
Abstract
Clostridium perfringens (C. perfringens) infection is recognized as one of the most challenging issues threatening food safety and perplexing agricultural development. To date, the molecular mechanisms of the interactions between C. perfringens and the host remain poorly understood. Here, we show that stimulator of interferon genes (STING)-dependent trained immunity protected against C. perfringens infection through mTOR signaling. Heat-killed Candida albicans (HKCA) training elicited elevated TNF-α and IL-6 production after LPS restimulation in mouse peritoneal macrophages (PM). Although HKCA-trained PM produced decreased levels of TNF-α and IL-6, the importance of trained immunity was demonstrated by the fact that HKCA training resulted in enhanced bacterial phagocytic ability and clearance in vivo and in vitro during C. perfringens infection. Interestingly, HKCA training resulted in the activation of STING signaling. We further demonstrate that STING agonist DMXAA is a strong inducer of trained immunity and conferred host resistance to C. perfringens infection in PM. Importantly, corresponding to higher bacterial burden, reduction in cytokine secretion, phagocytosis, and bacterial killing were shown in the absence of STING after HKCA training. Meanwhile, the high expression levels of AKT/mTOR/HIF1α were indeed accompanied by an activated STING signaling under HKCA or DMXAA training. Moreover, inhibiting mTOR signaling with rapamycin dampened the trained response to LPS and C. perfringens challenge in wild-type (WT) PM after HKCA training. Furthermore, STING‑deficient PM presented decreased levels of mTOR signaling-related proteins. Altogether, these results support STING involvement in trained immunity which protects against C. perfringens infection via mTOR signaling.
Similar content being viewed by others


Introduction
Clostridium perfringens (C. perfringens) is an opportunistic pathogen of humans and livestock, causing a range of serious enteric and histotoxic infections, including gas gangrene, enteritis/enterocolitis, and enterotoxemia [1]. In addition, C. perfringens can be present as a contaminant in meat, vegetables, and raw milk products. Therefore, it is frequently associated with foodborne outbreaks and classified as the second most prevalent etiological factor of bacterial food poisoning in the United States, Europe, and many other regions [2]. Traditional toxoid vaccines and antibiotics are effective countermeasures in combating C. perfringens infection. With the drug resistance spectrum expanding and waning vaccine immunity, the demand for novel control strategies is even more pressing.
The innate immune response serves as the initial barrier of host defense against pathogenic infections. Research on the anti-bacterial innate immune response is beneficial for finding the intervention target. According to recent studies, the innate immune cells (such as monocytes, macrophages, NK cells, and neutrophils) can achieve memory characteristics after primary stimulation and mount host response to restimulation, a phenomenon known as trained immunity or innate immune memory, accompanied by the epigenetic and metabolic reprogramming [3]. In contrast to adaptive immunity, trained immunity is nonspecific, rapid, modest, and exists for a relatively short time. One notable feature of trained immunity is the occurrence of the Warburg effect, wherein cells undergo a metabolic transition from oxidative phosphorylation to aerobic glycolysis. The PI3K/Akt/mTOR pathway and the TCA cycle appear to be the common denominators in this process, and their interplay influences the regulation of histone acetylation and methylation in the promoters and enhancers of genes responsible for encoding inflammatory cytokines [1]. Many studies have reported evidence for trained immunity in plants, invertebrates, and vertebrates [4]. Trained immunity can be activated by diverse stimuli, such as β-glucan, LPS, LTA, BCG, a Western diet, and oxLDL [1, 5,6,7,8,9,10,11]. It is like a double‐edged sword. The induction of trained immunity could both promote increased susceptibility to secondary infections and contribute to the progression of the inflammatory disorder [12]. Precisely due to the memory property, trained immunity provides protection against subsequent heterologous pathogenic infections, including Mycobacterium tuberculosis, Candida albicans, Leishmania braziliensis, and influenza A virus [13,14,15]. Further elucidation in the field of trained immunity is likely to open new avenues for the novel preventive and therapeutic strategies of host resistance to C. perfringens infection.
Germline-encoded pattern-recognition receptors (PRR) act as “sensors” of trained immunity. At present, only a proportion of the NOD-like receptors, the Toll-like receptors, and the C-type lectin receptors have been characterized for their roles in trained immunity [6, 16,17,18]. Nevertheless, the function of nucleic acid receptors and related signaling proteins has not yet been determined during the production of trained immunity. The adaptor protein stimulator of interferon genes (STING) has been widely investigated for its role in DNA sensing. Its function primarily revolves around the activation of type I interferons (IFN), which are mainly involved in the development of infectious diseases, autoimmune diseases, and cancer [19]. Its role in regulating trained immunity and the impact of STING-mediated trained immunity on antibacterial infection remains elusive. Herein, we demonstrate that STING-dependent trained immunity contributes to host defense against C. perfringens infection via mTOR signaling.
