
Abstract
Fowl typhoid (FT), a septicemic disease caused by Salmonella Gallinarum (SG), and infectious bronchitis (IB) are two economically important avian diseases that affect poultry industry worldwide. Herein, we exploited a live attenuated SG mutant, JOL967, to deliver spike (S) protein 1 of IB virus (V) to elicit protective immunity against both FT and IB in chickens. The codon optimized S1 nucleotide sequence was cloned in-frame into a prokaryotic constitutive expression vector, pJHL65. Subsequently, empty pJHL65 or recombinant pJHL65-S1 plasmid was electroporated into JOL967 and the resultant clones were designated as JOL2068 and JOL2077, respectively. Our results demonstrated that the chickens vaccinated once orally with JOL2077 elicited significantly (p < 0.05) higher IBV-specific humoral and cell-mediated immunity compared to JOL2068 and PBS control groups. Consequently, on challenge with the virulent IBV strain at 28th day post-vaccination, JOL2077 vaccinated birds displayed significantly (p < 0.05) lower inflammatory lesions in virus-targeted tissues compared to control groups. Furthermore, 33.3% (2 of 6) of birds vaccinated with JOL2077 vaccine had shown virus recovery from tracheal tissues compared to 100% (6 of 6) recovery obtained in both the control groups. Against wild-type SG lethal challenge, both JOL2077 and JOL2068 vaccinated groups exhibited only 10% mortality compared to 80% mortality observed in PBS control group. In conclusion, we show that JOL2077 can induce efficient IBV- and carrier-specific protective immunity and can act as a bivalent vaccine against FT and IB. Further studies are warranted to investigate the potential of JOL2077 vaccine in broiler and young layer birds.
Similar content being viewed by others

Introduction
Infectious bronchitis virus (IBV), one of the prototype viruses of the Coronaviridae family, is an enveloped, single-stranded, positive-sense RNA virus that causes significant economic losses in poultry industry worldwide [1, 2]. In chickens, IBV causes an acute highly contagious disease which affects the respiratory tract, gut, kidneys, bursa and reproductive systems, and the disease is characterized by 50–100% morbidity and 0–25% mortality rates in affected flocks [3, 4]. The viral genome encodes four major structural proteins, namely membrane protein (M), nucleocapsid protein (N), spike protein (S) and envelope protein (E), that facilitate host entry and virus assembly [5]. The S protein is post-translationally cleaved by host cell serine proteases to form two subunits, S1 and S2, and this cleavage plays an important role in viral infectivity [6]. The S1 subunit binds to host cell receptors and is the main target for induction of serotype-specific neutralizing and hemagglutination-inhibiting antibodies [7, 8], and it is sufficient to induce protective immune responses [9]. To prevent economic losses resulted by IB, live attenuated and inactivated oil adjuvanted IBV vaccines have been employed in the field [9]; however, these types of vaccines are associated with some limitations. Inactivated IBV vaccines are safe, but relatively costlier and less effective than live attenuated vaccines, while use of live attenuated vaccines may result in the emergence of novel variants of the virus [10, 11]. Furthermore, the existence of more than 20 IBV serotypes with little cross protective immunity among vaccine strains results in poor efficacy of the currently available vaccines. The introduction and development of an effective and safe vaccine against each IBV serotype is not an economically viable option for the poultry industry. Therefore, novel approaches should be devised that are not only egg independent and cost-effective, but easy to amplify and can provide efficient protection against the circulating IBV strains.
