between number of species and depth; for this analysis, numbers of species were grouped per 20-m depth interval. The depth range of all species occurrences (0–240 m) was further subdivided into 40-m intervals and the matrix species per depth interval was also subjected to multivariate analysis as described above.
Finally, the total of the Aegean and Levantine sponges were assigned to five zoogeographical categories according to their distribution as derived from the relevant literature: endemic species (E) distributed exclusively in the Mediterranean, Atlantic–Mediterranean species (AM) distributed in the Mediterranean and the eastern Atlantic, amphi-Atlantic species (AA) distributed in the Mediterranean and both sides of the Atlantic, Indo–Mediterranean species (IM) distributed in the Mediterranean and the Indo–Pacific, and species with a wide distribution or cosmopolitan species (C) distributed at least in the Atlantic and the Indo–Pacific.
Results
Species number and species abundance (in semiquantitative terms) per area are shown in Appendix. The sponge fauna of the Aegean Sea totals 200 species, 96 (48%) of which occur only in the North Aegean, 34 (17%) only in the South Aegean, and 70 (35%) in both sectors (Table 1). Eighty-five species have been reported from the Levantine Sea, 32 of which were not found in the Aegean. An estimation of the number of publications existing for each area showed that of a total of 48 publications referring to the Aegean, 31 included data on the sponge fauna of the northern sector and 23 on that of the southern sector (Table 1). Nine publications concerned the sponges of the Levantine Sea. The number of relevant publications was positively correlated with the number of species known per area (r= 0.902, P< 0.05, n= 8) (Fig. 2).

Relationship between species number (black circles) and degree of exploration (open circles) in the Aegean and Levantine subareas (see Fig. 1)
The most frequently reported demosponges in both sectors of the Aegean (see Appendix) were the species Agelas oroides, Geodia cydonium, Petrosia ficiformis, Spongia officinalis, Scalarispongia scalaris and Sarcotragus foetidus. Species found in all subareas of the North Aegean, but not reported from the South Aegean are: Halisarca dujardini, Oscarella lobularis, Pachastrella monilifera, Plakortis simplex, Pseudosuberites hyalinus, Stelletta grubii, Suberites carnosus, S. ficus and Ulosa stuposa. The species Chondrilla nucula, Cliona celata and Mycale massa, which are very common in the North Aegean, have been found only once in the South Aegean. The majority of the 37 species found exclusively in the southern sector were reported in only one of the three subareas. Only 20 out of the 85 species recorded in the Levantine Sea occur on the coasts of both Egypt and Israel (see Appendix Table 5).
Cluster analysis (Fig. 3a) performed for the semiquantitative data indicated, at a similarity level of 26.5%, two distinct groups of areas: one for the Levantine Sea and another for the Aegean Sea. Furthermore, the Aegean was separated into two groups (similarity 35%): one including the North Aegean subareas and another with the South Aegean subareas. The results of MDS confirmed the above grouping (Fig. 3b). The application of multivariate analysis on the presence/absence matrix provided the same grouping (results not shown), with the only exception being the Cretan Sea (F), which formed a group by itself within the Aegean Sea group.

Cluster analysis (a) and MDS (b) showing relationships among the Aegean and Levantine subareas
The species responsible for the similarity in each of the three groups are shown in Table 2. Overall, 19 species contributed 50% to the similarity of the North Aegean, each participating with almost similar percentages (from 2.00 to 3.33). In-group similarity of the South Aegean areas was mostly attributed to the participation of three highly contributing species (10.07% each). Dictyoceratida species, i.e., the two common commercial species Spongia officinalis and Hippospongia communis and the species Scalarispongia scalaris and Sarcotragus foetidus are among the eight species contributing 50% to the similarity of the South Aegean. In-group 50% similarity of the Levantine group was attributed to six species with relatively high contributions each.