Materials and methods
Mice
C57BL/6 J wild-type (WT) mice and STING−/− mice were procured from Jackson Laboratory. The mice were provided with sterilized food and water, and were subjected to a strict 12 h light cycle. They were housed in groups of up to 6 mice per cage. Age-and sex-matched mice were used for all experiments. All animal studies were conducted in accordance with the approved experimental practices and standards set by the Animal Welfare and Research Ethics Committee at Jilin University (KT202202182).
Trained immunity in vitro model
Peritoneal macrophages (PM) were obtained from WT mice at 3 days after mice were injected with 4% thioglycollate broth (Sigma-Aldrich, #70157). The PM were distributed in 96-well cell culture plates, each well containing 2 × 105 cells, or in 6-well cell culture plates, each well containing 3 × 106 cells. Subsequently, the PM were stimulated using RPMI1640 medium (Gibco, #31800–022), DMXAA (50 ug/mL, Sigma-Aldrich, #D5817), or heat-killed Candida albicans (HKCA, 1 × 105 cells/mL) for 24 h. Then, cells were washed and rested for 5 days in the culture medium with 10% FBS (Gibco, #A31608-02). On day 6, a final wash was conducted, followed by stimulation with medium, 100 ng/mL of LPS (Sigma-Aldrich, #L6529), and C. perfringens (ATCC13124, MOI = 5). The cell supernatants and lysates were subsequently collected to perform ELISA or Western blotting assays.
Trained immunity in vivo model
Age- and sex-matched WT and STING−/− mice were subjected to training with two intraperitoneal (i.p.) injections of 1 × 106 cells of HKCA on days −7 and −4. Phosphate-buffered saline (PBS) was used as the control. For in vitro experiments, mice were injected i.p. with 4% thioglycolate broth on day −3. PM were collected and stimulated on day 0 and day 1, respectively. For in vivo experiments, mice were subjected to intramuscular infection with a concentration of 2 × 107 colony-forming units (CFU) of C. perfringens diluted in PBS in a total volume of 100 μL on day 0. After 24 h post-infection, the mice were euthanized to obtain infected leg muscle samples for the purpose of quantifying bacterial load. Additionally, the severity of gas gangrene was evaluated using a scoring system that had been previously described in a study [20].
Phagocytosis and intracellular killing assays
PM were infected with C. perfringens at a multiplicity of infection (MOI: 5). Following infection, the PM were centrifuged at 515 × g for 2 min to ensure synchronous infection. Subsequently, the cells were incubated at 37 °C for 1 h and washed twice with RPMI-1640 medium. The PM were then cultured in RPMI-1640 medium supplemented with 200 U/mL penicillin, 200 U/mL streptomycin, and 100 μg/mL gentamicin to eliminate excessive extracellular bacteria for 1 h. Finally, the PM were incubated at 37 ℃ for the specified time intervals. Killing efficiency was calculated as ([CFU/mL at 2 h] − [CFU/mL at 4 h])/(CFU/mL at 2 h).
Real‑time PCR
RNA was extracted using TRI reagent (Sigma-Aldrich, #T9424) and converted into cDNA. Subsequently, real-time PCR assays were conducted using SYBR Green (Roche, #4913914001) on an ABI Prism 7500 sequence detection system (Life Tech [Applied BioSystems], Waltham, USA). The Agilent integrity check was used to verify RNA quality. Gene expression levels were determined using the 2−ΔCt method. The following primer sequences were utilized: β-actin sense 5′-CGTGGGCCGCCCTAGGCACCA-3′ and antisense 5′-TTGGCCTTAGGGTTCAGGGGGG-3′. IFN-β sense 5′-ACTGCCTTTGCCATCCAAGA-3’ and antisense 5’-CACTGTCTGCTGGTGGAGTT-3’. CXCL10 sense 5′-ATCCCTGCGAGCCTATC-3’ and antisense 5′-GCCATCCACTGGGTAAA-3′.
Cytokine measurements
The supernatants were used for ELISA measurements according to the manufacturer’s instructions. Mouse TNF-α and IL-6 ELISA kits were purchased from R&D Systems (#DY410 and #DY406).
Western blotting analysis
PM were harvested and lysed using cold RIPA lysis buffer supplemented with complete protease inhibitor cocktail (Sigma-Aldrich, #P8340). Samples were centrifuged and supernatants were used for immunoblotting. The cell lysates were separated by SDS-PAGE and transferred to PVDF membranes (Millipore, # ISEQ00010). Following blocking with 5% nonfat milk, the membranes were blotted with antibodies against p-mTOR (Abcam, #ab109268), p-AKT (Proteintech, #66444-1-Ig), HIF1α (Abcam, #ab179483), STING (Proteintech, #19851-1-AP), p-TBK1 (Cell Signaling Technology, #5483), p-IRF3 (Cell Signaling Technology, #29047), and β-actin (Proteintech, #81115-1-RR).