Identification and characterization of regional and most circulating serotypes plays an important role in the development of a successful vaccination program. Over the past decade, nephropathogenic IBV variants have been continuously evolved in South Korea via accumulated point mutations and by recombination with other existing strains [12]. Thus, emphasizing the need for the development of efficient and effective vaccination strategies which respond to the circulating field strains in a very short notice. In accordance with this notion, the present study exploited Salmonella Gallinarum (SG) to deliver immunogenic S1 protein (20–263 amino acid residues) of most emerging and circulating nephropathogenic strain of IBV in South Korea. SG is an intracellular Gram-negative bacterium that causes fowl typhoid (FT) in domestic birds, primarily chickens, and rarely causes food poisoning in humans [13]. In chickens, FT is characterized by severe inflammation and acute mortality that causes significant economic losses in poultry industry worldwide [13,14,15]. The disease has worldwide distribution and is endemic in many parts of the world. Besides farm hygiene and biosecurity measures, the control and eradication of FT can be effectively achieved by vaccination with live attenuated and inactivated killed vaccines. We previously have developed an attenuated SG vaccine strain (JOL916) that effectively controls FT infection in chickens [16]. Exploiting live attenuated Salmonella system to deliver IBV S1 immunogenic protein is a highly economical vaccination strategy that allows for a quick response to deployment of vaccines, as it circumvents the need for a constant supply of specific-pathogen-free (SPF) embryonated eggs. Furthermore, Salmonella-based vaccination strategy induces efficient antigen-specific humoral and cell-mediated immunity [17,18,19], and these vaccines have already been approved for use in poultry [20]. Herein, we show that attenuated SG mutant delivering immunogenic S1 protein elicited efficient IBV-specific humoral and cell-mediated immune (CMI) responses and conferred significant protection against a virulent IBV challenge. We also demonstrate that attenuated SG delivering S1 protein protected chickens against a lethal wild-type SG challenge.
Materials and methods
Bacterial strains, plasmids and virus
The bacterial strains and plasmids used in this study are listed in Table 1. All the bacterial strains were grown in Luria–Bertani (LB) broth at 37 °C. IBV strain was purchased from Korea Veterinary Culture Collection (KVCC-VR1100015; South Korea) and propagated in the allantoic cavities of 9- to 10-day-old SPF embryonated chicken eggs. The 50% egg infective dose (EID)50 was calculated as described previously [21].
Construction of an attenuated SG mutant delivering IBV spike protein 1
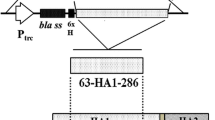
The partial immunogenic S1 amino acid sequence20–263 of most emerging nephropathogenic IBV Korean strain (Accession No. ADQ01077.1) was chosen, in this study, based on the presence of major neutralizing B and T cell epitopes that are well-known to elicit complete protection against IBV infection in vaccinated chickens [22,23,24]. The S1 nucleotide sequence was codon optimized for efficient expression in SG. The optimized gene sequence was chemically synthesized (Bionee, Korea) and then built into the pJHL65 plasmid, an asd+ constitutive expression vector, and propagated in an asd mutated Escherichia coli strain as described previously [25]. The S1 gene sequence was cloned in frame downstream to the beta-lactamase signal sequence (bla SS) of pJHL65 vector, so that the intended protein would get secreted out of the bacteria [26]. The recombinant plasmid, pJHL65-S1, was subsequently transformed into an attenuated auxotrophic mutant of SG strain, JOL967, and the resultant clone was designated as JOL2077. The JOL967 strain was constructed by the deletion of the lon, cpxR, and asd genes from the wild-type SG strain, JOL394 isolate, using allelic exchange method as described and reported elsewhere [16], and used as the delivery vehicle for the S1 protein. To produce the coating antigen for determination of IBV-specific antibody responses, the S1 gene was cloned into pET28a (+) expression vector (Novagen, San Diego, USA) that was subsequently transformed into E. coli BL21 (DE3) pLysS strain (Novagen, USA) for protein expression. The expressed S1 protein in both S. typhimurium and E. coli was confirmed by Western blot analysis using polyclonal infectious bronchitis virus antibody (#ab31671, Abcam). The E. coli expressed S1 protein was purified by Ni–NTA affinity column chromatography and dialysed against PBS (three washes). Purified protein was quantified by a Bradford assay [27], filtered, and stored at −20 °C until further use.