The allocation of sponge species occurring in the Aegean Sea to the different zoogeographical categories (Table 1, Appendix Table 4) was: 111 Atlantic–Mediterranean species (55.5%), 59 endemic species (29.5%), 27 cosmopolitan species (13.5%) and 3 amphi-Atlantic species (1.9%). These values did not generally differ between the North and South Aegean (Table 1). The Levantine Sea (Table , 1, Appendix Table 5) exhibited a similar participation of the endemic component (30%), but a decrease in the percentage of Atlantic–Mediterranean (46.5%) and an increase in the percentage of cosmopolitan species (17.5%). Four species (4.8%) with an Indo–Mediterranean distribution also occurred in this area.
The allocation of species to the main orders of the class Demospongiae (Table 3, 4, 5) showed that the richest order, in terms of species number in the Aegean Sea, was Poecilosclerida (24% of the total species) followed by the Hadromerida (20%). In the Levantine Sea, Halichondrida prevailed (22%), while Poecilosclerida, Dictyoceratida and Haplosclerida had an almost equal participation in the total demosponge fauna of this area. A gradual decrease in the contribution of Poecilosclerida, Hadromerida and Astrophorida and a corresponding increase of Halichondrida and Haplosclerida from the North Aegean to the Levantine was observed. The horny sponges (Dictyoceratida) had their lowest participation in the fauna of the North Aegean (14% of the total species of the area) and the highest in that of the South Aegean (17%).
With respect to depth, out of the 200 species found in the Aegean Sea, 158 (79%) were found in the sublittoral (0–40 m), 112 (56%) in the circalittoral zone (40–120 m), and 34 (17%) in the bathyal zone (deeper than 120 m) (Table 4). It should be mentioned that 26 species recorded in the upper sublittoral zone (13% of the total number of Aegean species) were found in submarine semidark or dark caves (Table 4). Regression analysis (Fig. 4) showed that the number of species was a negative function of depth (r2=0.954, P<0.05). Moreover, 116 species (58%) were restricted in one of the three depth zones, 19 species (9.5%) occurred at all depths and 65 species (32.5%) were distributed in two out of the three zones studied. Cluster analysis performed for species presence/absence at 40-m depth intervals (Fig. 5a) revealed two distinct groups of depth zones at a similarity level of 27.2%, one of them further separated in two groups (similarity 57.1%). These three groups correspond to the three depth zones of the Mediterranean: sublittoral, circalittoral and bathyal. The results of MDS confirmed the above grouping (Fig. 5b). In the Levantine, 88% of the species were found in the sublittoral zone (Table 5). A considerable number of species (22.3%), however, were living in caves.

Regression analysis of depth and number of demosponge species found at 20-m depth intervals in the Aegean Sea

Cluster analysis (a) and MDS (b) showing relationships among depth zones (depth in m) in the Aegean Sea
Discussion
To date, the demosponge fauna of the Aegean Sea, counting up to 200 species, constitutes 37% of the total Mediterranean demosponges (Voultsiadou 2005). Thus, the Aegean Sea ranks third in the Mediterranean in terms of demosponge diversity, following the western Mediterranean (80%) and the Adriatic (40%), although until recently its sponge fauna was considered underestimated (Pansini et al. 2000). This scaling agrees with that for the Mediterranean octocorals (Vafidis et al. 1994) and amphipods (Stefanidou and Voultsiadou 1995), but other groups such as decapods (Koukouras et al. 1992), ascidians (Koukouras et al. 1995), gastropods (Koutsoubas et al. 1997) and polychaetes (Arvanitidis et al. 2002) are represented by higher species numbers in the Aegean than in the Adriatic Sea.
The demosponge fauna known up-to-date from the Aegean and Levantine Sea totals 284 species. A decrease in demosponge species numbers is apparent from the North Aegean to the Levantine Sea. The majority (58.5%) of the species has been found in the North Aegean, 36.6% in the South and 30% in the Levantine basin. This was related to the research effort devoted to the three areas. The same is also true for the bivalve species of the Greek seas (Zenetos 1997). Although further research could possibly reduce the differences in species numbers among areas, the overall picture is not expected to change since it reflects the different conditions prevailing in these areas discussed in detail below.