Statistical analysis
The experiments were conducted independently on three occasions. The differences between mean values were assessed with Student’s t-test, one-way ANOVA with Dunnett multiple comparison test, or two-way ANOVA with Bonferroni multiple comparison test (*P < 0.05; **P < 0.01; ***P < 0.001). Data are shown as mean ± SEM. The statistical significance was determined by the software GraphPad Prism6.
Results
HKCA-induced trained immunity enhances macrophage function and promotes host defense against C. perfringens infection
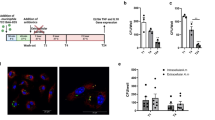
To evaluate the role of trained immunity in host defense against C. perfringens infection, an in vitro model was established for trained immunity. WT mice were trained with two i.p. injections of HKCA on days -7 and -4. PBS was used as the control. On day -3, mice were injected i.p. with 4% thioglycolate broth to induce macrophage differentiation and accumulation. Then PM were collected and restimulated (LPS or C. perfringens) on day 0 and day 1, respectively (Figure 1A). We found that HKCA training induced a significant increase in the concentrations of TNF-α and IL-6 upon LPS stimulation (Figure 1B, C). Unlike LPS, there was a lower production of TNF-α and IL-6 response to C. perfringens infection under HKCA training compared with the control group (Figure 1D, E). Based on the aforementioned results, we further analyzed the phagocytosis and bacterial killing ability of PM before and after HKCA training. Interestingly, HKCA training resulted in enhanced bacterial phagocytic ability and clearance (Figure 1F, G). Collectively, these results suggest that HKCA-induced trained immunity enhances macrophage function and promotes host defense against C. perfringens infection.

Trained immunity protects the host against C. perfringens infection. A Flow chart of in vitro trained immunity experimental procedure. (B−E) TNF-α and IL-6 production was measured in the supernatants of mouse PM response to LPS or C. perfringens stimulation according to A (n = 3 independent experiments). F, G Phagocytosis and intracellular killing of C. perfringens was detected by CFU enumeration. Data are shown as mean ± SEM. Data were pooled from 3 independent experiments. Statistical significance is indicated by *p < 0.05, **p < 0.01, and ***p < 0.001.
Trained immunity activates STING signaling
STING is recognized as a significant adaptor protein of DNA sensors. Its role in regulating trained immunity and the impact of STING-mediated trained immunity on antibacterial infection remains elusive. To investigate STING signaling activation during the process of trained immunity, we detected the protein levels of STING, p-TBK1, and p-IRF3 before and after stimulation with LPS or C. perfringens. The results show that STING signaling was upregulated in all HKCA-trained samples compared with their controls (Figure 2A–F). Meanwhile, the expression levels of CXCL10 and IFN-β were significantly increased in HKCA training PM compared with the control cells after C. perfringens infection (Figure 2G , H). These results suggest that trained immunity induces STING signaling activation.

Trained immunity activates STING signaling. WT PM were trained with HKCA and then were restimulated with LPS or C. perfringens at an MOI of 5 for the indicated times. A, D Whole cell lysates were analyzed for STING, p-TBK1, p-IRF3, and β-actin by Western blotting. B, C, E, and F The gray intensity value of proteins was calculated using ImageJ software. G IFN-β mRNA levels were measured at 4 h post‑infection by qRT‑PCR. H CXCL10 mRNA levels were measured at 24 h post‑infection by qRT‑PCR. Data are shown as mean ± SEM. Data were pooled from 3 independent experiments. Statistical significance is indicated by *p < 0.05, **p < 0.01, and ***p < 0.001.
STING agonist DMXAA is a strong inducer of trained immunity and confers host resistance to C. perfringens infection in macrophages
To further examine the potential involvement of STING in trained immunity, we trained PM with STING agonist DMXAA for 24 h and rested for 5 d to induce macrophage differentiation. HKCA and PBS served as positive and negative control, respectively. Subsequently, the cytokine production was measured after treatment with LPS (Figure 3A). We found that training with DMXAA showed a higher production of IL-6 and TNF-α upon LPS stimulation compared with the control group, nearly to the extent observed in the HKCA training group. (Figure 3B, C). We further characterized the phagocytosis and the bacterial killing ability of PM before and after DMXAA training. Expectedly, DMXAA training resulted in enhanced bacterial phagocytic ability and clearance (Figure 3D , E). These results indicate that DMXAA is a strong inducer of trained immunity and confers host resistance to C. perfringens infection in macrophages.