Immunization and challenge studies
All animal experimentation work was approved by the Chonbuk National University Animal Ethics Committee (CBNU2015-00085) and the chicken experiment was carried out according to the guidelines of the Korean Council on Animal Care. One-day-old female layer chickens (Corporation of Join hatchery, Republic of Korea) were maintained under standard conditions and provided antibiotic-free food and water ad libitum. Four weeks later, the chickens were randomly divided into three groups (n = 15) and vaccinated once orally with sterile phosphate buffered saline (PBS, 100 µL in each bird), JOL2068 (SG carrying an empty pJHL65 vector) or JOL2077 (SG-pJHL65-S1). The 109 colony forming units (CFU) of JOL2068 or JOL2077 were used for inoculation in each bird. Blood was drawn from jugular vein of six randomly vaccinated chickens on day 28 post-vaccination and serum samples were subsequently isolated for IgG analysis. To assess mucosal secretory (s) IgA responses, five chickens on day 28 post-vaccination were sacrificed from each group and intestinal wash samples were collected in PBS (pH 7.2) containing antibiotics and protease inhibitor cocktail (Sigma Aldrich).
Four weeks post-vaccination, all the vaccinated chickens were intranasally challenged with a virulent dose of IBV (104 EID50) and 6 days later post-challenge, the tracheal tissues (n = 6) were aseptically processed and inoculated into 15–16 day old chicken embryos to isolate the challenged virus as described previously [21]. The clinical protection against IBV was assessed based on the death and/or stunting of embryos. For histopathological analysis of trachea, lungs, kidneys and bursa, each tissue from sacrificed birds (n = 6) was aseptically collected on day 6 post-challenge and fixed with 10% formalin, embedded in paraffin and cut into 5 μm thin sections, which were subsequently processed as previously described [18]. The hematoxylin and eosin (H&E) stained sections were examined by light microscopy (Axio Imager 2, Zeiss, Germany) and digital imaging software (Axio Vision, Zeiss, Germany). We also separately maintained three groups (n = 10) vaccinated once orally with JOL2068, JOL2077 or sterile PBS, and 4 weeks later, all the birds were challenged orally with 100 µL of a suspension containing 1.6 × 106 CFU of a wild-type SG (JOL394 strain) as described previously [28]. The protective efficacy of the SG-S1 based vaccine (JOL2077) against JOL394 was evaluated in the context of the mortality of the challenged birds, which was monitored daily for 12 consecutive days. At the end of the observation period post-challenge, the surviving birds were sacrificed to determine the bacterial load in liver and spleen as previously described [28].
Systemic IgG and mucosal IgA antigen-specific humoral responses
The systemic IBV and SG-ompA specific IgG and mucosal IgA responses were analysed in vaccinated sera and in intestinal wash samples, respectively, by an indirect ELISA [28]. Purified S1 protein and ompA antigen (each 250 ng/well) were used as coating antigens in an indirect ELISA to determine the IBV and the SG specific humoral responses, respectively.
IBV-specific cellular immune responses
The IBV-specific cell-mediated immunity elicited by vaccination was evaluated by a lymphocyte proliferation test. Four weeks post-vaccination, in vitro proliferative capacity of vaccinated peripheral blood mononuclear cells (PBMCs) (1 × 106, n = 6) in response to the recall S1 protein was determined by a MTT [3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide] based assay [18]. The PBMCs were isolated using Histopaque density gradient as previously described [18]. The proliferative capacity of vaccinated PBMCs in response to recall S1 antigen was further analyzed by flow cytometry. To this end, PBMCs (1 × 106, n = 6) isolated from vaccinated birds on day 28 post-vaccination were stimulated with S1 antigen (10 µg/mL) for 48 h and then S1-specific CD4+ and CD8+ T cell responses were analysed by flow cytometry as previously described [28].
For analysis of Th1 (IFN-γ) and Th2 (IL-4 and IL-10) cytokine responses, PBMCs (1 × 106, n = 6) were isolated from vaccinated and control birds and restimulated with S1 protein (10 µg/mL) in vitro for 24 h incubation at 37 °C in 5% CO2. Then total RNA isolated from stimulated cells was analysed for IFN-γ, IL-4 and IL-10 mRNA transcription levels by qRT-PCR assay as previously described [28].