Multivariate analysis indicated a clear separation of the Levantine from the Aegean Sea. The former, which is the oldest part of the Mediterranean, has been described as an extremely stressed environment having high salinities and temperatures, defined by an annual mean isotherm of 20°C and a surface isohaline of 39 ppt (Por 1978). Its severe oligotrophic conditions have been pointed out in order to explain the impoverished species diversity and the small sizes of the Levantine biota (e.g., Por and Dimentman 1989). This “nanism” was observed in sponge specimens found on the coasts of Egypt (Burton 1936) and Israel (Lévi 1956). Spicules of smaller size were observed by Carteron (2002) in demosponges from the Lebanon coast, in comparison to specimens from the western Mediterranean. Furthermore, the Levantine fauna has been recently receiving the influence of the Lessepsian migration phenomenon (Por and Dimentman 1989), which is also reflected in its sponge diversity (see below).
The Aegean Sea was further separated into two groups, one for the North Aegean and another for the South Aegean, by multivariate analysis. This grouping is again consistent with the differences in the geographical, physical, chemical and biological characteristics of the two areas. The North Aegean coasts are in their greatest length continental and receive high river inflow amounts, while the South Aegean ones include open areas and a plethora of islands. Moreover, the North Aegean is subjected to the strong influence of the low-salinity and cooler Black Sea water and has, in comparison to the South Aegean, lower temperature and salinity values, but higher phytoplankton concentration, chlorophyll a and PLP (phytoplankton-like pigments) values as well as higher zooplankton abundance comparable to that of other Mediterranean regions (Valioulis and Krestenitis 1994; Stergiou et al. 1997, etc.). On the other hand, the South Aegean receives the salty Levantine Intermediate Water, carrying a limited supply of nutrients and is generally characterized as one of the most oligotrophic parts of the Mediterranean Sea (Valioulis and Krestenitis 1994; Stergiou et al. 1997; Psarra et al. 2000, etc.). Such a differentiation agrees with Bianchi and Morri (2000) who, discussing the Mediterranean marine biodiversity, distinguish the North Aegean as one of the ten biogeographical sectors of the Mediterranean Sea and include the South Aegean in one sector along with the Levantine Basin. The above authors observed a significant qualitative impoverishment of benthos in the South Aegean (Bianchi and Morri 1983). The effect of nutrient availability on the differences in composition and diversity of benthic communities between continental margins and oceanic islands in tropical areas has been shown by various authors according to Taylor (1997). Dissimilarity between North and South Aegean island coasts on the basis of a 64 sponge species presence/absence has been previously calculated (Kefalas et al. 2003), while no differentiation in bivalve diversity was found corresponding to a geographical pattern (Zenetos 1997). Sponges, as sessile filter feeding organisms, strongly reflect differences in the abiotic environment and are considered as live indicators of environmental characteristics (Carballo et al. 1996).
The observed dissimilarity between the two sectors of the Aegean has been attributed to a large number of species appearing very frequently in the northern sector, while being absent or present in only low frequencies in the southern sector. The impoverishment of the South Aegean was further supported by the fact that its in-group similarity was the result of the contribution of a small number of very common and widely distributed or commercial Mediterranean species. The same was true for the Levantine where, additionally, four Lessepsian migrants (Cinachyrella australiensis, Geodia micropunctata, Lissodendoryx schmidti and Hyrtios erecta) contributed, of which Cinachyrella australiensis was among the first six highly contributing species.