STING agonist DMXAA directly induces trained immunity. WT PM were trained with DMXAA or HKCA for 24 h and rested for 5 days. WT PM were restimulated with LPS or C. perfringens at an MOI of 5 for the indicated times. A Flow chart of in vitro trained immunity experimental procedure. B, C TNF-α and IL-6 production were measured in the supernatants of mouse PM response to LPS stimulation according to A. D, E Phagocytosis and intracellular killing of C. perfringens was detected by CFU enumeration. Data are shown as mean ± SEM. Data were pooled from 3 independent experiments. Statistical significance is indicated by *p < 0.05, **p < 0.01, and ***p < 0.001.
STING deficiency impairs trained immunity and has a defect in bacterial clearance
Given the strong relationship between STING and trained immunity, our study aimed to investigate whether STING deficiency impairs trained immunity following HKCA training. WT and STING−/− mice were trained with two i.p. injections of HKCA, and then PM were collected and restimulated with LPS and C. perfringens (Figure 4A). As expected, we found that PM from WT mice training with HKCA exhibited a heightened production of IL-6 and TNF-α in response to LPS stimulation compared with the control group, whereas no significant differences were observed in STING−/− PM in comparison with before and after HKCA training (Figure 4B , C). In accordance with the above results, HKCA training did not impact bacterial phagocytic ability and clearance in STING−/− PM (Figure 4D, E). STING−/− PM presented decreased levels of IL-6 and TNF-α compared to WT PM after infection with C. perfringens, stimulated or not with HKCA (Additional files 1A and B). Together, STING plays a crucial role in inducing protective trained immunity.

STING deficiency dampens the trained response to LPS stimulation and C. perfringens infection. WT and STING−/− PM were trained with HKCA and then were restimulated with LPS or C. perfringens at an MOI of 5 for the indicated times. A Flow chart of in vitro trained immunity experimental procedure. B, C TNF-α and IL-6 production was measured in the supernatants of mouse PM response to LPS stimulation according to A. D, E Phagocytosis and intracellular killing of C. perfringens was detected by CFU enumeration. Data are shown as mean ± SEM. Data were pooled from 3 independent experiments. Statistical significance is indicated by *p < 0.05, **p < 0.01, and ***p < 0.001.
STING is dependent on the Akt/mTOR/HIF1α pathway in HKCA-induced trained immunity
These findings prompted us to further investigate the specific mechanism of the regulation of HKCA-induced trained immunity by STING. Since mTOR is a key regulator of β-glucan-induced trained immunity in monocytes, we sought to determine if mTOR signaling is also required for STING-mediated trained immunity in macrophages. Surprisingly, HKCA or DMXAA training both activated the mTOR and STING signaling pathways, as shown by the increased levels of p-AKT, p-mTOR, HIF1α, p-TBK1, and p-IRF3 compared to the control cells before secondary stimulation (Figure 5A–F). When mTOR inhibitor rapamycin was added, HKCA training-induced proinflammatory cytokine (TNF-α and IL-6) production was significantly decreased upon LPS stimulation compared with the control group (Figure 6A, B). Consistently, the addition of rapamycin reduced the bacterial phagocytic ability and clearance upon C. perfringens infection (Figure 6C, D). Consistent with the above results, we found that STING−/− PM displayed notably reduced levels of p-AKT, p-mTOR, and HIF1α after HKCA training compared with those in WT PM before secondary stimulation (Figure 6E–H). Collectively, these results indicate that STING deficiency leads to a defect in trained immunity by impairing the mTOR signaling pathway.

DMXAA or HKCA training activates mTOR signaling. WT PM were trained with DMXAA or HKCA for 24 h and rested for 5 days. A The cellular lysate was analyzed for p-mTOR, p-AKT, HIF1α, p-TBK1, p-IRF3, and β-actin by Western blotting. B–F The gray intensity value of proteins was calculated using ImageJ software. Data are shown as mean ± SEM. Data were pooled from 3 independent experiments. Statistical significance is indicated by *p < 0.05, **p < 0.01, and ***p < 0.001.

Akt/mTOR/HIF1α pathway is indispensable for STING-mediated trained immunity. PM were pretreated with rapamycin (10 μM) for 1 h, and then the cells were trained with HKCA for 24 h and rested for 5 days. PM were restimulated with LPS or C. perfringens at an MOI of 5 for the indicated times. A, B TNF-α and IL-6 production were measured in the supernatants of PM response to LPS stimulation. C, D Phagocytosis and intracellular killing of C. perfringens was detected by CFU enumeration. E The cellular lysate was analyzed for p-mTOR, p-AKT, HIF1α, and β-actin by Western blotting. F–H The gray intensity value of proteins was calculated using ImageJ software. Data are shown as mean ± SEM. Data were pooled from 3 independent experiments. Statistical significance is indicated by *p < 0.05, **p < 0.01, and ***p < 0.001.