Statistical analysis
All the obtained data was analysed using GraphPad prism 6.00 program (San Diego, CA, USA). Statistical significance was determined by one-way ANOVA (with Tukey’s multiple comparisons tests). A non-parametric Chi square test was used to analyze significant differences in mortality of birds following SG challenge. Data are represented as mean ± standard deviation. p values of < 0.05 were considered statistically significant.
Results
Attenuated SG mutant efficiently expressed and secreted IBV S1 protein
To construct the SG-based IB vaccine, we predicted highly antigenic epitopes of IBV S1 protein using BepiPred 1.0 Server program based on the protein characteristics such as structural domains, hydrophilicity residues, antigenicity and surface probability (Additional file 1) [29]. To direct the expressed protein to the periplasmic space, the S1 gene was cloned in frame downstream to the bla SS of the pJHL65 vector. The insertion of S1 into pJHL65 vector was confirmed by digestion of the positive clones with EcoR1 and HindIII to release a fragment of 738 bp. Subsequently, the pJHL65-S1 gene construct was electroporated into JOL967 strain and the resultant clone was designated as JOL2077. Western blot analysis revealed that JOL2077 strain efficiently expressed and secreted S1 protein in the culture supernatant of cultured JOL2077 bacteria (Additional file 2). The S1 protein was biologically active as evidenced by the reactivity with polyclonal IBV-specific antibody and by the induction of the antigen-specific immune responses in vaccinated chickens as observed in this study.
Orally administered JOL2077 vaccine induces efficient S1-specific systemic and mucosal antibody responses
To investigate the effect of vaccination on the systemic IgG and the mucosal sIgA antibody responses, we orally vaccinated chickens with PBS, JOL2068 or JOL2077, and 28 days later IgG in serum and sIgA in intestinal wash samples were measured by an indirect ELISA (Figure 1). Our results demonstrated that the chickens inoculated once orally with JOL2077 elicited significantly (p < 0.05) higher both IgG and sIgA responses compared to the chickens that received either PBS or JOL2068 strain (Figure 1). The S1-specific IgG (Figure 1A) and sIgA (Figure 1B) responses were 3.6 and 3 folds, respectively, higher than JOL2068 control group. Our data thus clearly indicate that attenuated SG delivering S1 protein is capable of inducing antigen-specific systemic and mucosal humoral immune responses.

IBV-specific systemic IgG and mucosal IgA responses. Chickens (N = 15) were vaccinated with sterile PBS, JOL2068 or JOL2077, and 28 days later serum and intestinal wash samples were analysed for S1-specific IgG and IgA responses, respectively, by an indirect ELISA. A Serum IgG. B Intestinal sIgA responses. Each data point represents mean ± SD of six chickens. *p < 0.05.
Orally administered JOL2077 vaccine elicits efficient IBV-specific cellular immunity
To assess the effect of vaccination on the IBV-specific CMI responses, we measured mRNA levels of IFN-γ, IL-4 and IL-10 cytokines in vaccinated PBMCs restimulated with S1 protein in vitro for 24 h (Figure 2). Our results demonstrated that the chickens vaccinated with JOL2077 vaccine induced significantly (p < 0.05) higher mRNA levels of both Th1-type (IFN-γ) and Th2-type (IL-4 and IL-10) cytokines compared to the JOL2068 and the PBS control groups (Figure 2). The IFN-γ, IL-4 and IL-10 cytokines were 3.5, 5 and 4 folds, respectively, higher in JOL2077 vaccinated chickens compared to the JOL2068 control chickens, which showed non-significant and comparable levels to that of the PBS control chickens. Our data further revealed that mRNA levels of Th1 and Th2 cytokine levels were almost comparable, suggesting that vaccination with JOL2077 vaccine elicited mixed-type of immunity.