The analysis of the distribution of the zoogeographical categories showed that most of the species occurring in the Aegean and the Levantine Sea were Atlanto–Mediterranean, followed by the endemics, the cosmopolitan and the few amphi-Atlantic species. This ranking disagrees with that given for the Mediterranean demosponges by Pansini (1992) who calculated a composition of 45.7% endemics, 38% Atlantic–Mediterranean and 10% cosmopolitan species. These percentages stand, more or less, for the western Mediterranean demosponges since 80% of the Mediterranean species have been found in this area (Pansini 1996), an area that supports the most “typical” Mediterranean fauna (Bianchi and Morri 2000). One possible reason for the reduced number of endemic sponges recorded in the Aegean up-to-date is the lack of research in deep waters and special habitats (e.g., caves). Two of the recently described species, Coscinoderma sporadense (Voultsiadou et al. 1991) and Hemiasterella aristoteliana (Voultsiadou and van Soest 1991), were found in semidark caves of the North Aegean. Pansini (1992) indicated that research in waters deeper than 100 m increased the number of the known endemic sponges in the western Mediterranean during 1980. On the other hand, recent works (e.g., Carballo et al. 1997) suggest that some species previously characterized as Mediterranean endemics, should be considered Atlantic–Mediterranean, thus decreasing the endemic component. Above all, it is possible that sponges follow the general west–east gradient of the endemic species numbers in the Mediterranean: 77.5% of the total in the Western Mediterranean, 34.6% in the Adriatic Sea and 22.5% in the Eastern Mediterranean (Fredj et al. 1992). This gradient was evident from the comparison of zoogeographical categories in the Aegean and the Levantine. The stressing conditions and their lability over short time-spans did not allow for speciation in this eastern part of the Mediterranean (Por and Dimentman 1989). Various invertebrate groups demonstrate this trend of decreasing endemism in the Mediterranean from west to east (Koukouras et al. 2001). However, Arvanitidis et al. (2002) did not find differences in the distribution of the zoogeographical categories of polychaetes among the Mediterranean areas. It is worth mentioning that none of the four Indo–Mediterranean species that entered the Levantine basin as lessepsian migrants (Tsurnamal 1969b) have been up-to-now found in the Aegean Sea. The very high contribution of the Atlantic–Mediterranean component in the Aegean and the Levantine confirms that the exchange of faunal elements between the Atlantic and the Mediterranean extends to the eastern part of the latter (Pansini 1992; Koukouras et al. 2001). The various zoogeographical categories do not seem to exhibit important differences in the two sectors of the Aegean possibly due to the small size of this area.
The ordinal distribution of demosponges in the three study areas indicated a NW–SE gradient, which is in accordance with the general aspect that sponges with skeletons rich in spongin are well distributed in warm waters, being replaced at high latitudes or deeper waters by sponges with siliceous skeletons (Vacelet 1988). The preference of keratose sponges for shallow and warm habitats has been attributed to the inhibition of their larval dispersal and settlement success by the decrease in temperature (Maldonado and Young 1996). Out of the 56 genera recorded in the Levantine (Table 5), two (Gastrophanella and Microscleroderma) have a tropical distribution (Perez et al. 2004), while at least 32 (e.g., Agelas, Alectona Chelonaplysilla, Chondrilla, Chondrosia, Didiscus, Erylus, Spongia) have a distinct tropical-subtropical diversity center, lacking or rare in cold water areas (estimated according to van Soest 1994).
An estimation of demosponge diversity at the order level from two species lists, one for the Italian coasts (Pansini 1995) and another for the European coasts (van Soest 2001), given in Table 3, permitted a comparison with the Aegean and Levantine data: Poecilosclerida, the most diverse of all poriferan orders, both in species numbers and in morphology (Hooper and van Soest 2002), demonstrates an obvious gradual decrease in species numbers from NW to SE. This decrease may possibly be attributed to the fact that major families and genera of this order such as Myxillidae, Crellidae, Raspaillidae, Mycale, Tedania, Hymedesmia Phorbas, Clathria (van Soest 1994, etc.), show a distinct diversity center in cold and deep waters or have a worldwide distribution. On the other hand, an increasing tendency in the contribution of the orders Dictyoceratida, Halichondrida, and Homosclerophorida is observed. These orders are highly represented in the Mediterranean by genera or families such as Ircinia, Dysidea, Spongiidae, Acanthella, Axinella, Myrmekioderma, Plakina, Corticium, mostly distributed in warm temperate or tropical environments (van Soest 1994).