STING-dependent trained immunity contributes to host protection against C. perfringens soft tissue infection
We further established a gas gangrene model to validate the role of HKCA-induced trained immunity during C. perfringens infection in vivo. WT and STING−/− mice were trained with two i.p. injections of HKCA on days -7 and -4. 7 days later, mice were intramuscularly infected with 1 × 107 CFU of C. perfringens for 24 h (Figure 7A). Mice developed gas gangrene in the control group, characterized by marked swelling and hemorrhage. The HKCA training group exhibited much milder symptoms in WT mice, but there was no significant difference in STING−/− mice before and after HKCA training (Figure 7B, C). In line with this, the HKCA training group harbored dramatically decreased loads of C. perfringens in the muscles of WT mice. No significant change in bacterial burden was observed in STING−/− mice before and after HKCA training (Figure 7D). These findings suggest that STING-dependent trained immunity enhances host resistance to C. perfringens soft tissue infection.

STING-dependent trained immunity protects mice from C. perfringens soft tissue infection. Age- and sex-matched WT and STING−/− mice were trained with two i.p. injections of HKCA on days −7 and −4. PBS was used as the control. 7 days later, mice were intramuscularly infected with 2 × 107 CFU of C. perfringens for 24 h. A Flow chart of in vivo trained immunity experimental procedure. B Representative gross images of legs. C Cumulative gross pathology scores. D Bacterial counts. Data are shown as mean ± SEM. Data were pooled from 3 independent experiments. Statistical significance is determined by *p < 0.05 and **p < 0.01.
Discussion
C. perfringens is an opportunistic pathogen that ubiquitously spreads in the abiotic environment and biotic intestinal tracts. Due to its diverse virulence factors and strong resistance to adversity, C. perfringens causes a wide variety of diseases including intestinal or foodborne diseases as well as gangrenes, which harms both human health and the development of agriculture. A crucial first step in tackling this problem is the detailed understanding of the host's innate immune response against C. perfringens infection based on the relative paucity of research. At present, only a few studies have demonstrated that toll-like receptor 4 accelerates granulopoiesis and enhances host defense against C. perfringens infection [21]. The toxins lecithinase and perfringolysin O of C. perfringens have been found to activate the NLRP3 inflammasome, and NLRP3 deficiency leads to increased host susceptibility to C. perfringens infection [22, 23]. Consequently, the need for new preventive and therapeutic strategies has become more urgent.
Trained immunity offers interesting perspectives for dual memory vaccine designing and treatment strategies in infectious diseases. In our study, there is a novel finding that STING-dependent trained immunity protected against C. perfringens infection by regulating mTOR signaling. Using a trained immunity model in PM, we first demonstrate that HKCA training induced significant elevations of TNF-α and IL-6 after LPS restimulation. This was consistent with the “classic” trained immunity model in human monocytes/macrophages [24]. Notably, HKCA-trained PM exhibited decreased levels of cytokines but enhanced bacterial phagocytic ability and clearance response to C. perfringens infection. We speculate that this may be due to the intertwined interactions between inflammation and infection, both of which are mutually the cause or consequence. As has been reported previously, trained immunity confers broad-spectrum protection against bacterial infections, whereas the trend of cytokine secretion is inconsistent with each other [25]. These results demonstrate that HKCA-induced trained immunity enhances macrophage function and promotes host defense against C. perfringens infection.
As an important component of the innate immune system, STING functions as an adaptor molecule for intracellular DNA sensors (such as cGAS, IFI16, and DNA-PK) in addition to as a direct sensor of cyclic dinucleotides [26,27,28,29]. Traditionally, STING activation induces type I interferon responses which have been associated with inflammation, infection, cancer, and autoimmune diseases. However, the biological implications of STING in the process of trained immunity have not been characterized. In this study, we found that STING signaling was upregulated in all HKCA-trained samples before and after stimulation with LPS or C. perfringens. To further explore the underlying causes, we trained PM with the STING agonist DMXAA. Intriguingly, our results show that DMXAA was a strong inducer of trained immunity, characterized by increased responsiveness to LPS challenge and enhanced phagocytosis and bacterial killing ability to C. perfringens infection. Importantly, using STING−/− mice, we further show that STING deficiency impairs trained immunity and has a defect in bacterial clearance. These results align with previous research demonstrating that a recombinant BCG expressing a STING agonist shows enhanced antitumor efficacy by triggering trained immunity remodeling [30]. Taken together, STING acts as a key regulator and maintainer of trained immunity.