Analysis of cytokine gene expressions in PBMCs after stimulation with S1 antigen. Chickens (N = 15) were vaccinated with sterile PBS, JOL2068 or JOL2077, and PBMCs from vaccinated chickens after 28 days post-vaccination were stimulated with purified S1 antigen for 24 h and then analysed for induction of IFN-γ, IL-4 and IL-10 mRNA transcription levels by qRT-PCR assay. Results are expressed as log10 fold induction in mRNA transcription levels of stimulated PBMCs from vaccinated chickens compared to the unstimulated naïve PBMCs. Gene expressions were normalized to GAPDH and mRNA levels of naive cells were used as the calibrator. Data presented are mean ± SD of six chickens per group. *p < 0.05.
Next, we measured T cell proliferative responses by a MTT-based assay after restimulation of vaccinated and control PBMCs with S1 protein in vitro for 72 h. The proliferative responses were significantly (p < 0.05) higher in chickens that received JOL2077 vaccine compared to the JOL2068 and PBS treated groups (Figure 3A). The lymphoproliferative responses were consistent with the results of CD4+ and CD8+ T cell responses measured by flow cytometry after restimulation of vaccinated PBMCs with S1 protein in vitro for 48 h (Additional file 3). Our results indicated that JOL2077 vaccinated chickens elicited significantly (p < 0.05) higher IBV-specific CD4+ T cell responses compared to the JOL2068 and the PBS control groups (Figure 3B). Although CD8+ T cell responses were higher in JOL2077 vaccinated chickens, but the responses were non-significant compared to the PBS and the JOL2068 control groups (Figure 3B). Thus, our results clearly demonstrate that oral inoculation of SG delivering S1 protein is capable of eliciting both IBV-specific humoral and CMI responses.

IBV-specific cellular immune responses. A In vitro proliferations of lymphocytes from vaccinated chickens in response to the recall S1 antigen. Results are expressed as stimulation indices, defined as proliferation in response to recall antigen relative to the mock stimulated cells. Each data points represent mean ± SD of six chickens per group. B The CD3+CD4+ and CD3+CD8+ T-cell subsets in vaccinated and control chickens are expressed as a percentage of the gated cells. The data are presented as the mean ± S.D of six chickens. *p < 0.05.
Orally administered JOL2077 provides protection against virulent IBV challenge
To evaluate whether SG-based S1 vaccine could elicit immunoprotection against IB, we intranasally challenged all the vaccinated and the control chickens with a virulent dose of 104 EID50 IBV at 4 weeks post-vaccination. No mortality was observed in any of the chicken groups, but chickens in control groups showed depression, huddling and anorexia, and even diarrhea was observed in some birds. In contrast, JOL2077 vaccinated birds appeared normal and free from any disease symptoms. The protective efficacy of the JOL2077 vaccine was further analyzed by isolating the virus from tracheal tissues on day 6 post-challenge. The virus was recovered from 33.3% (2/6) of birds vaccinated with JOL2077 vaccine compared to the 100% (6/6) recovery obtained in both JOL2068 and PBS control groups (Table 2). These results clearly indicate that vaccination with JOL2077 vaccine had efficiently contained IBV replication inside the host tissues and thus protection from the clinical disease.
To further investigate the effect of JOL2077 vaccination on the virus-specific immune protection, histopathological studies were performed on tracheal, lung, kidney and bursal tissues collected from birds on day 6 post-IBV challenge (Figure 4). As expected, no lesions were found in any of the organs collected from the uninfected chickens. In contrast, chickens treated with either PBS or JOL2068 showed significant inflammatory lesions in all of the collected tissues. The trachea of both PBS and JOL2068 chicken groups showed extensive dropout, deciliation, epithelial and glandular desquamation, and infiltration of inflammatory cells (Figure 4A). In lungs, the most prominent histopathological changes observed in PBS and JOL2068 groups were congestion, hemorrhages, and infiltration of lymphocytes into the submucosa of secondary bronchi (Figure 4B). In kidneys, hemorrhages, multifocal necrosis of the renal tubules, and infiltration of mononuclear leukocytes were observed in PBS and JOL2068 control chickens (Figure 4C). The most prominent histopathological changes were observed in bursa of PBS and JOL2068 control chickens. The bursa showed atrophy of lymphoid follicles and widening of the interstitium (Figure 4D). In contrast to JOL2068 and PBS groups, chickens vaccinated with JOL2077 showed insignificant inflammation and the tissues appeared histologically similar to that of normal tissues. Thus, our data clearly indicate that JOL2077 vaccination had offered significant protection against the IBV challenge.