Multivariate analysis of the bathymetric distribution of the Aegean sponges showed a clear distinction of the three depth zones: sublittoral, circalittoral and bathyal. The number of species was negatively correlated with depth, partly reflecting the intense research activities in the shallow waters of this area and the poor knowledge of the deeper waters (Pansini 1992). It is generally accepted that in the western Mediterranean, the richest zone in demosponges is the circalittoral (Vacelet 1988; Maldonado and Uriz 1995). Many of the sciaphilic species of this zone, however, also live in dark sites of the shallow sublittoral (Posidonia rhizomes, algae, rock crevices, caves, etc.), a very common case in the Aegean Sea. Demosponge distribution in the sublittoral zone has been connected to the level of direct solar irradiance and the algal competition, rather than to depth (Uriz et al. 1992; Pansini 1997). Moreover, Maldonado and Uriz (1995) showed that outside the Mediterranean, in the Azores and the Canary Islands, the medium sublittoral zone exhibits maximal species richness, whereas faunal impoverishment occurs in the lower sublittoral and circalittoral levels. The above authors claim that the general pattern of biotic affinity is determined predominantly by horizontal (geographic) distances while the role of vertical distance (bathymetry) is much less significant.
References
Arvanitidis C, Bellan G, Drakopoulos P, Valavanis V, Dounas C, Koukouras A, Eleftheriou A (2002) Seascape biodiversity patterns along the Mediterranean and the Black Sea: lessons from the biogeography of benthic polychaetes. Mar Ecol Prog Ser 244:139–152
Azov Y (1991) Eastern Mediterranean-a marine desert? Mar Pollut Bull 23:225–232
Bianchi CN, Morri C (1983) Note sul benthos marino costiero dell’ isola di Kos (Egeo sud-orientale). Natura (Milan) 74:96–114
Bianchi CN, Morri C (2000) Marine biodiversity of the Mediterranean Sea: situation, problems and prospects for future research. Mar Pollut Bull 40:367–376
Bray RJ, Curtis JI (1957) An ordination of the upland forest communities of southern Wisconsin. Ecol Monogr 27:325–349
Burton M (1936) The fishery grounds near Alexandria. IX. Sponges. Notes Mem Fish Res Cairo 17:1–28
Carballo JL, Naranjo SA, Garcia-Gomez JC (1996) Use of marine sponges as indicators in marine ecosystems at Algeciras Bay (southern Iberian Peninsula). Mar Ecol Prog Ser 135:109–122
Carballo JL, Naranjo SA, Garcia-Gomez JC (1997) Where does the Mediterranean Sea begin? Zoogeographical affinities of the littoral sponges of the Straits of Gibraltar. J Biogeogr 24:223–323
Carteron S (2002) Etude taxonomique des spongiaires du Liban. Stage de Maîtrise, Centre d’ Océanologie de Marseille
Clarke KR, Warwick RM (1994) Change in marine communities: an approach to statistical analysis and interpretation. Nat Envir Res Coun Plym Mar Lab Plym
Field JG, Clarke KR, Warwick RM (1982) A practical strategy for analyzing multispecies distribution patterns. Mar Ecol Prog Ser 8:37–52
Fredj G, Bellan-Santini D, Meinardi M (1992) Etat des connaissances sur la faune marine méditerranéenne. Bull Inst Oceanogr no special 9:133–145
Hooper JNA, Soest van RWM (2002) Order Poecilosclerida Topsent. In: Hooper JNA and van Soest RWM (eds) Systema Porifera. A guide to the classification of sponges. Kluwer/Plenum, New York, pp 403–408