Epigenetic and metabolic reprogramming underlies the induction of trained immunity. mTOR functions as a master metabolic regulator in the complex networks of reprogramming. PI3K signaling is a key in the mTOR upstream activation process, which has long been known as an essential mediator of trained immunity [31]. Akt/mTOR/HIF1α-dependent induction of aerobic glycolysis has been found to represent the metabolic basis of trained immunity [1]. In our study, enhanced mTOR signaling activation corresponded to increased expression levels of STING signaling pathway-related proteins after HKCA or DMXAA training. More importantly, inhibiting the mTOR signaling with rapamycin attenuated trained-PM functions, mainly characterized by decreased inflammatory responses after LPS stimulation and reduced phagocytosis and the bacterial killing ability upon C. perfringens challenge. Similarly, mTOR-dependent ROS production regulates the oxLDL-induced trained immunity in human monocytes [32]. A defect in HIF-1α recruitment of neonatal immune cells under low oxygen partial pressure leads to decreased phagocytosis and ROS production [33]. The application of rapamycin effectively inhibits the proinflammatory memory-like response of microglia induced by B cell-activating factor [34]. To further clarify the role of mTOR signaling in the process of STING-dependent trained immunity, we detected the expression of p-mTOR, p-AKT, and HIF1α. Compared with WT cells, STING−/− PM exhibit decreased levels of the above proteins after HKCA training, approaching the levels seen in the non-training group. Hence, our investigation highlighted the significance of STING in promoting trained immunity induction for host defense against C. perfringens infection via the mTOR signaling pathway. Hopefully, these studies will provide candidate resources for the development of STING-based vaccine adjuvants, the design of dual memory vaccines which induce a strong persistent memory response, and preventive therapeutics against infectious disease.
Availability of data and materials
All data generated or analyzed during this study are included in this published article.
Abbreviations
- C. perfringens :
-
Clostridium perfringens
- HKCA:
-
heat-killed Candida albicans
- STING:
-
stimulator of interferon genes
- WT:
-
wild-type
- PRR:
-
pattern-recognition receptors
- IFN:
-
type I interferons
- PM:
-
peritoneal macrophages
- PBS:
-
phosphate-buffered saline
- i.p.:
-
intraperitoneally
- CFU:
-
colony-forming units
References
Cheng SC, Quintin J, Cramer RA, Shepardson KM, Saeed S, Kumar V, Giamarellos-Bourboulis EJ, Martens JH, Rao NA, Aghajanirefah A, Manjeri GR, Li Y, Ifrim DC, Arts RJ, van der Veer BM, Deen PM, Logie C, O’Neill LA, Willems P, van de Veerdonk FL, van der Meer JW, Ng A, Joosten LA, Wijmenga C, Stunnenberg HG, Xavier RJ, Netea MG (2014) mTOR- and HIF-1α-mediated aerobic glycolysis as metabolic basis for trained immunity. Science 345:1250684
European Food Safety Authority and European Centre for Disease Prevention and Control (EFSA and ECDC) (2018) The European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2017. EFSA J 16:e05500
Netea MG, Joosten LA, Latz E, Mills KH, Natoli G, Stunnenberg HG, O’Neill LA, Xavier RJ (2016) Trained immunity: a program of innate immune memory in health and disease. Science 352:aaf1098
Hu Z, Lu SH, Lowrie DB, Fan XY (2022) Trained immunity: a Yin-Yang balance. MedComm 3:e121
Shao Y, Saredy J, Xu K, Sun Y, Saaoud F, Drummer Ct LuY, Luo JJ, Lopez-Pastrana J, Choi ET, Jiang X, Wang H, Yang X (2021) Endothelial immunity trained by coronavirus infections, DAMP stimulations and regulated by anti-oxidant NRF2 may contribute to inflammations, myelopoiesis, COVID-19 cytokine storms and thromboembolism. Front Immunol 12:653110
Christ A, Günther P, Lauterbach MAR, Duewell P, Biswas D, Pelka K, Scholz CJ, Oosting M, Haendler K, Baßler K, Klee K, Schulte-Schrepping J, Ulas T, Moorlag S, Kumar V, Park MH, Joosten LAB, Groh LA, Riksen NP, Espevik T, Schlitzer A, Li Y, Fitzgerald ML, Netea MG, Schultze JL, Latz E (2018) Western diet triggers NLRP3-dependent innate immune reprogramming. Cell 172:162–175.e14
O’Neill LAJ, Netea MG (2020) BCG-induced trained immunity: can it offer protection against COVID-19? Nat Rev Immunol 20:335–337