Photomicrographs of hematoxylin-and eosin-stained lung sections of chickens on 6th day post-IBV challenge. Chickens (N = 15) were immunized with sterile PBS, JOL2068 or JOL2077, and 28 days later all the vaccinated chickens were challenged with 104 EID50 IBV. At 6th day post-challenge, chickens (n = 10) were sacrificed and trachea, lung, kidneys and bursal tissues were collected for histological analysis. Black arrows represent inflammatory lesions.
JOL2077 vaccine induces SG-specific protective immune responses
To investigate whether JOL2077 vaccine delivering S1 protein can induce SG-specific protective immune responses, three groups of chickens were separately maintained and vaccinated with PBS, JOL2068 or JOL2077, and 4 weeks later SG-specific IgG and sIgA responses were measured in serum and intestinal wash samples, respectively (Figure 5), Our results demonstrated that chickens vaccinated with either JOL2068 or JOL2077 elicited significantly (p < 0.05) higher both systemic IgG and sIgA responses compared to the PBS control group. Both JOL2077 and JOL2068 vaccination groups showed comparable levels of antibody responses, suggesting that SG can be exploited to deliver foreign antigens without affecting the SG-specific immunity.

JOL2077 vaccine induced efficient SG-specific systemic IgG and mucosal IgA responses. Chickens (N = 10) were vaccinated with sterile PBS, JOL2068 or JOL2077, and serum and intestinal wash samples were analysed for ompA SG-specific IgG and sIgA levels, respectively, by an indirect ELISA. A Serum IgG. B Intestinal sIgA responses. Each data point represents mean ± SD of six chickens. *p < 0.05. ns: non-significant.
Next, we investigated the protective efficacy of JOL2077 vaccine against FT by orally challenging vaccinated and control chickens with a virulent dose (1.6 × 106 CFU) of wild-type SG bacteria. The mortality of birds, index of protection efficacy in this study, was observed for 12 days post-challenge (Table 3). The chickens vaccinated with either JOL2068 or JOL2077 exhibited lower mortality rate (10%, 1 of 10 birds died) compared to the control chickens, which showed 80% mortality (8 of 10 birds died). The efficacy of JOL2077 vaccine was further evaluated by post-mortem analysis of the survived birds at 12 days post-challenge. Our results showed that the birds vaccinated with either JOL2068 or JOL2077 vaccine had significantly (p < 0.05) lower bacterial load in spleen and liver compared to the control group (Table 3).
Discussion
This study was aimed to investigate whether attenuated SG delivering S1 protein could induce protective immune responses against both IB and FT in chickens. Our strategy of employing Salmonella system as a delivery system for S1 protein has certain advantages over live attenuated or inactivated IB vaccines. These types of vaccines are easy to prepare, stable, convenient, economical, and easy to administer in chickens. Moreover, these types of vaccines does not require exogenous adjuvants as Salmonella bacteria provide an appropriate danger signals to the immune system, acting as natural adjuvant, thereby promoting efficient maturation and activation of dendritic cells (DCs) [25, 30], which is prerequisite for the induction of potent adaptive immune responses. Also SG-based IB vaccination can differentiate vaccinated birds from infected ones as only structural and immunogenic antigens are exploited for the vaccine development. Previously, Salmonella-based system has been exploited to deliver DNA encoded foreign antigens to induce protective immune responses against a variety of pathogens, especially those invading via the mucosal route [19, 31, 32]. However, a potential limitation associated with the Salmonella delivered DNA vaccines is the inability to stably maintain the plasmid DNA inside the bacterium. Moreover, the gene expressions of heterologous antigens encoded by DNA is inefficient, resulting in poor immunogenicity of the vaccine [31], thus, requiring multiple immunizations to elicit some level of significant protection in vaccinated animals [21]. In accordance with this notion, we hypothesized that SG delivering S1 antigen in protein format could induce efficient IBV-specific protective immunity in chickens. We show that single oral inoculation of SG-based S1 vaccine induced efficient protective immune responses against IBV in chickens. We also show that SG delivering S1 protein does not affect the induction of SG-specific humoral and cell-mediated protective immune responses.