Ilan M, Ben-Eliahu MN, Galil BS (1994) Three deep water sponges from the Eastern Mediterranean and their associated fauna. Ophelia 39:45–54
Kefalas E, Tsirtsis G, Castritsi-Catharios I (2003) Distribution and ecology of Demospongiae from the circalittoral of the islands of the Aegean Sea (Eastern Mediterranean). Hydrobiologia 499:125–134
Koukouras A, Dounas C, Turkay M, Voultsiadou-Koukoura E (1992) Decapod crustacean fauna of the Aegean Sea: new information, check-list, affinities. Senckenb Marit 22:217–244
Koukouras A, Voultsiadou-Koukoura E, Kevrekidis T, Vafidis D (1995) Ascidian fauna of the Aegean Sea with a check list of the Eastern Mediterranean and Black Sea species. Ann Inst Océanogr Paris 71:19–34
Koukouras A, Voultsiadou E, Kitsios MS, Doulgeraki S (2001) Macrobenthic fauna diversity in the Aegean Sea, affinities with other Mediterranean regions and the Black Sea. Bios (Macedonia, Greece) 6:61–76
Koutsoubas D, Koukouras A, Voultsiadou-Koukoura E (1997) Prosobranch mollusc fauna of the Aegean Sea: new information, checklist, distribution. Isr J Zool 43:19–54
Lévi C (1956) Spongiaires des côtes d’ Israel. Bull Res Counc Israel 6B:201–212
Maldonado M, Uriz MJ (1995) Biotic affinities in a transitional zone between the Atlantic and the Mediterranean: a biogeographical approach based on sponges. J Biogeogr 22:89–110
Maldonado M, Young CM (1996). Effects of physical factors on larval behavior, settlement and recruitment of four tropical demosponges. Mar Ecol Prog Ser 138:169–180
Muricky G, Boury-Esnault N, Bézak Ch, Vacelet J (1998) Taxonomic revision of the Mediterranean Plakina Schulze (Porifera, Demospongiae, Homoscleromorpha). Zool J Linn Soc 124:169–203
Pansini M (1992) Considérations biogéographiques et systématiques pour une mise à jour des données sur le peuplement de spongiaires mediterranéens. Bull Inst Oceanogr 9:43–51
Pansini M (1995) Porifera. In: Minelli A, Ruffo S, La Posta S (eds) Checklist delle specie della fauna italiana. Calderini, Bologna 2:1–38
Pansini M (1996) Considerazioni sulla biodiversità dei poriferi dei mari italiani e del Mediterraneo. Biol Mar Medit 3:128–135
Pansini M (1997) Effeti della luce sulla morfologia, distribuzione ed ecologia di alcune demosponge mediterrenee. Biol Mar Medit 4:74–80
Pansini M, Morri C, Bianchi CN (2000) The sponge community of a subtidal area with hydrothermal vents: Milos Island, Aegean Sea. Est Coast Shelf Sci 51:627–635
Perez T, Vacelet J, Bitar G, Zibrowius H (2004) Two new lithistids (Porifera: Demospongiae) from a shallow Mediterraenan cave (Lebanon). J Mar Biol Ass UK 84:15–24
Por FD (1978) Lessepsian migration. The influx of Red Sea biota into the Mediterranean by way of the Suez Canal. In: Billings WB, Goley F, Lange OL, Olson JS (eds) Ecological studies, book 23. Springer, Berlin Heidelberg New York, p 228
Por FD, Dimentman C (1989) The legacy of Tethys: an aquatic biogeography of the Levant. In: Dumont J, Wergel A (eds) Monographiae biologicae. Kluwer, Dordrecht, 63:1–214
Psara S, Tselepidis A, Ignatiades L (2000) Primary production in the oligotrophic Cretan Sea (NE Mediterranean): seasonal and interannual variability. Prog Oceanogr 46:187–204
Soest van RW (1994) Demosponge distribution patterns. In: Van Soest RMW, van Kempen TMG, Braekman J-C (eds) Sponges in time and space. Balkema, Rotterdam, pp 213–223