Schnack L, Sohrabi Y, Lagache SMM, Kahles F, Bruemmer D, Waltenberger J, Findeisen HM (2019) Mechanisms of trained innate immunity in oxLDL primed human coronary smooth muscle cells. Front Immunol 10:13
Lajqi T, Frommhold D, Braun M, Alexander Kranig S, Pöschl J, Hudalla H (2022) Gram-positive Staphylococcus aureus LTA promotes distinct memory-like effects in murine bone marrow neutrophils. Cell Immunol 376:104535
Lajqi T, Braun M, Kranig SA, Frommhold D, Pöschl J, Hudalla H (2021) LPS induces opposing memory-like inflammatory responses in mouse bone marrow neutrophils. Int J Mol Sci 22:9803
Lajqi T, Marx C, Hudalla H, Haas F, Große S, Wang ZQ, Heller R, Bauer M, Wetzker R, Bauer R (2021) The role of the pathogen dose and PI3Kγ in immunometabolic reprogramming of microglia for innate immune memory. Int J Mol Sci 22:2578
Lajqi T, Köstlin-Gille N, Bauer R, Zarogiannis SG, Lajqi E, Ajeti V, Dietz S, Kranig SA, Rühle J, Demaj A, Hebel J, Bartosova M, Frommhold D, Hudalla H, Gille C (2023) Training vs. tolerance: the Yin/Yang of the innate immune system. Biomedicines 11:766
Zhou J, Lv J, Carlson C, Liu H, Wang H, Xu T, Wu F, Song C, Wang X, Wang T, Qian Z (2021) Trained immunity contributes to the prevention of Mycobacterium tuberculosis infection, a novel role of autophagy. Emerg Microbes Infect 10:578–588
Dos Santos JC, Barroso de Figueiredo AM, Teodoro Silva MV, Cirovic B, de Bree LCJ, Damen M, Moorlag S, Gomes RS, Helsen MM, Oosting M, Keating ST, Schlitzer A, Netea MG, Ribeiro-Dias F, Joosten LAB (2019) β-glucan-induced trained immunity protects against Leishmania braziliensis infection: a crucial role for IL-32. Cell Rep 28:2659–2672.e6
Brandi P, Conejero L, Cueto FJ, Martínez-Cano S, Dunphy G, Gómez MJ, Relaño C, Saz-Leal P, Enamorado M, Quintas A, Dopazo A, Amores-Iniesta J, Del Fresno C, Nistal-Villán E, Ardavín C, Nieto A, Casanovas M, Subiza JL, Sancho D (2022) Trained immunity induction by the inactivated mucosal vaccine MV130 protects against experimental viral respiratory infections. Cell Rep 38:110184
Owen AM, Luan L, Burelbach KR, McBride MA, Stothers CL, Boykin OA, Sivanesam K, Schaedel JF, Patil TK, Wang J, Hernandez A, Patil NK, Sherwood ER, Bohannon JK (2022) MyD88-dependent signaling drives toll-like receptor-induced trained immunity in macrophages. Front Immunol 13:1044662
Quintin J, Saeed S, Martens JHA, Giamarellos-Bourboulis EJ, Ifrim DC, Logie C, Jacobs L, Jansen T, Kullberg BJ, Wijmenga C, Joosten LAB, Xavier RJ, van der Meer JWM, Stunnenberg HG, Netea MG (2012) Candida albicans infection affords protection against reinfection via functional reprogramming of monocytes. Cell Host Microbe 12:223–232
Kleinnijenhuis J, Quintin J, Preijers F, Joosten LA, Ifrim DC, Saeed S, Jacobs C, van Loenhout J, de Jong D, Stunnenberg HG, Xavier RJ, van der Meer JW, van Crevel R, Netea MG (2012) Bacille Calmette-Guerin induces NOD2-dependent nonspecific protection from reinfection via epigenetic reprogramming of monocytes. Proc Natl Acad Sci U S A 109:17537–17542
Zhang Z, Zhou H, Ouyang X, Dong Y, Sarapultsev A, Luo S, Hu D (2022) Multifaceted functions of STING in human health and disease: from molecular mechanism to targeted strategy. Signal Transduct Target Ther 7:394
Awad MM, Ellemor DM, Boyd RL, Emmins JJ, Rood JI (2001) Synergistic effects of alpha-toxin and perfringolysin O in Clostridium perfringens-mediated gas gangrene. Infect Immun 69:7904–7910
Takehara M, Kobayashi K, Nagahama M (2021) Toll-like receptor 4 protects against Clostridium perfringens infection in mice. Front Cell Infect Microbiol 11:633440
Liu Y, Lei YX, Li JW, Ma YZ, Wang XY, Meng FH, Wu YJ, Wang N, Liang J, Zhao CQ, Yang Y, Chen GX, Yu SX (2023) G protein-coupled receptor 120 mediates host defense against Clostridium perfringens infection through regulating NOD-like receptor family pyrin domain-containing 3 inflammasome activation. J Agric Food Chem 71:7119–7130
Mathur A, Kay C, Xue Y, Pandey A, Lee J, Jing W, Enosi Tuipulotu D, Lo Pilato J, Feng S, Ngo C, Zhao A, Shen C, Rug M, Miosge LA, Atmosukarto II, Price JD, Ali SA, Gardiner EE, Robertson AA, Awad MM, Lyras D, Kaakoush NO, Man SM (2023) Clostridium perfringens virulence factors are nonredundant activators of the NLRP3 inflammasome. EMBO Rep 24:e54600
Zhang B, Moorlag SJ, Dominguez-Andres J, Bulut Ö, Kilic G, Liu Z, van Crevel R, Xu CJ, Joosten LA, Netea MG, Li Y (2022) Single-cell RNA sequencing reveals induction of distinct trained-immunity programs in human monocytes. J Clin Invest 132:e147719
Ciarlo E, Heinonen T, Théroude C, Asgari F, Le Roy D, Netea MG, Roger T (2020) Trained immunity confers broad-spectrum protection against bacterial infections. J Infect Dis 222:1869–1881
Sun L, Wu J, Du F, Chen X, Chen ZJ (2013) Cyclic GMP-AMP synthase is a cytosolic DNA sensor that activates the type I interferon pathway. Science 339:786–791
Unterholzner L, Keating SE, Baran M, Horan KA, Jensen SB, Sharma S, Sirois CM, Jin T, Latz E, Xiao TS, Fitzgerald KA, Paludan SR, Bowie AG (2010) IFI16 is an innate immune sensor for intracellular DNA. Nat Immunol 11:997–1004
Burdette DL, Monroe KM, Sotelo-Troha K, Iwig JS, Eckert B, Hyodo M, Hayakawa Y, Vance RE (2011) STING is a direct innate immune sensor of cyclic di-GMP. Nature 478:515–518
Ferguson BJ, Mansur DS, Peters NE, Ren H, Smith GL (2012) DNA-PK is a DNA sensor for IRF-3-dependent innate immunity. Elife 1:e00047
Singh AK, Praharaj M, Lombardo KA, Yoshida T, Matoso A, Baras AS, Zhao L, Srikrishna G, Huang J, Prasad P, Powell JD, Kates M, McConkey D, Pardoll DM, Bishai WR, Bivalacqua TJ (2022) Re-engineered BCG overexpressing cyclic di-AMP augments trained immunity and exhibits improved efficacy against bladder cancer. Nat Commun 13:878
Lajqi T, Lang GP, Haas F, Williams DL, Hudalla H, Bauer M, Groth M, Wetzker R, Bauer R (2019) Memory-like inflammatory responses of microglia to rising doses of LPS: key role of PI3Kγ. Front Immunol 10:2492
Sohrabi Y, Lagache SMM, Schnack L, Godfrey R, Kahles F, Bruemmer D, Waltenberger J, Findeisen HM (2018) mTOR-dependent oxidative stress regulates oxLDL-induced trained innate immunity in human monocytes. Front Immunol 9:3155
Schlegel C, Liu K, Spring B, Dietz S, Poets CF, Hudalla H, Lajqi T, Köstlin-Gille N, Gille C (2023) Decreased expression of hypoxia-inducible factor 1α (HIF-1α) in cord blood monocytes under anoxia. Pediatr Res 93:870–877
Wang J, Yang C, Hou X, Xu J, Yun Y, Qin L, Yang P (2021) Rapamycin modulates the proinflammatory memory-like response of microglia induced by BAFF. Front Immunol 12:639049
Acknowledgements
We would like to acknowledge Tao Wang, Xu Zhang, Heng Yang, and Meng-Ge Chen, the College of Veterinary Medicine, Jilin University, for their helpful advice and technical assistance with our experiments. Our thanks are also extended to express our gratitude to all the people who made this work.
Funding
This work was supported by National Natural Science Foundation of China (No. 32202822, No. 32070119, and No. 31972682).
Author information
Authors and Affiliations
Contributions
Z-ZL, Y-JY, and WC designed experiments. Z-ZL, C-KZ, X-QL, YG, X-YL, J-BZ, QY, LZ, J-GZ, and XA performed the experiments and analyzed the data. Z-ZL wrote the manuscript. Y-JY and WC revised the manuscript. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
All animal studies were conducted according to experimental practices and standards approved by the Animal Welfare and Research Ethics Committee at Jilin University (No. KT202202182).
Competing interests
The authors declare that they have no competing interests.
Additional information
Handling editor: Tina Dalgaard
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1:
The secretion of TNF-α and IL-6 in WT and STING-/- PM after C. perfringens infection. WT and STING-/- PM were trained with HKCA and then were restimulated with C. perfringens at an MOI of 5 for the indicated times. A, B TNF-α and IL-6 production was measured in the supernatants of mouse PM. Data are shown as mean ± SEM. Data were pooled from 3 independent experiments. Statistical significance is indicated by *p < 0.05 and ***p < 0.001.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Liu, ZZ., Zhou, CK., Lin, XQ. et al. STING-dependent trained immunity contributes to host defense against Clostridium perfringens infection via mTOR signaling. Vet Res 55, 52 (2024). https://doi.org/10.1186/s13567-024-01301-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13567-024-01301-1