Soest van RWM (2001) Porifera. In: Costello MJ, Emblow C, White R (eds) European register of marine species. A check-list of the marine species in Europe and a bibliography of guides to their identification. Publ Scient M.N.H.N. Paris, pp 85–103
Stefanidou D, Voultsiadou-Koukoura E (1995) An account of our knowledge of the Amphipoda fauna of the Aegean Sea. Crustaceana 68:597–615
Stergiou KI, Christou ED, Georgopoulos D, Zenetos A, Souvermezoglou C (1997) The Hellenic Seas: physics, chemistry, biology and fisheries. Oceanogr Mar Biol Annu Rev 35:415–538
Taylor JD (1997) Diversity and structure of tropical Indo-Pacific benthic communities: relation to regimes of nutrient input. In: Ormond RFG, Gage J, Angel MV (eds) Marine biodiversity. Cambridge University Press, London, pp 178–200
Tsurnamal M (1967) Chelonaplysilla erecta, n. sp. (Demospongiae, Keratosa) from Mediterranean coast of Israel. Isr J Zool 16:96–100
Tsurnamal M (1969a) Four new species of Mediterranean demospongiae and new data on Callites lacazei Schmidt. Cah Biol Mar 10:343–357
Tsurnamal M (1969b) Sponges of Red Sea origin on the Mediterranean coast of Israel. Isr J Zool 18:149–155
Uriz MJ, Rosell D, Martin D (1992) The sponge population of the Cabrera Archipelago (Balearic Islands): characteristics, distribution and abundance of the most representative species. PSZNI Mar Ecol 13:101–117
Vacelet J (1988) Indications de profondeur données par les Spongiaires dans les milieux benthiques actuels. Geol Mediterr 15:13–26
Vafidis D (2002) Biogeography of the Anthozoa coelenterate fauna of the Mediterranean Sea (Abstr ICZEGAR)
Vafidis D, Koukouras A, Voultsiadou-Koukoura E (1994) Octocoral fauna of the Aegean Sea with a checklist of the Mediterraenean species: new information, faunal comparisons. Ann Inst océanogr Paris 70:217–229
Valioulis IA, Krestenitis YN (1994) Modelling the water mass circulation in the Aegean Sea. Part I: wind stresses, thermal and haline fluxes. Ann Geophys 12:794–807
Voultsiadou E (2005) Sponge diversity in the Aegean Sea: checklist and new information. Ital J Zool 72(1):53–64
Voultsiadou-Koukoura E, van Soest RWM (1991) Hemiasterella aristoteliana n. sp. (Porifera, Hadromerida) from the Aegean Sea with a discussion of the family Hemiasterellidae. Bijdr Dierk 61:43–49
Voultsiadou-Koukoura E, van Soest RWM, Koukouras A (1991) Coscinoderma sporadense sp. n. from the Aegean Sea with comments on Coscinoderma confragosum (Porifera, Dictyoceratida). Zool Scr 20:195–199
Zar JH (1999) Biostatistical analysis, 4th edn. Prentice-Hall, Englewood Cliffs, NJ
Zenetos A (1997) Diversity of Bivalvia in Greek waters: effects of geography and environment. J Mar Biol Ass UK 77:463–472
Acknowledgements
I thank C. Antoniadou, D. Vafidis, C. C. Chintiroglou, A. Koukouras and S. Gkelis for their help in different stages of this work, as well as K. I. Stergiou for his valuable and constructive suggestions.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by H.-D. Franke
Appendix
Appendix
Rights and permissions
About this article
Cite this article
Voultsiadou, E. Demosponge distribution in the eastern Mediterranean: a NW–SE gradient. Helgol Mar Res 59, 237–251 (2005). https://doi.org/10.1007/s10152-005-0224-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10152-005-0224-8