The present study demonstrated that orally administered JOL2077 vaccine elicited efficient circulating IgG responses in vaccinated chickens. The role of circulating antibody titers in conferring protection against IBV remains controversial. Some studies have demonstrated that circulating antibodies did not correlate well with the protection from IBV infection [33, 34], while others have shown the importance of humoral immunity in disease recovery and virus clearance [35, 36]. Nevertheless, JOL2077 vaccination had conferred significant protection against IBV challenge and had efficiently contained IBV replication in virus-targeted tissues. Furthermore, JOL2077 vaccinated chickens displayed insignificant inflammatory lesions in IBV-targeted tissues. The role of IgA antibodies in providing resistance to IBV infection is well-documented [35, 37]. The elicitation of IgA antibodies at mucosal surfaces plays an important role in limiting spread of pathogens that primarily enter into systemic circulation via mucosal route. This has direct implication on the pathogenesis and the clinical outcome of the disease. We found significantly higher IgA responses in JOL2077 vaccinated chickens compared to the JOL2068 control group. This finding might explain why JOL2077 vaccinated chickens displayed insignificant inflammatory lesions in targeted tissues and achieved higher level of protection than control groups. Consistent with the notion that cellular immunity is protective against IBV infection [38], we next investigated cytokine and T cell recall responses elicited by our JOL2077 vaccine. After antigenic stimulation, CD4+ T cells secrete anti-viral cytokines, which generate potent antibody formation and CD8+ cytotoxic T lymphocytes, thus playing a critical role in controlling IBV infection [39, 40]. The present study demonstrated that chickens vaccinated with JOL2077 vaccine displayed higher CD4+ and CD8+ T cell responses compared to the control chickens. Consequently, the higher protection rate was observed in chickens vaccinated with JOL2077 vaccine. The nature of the cytokines produced after antigenic stimulation in vitro is an important parameter to define the type of immunity elicited [41]. Our data showed that JOL2077 vaccine induced efficient IBV-specific both Th1 (IFN-γ) and Th2 (IL-4 and IL-10) cytokine responses, which is indicative of the mixed-type of immunity. IFN-γ has potent anti-viral activity through the promotion of natural-killer cells and macrophage activation that are likely to contribute to the containment of IBV replication and spread within the host. Further, IFN-γ upregulates MHC-I and MHC-II molecules and induces IL-12, nitric oxide and superoxide production in macrophages, all of which are important in the elimination of intracellular pathogens [42]. These findings might further explain the higher protection rate observed in JOL2077 vaccinated chickens. The IL-4 and IL-10 have a clear role in promoting humoral immune responses [43, 44]. Previous studies have reported that IL-10 augment the proliferation and plasma differentiation of the B cells that have switched to IgA production, suggesting that IL-10 plays an important role in development of mucosal immunity. Considering that both humoral and cell-mediated immunity are important in containment of IBV replication and clinical disease, our study clearly shows that JOL2077 vaccine is capable of eliciting efficient humoral and CMI responses, and can, thus, offer significant protection against IBV in chickens.
The induction of SG-specific IgG responses following vaccination strongly correlates with the recovery from FT and protection from subsequent challenge infections in chickens [16, 28]. The systemic IgG mediate protection during the early stages of infection when SG circulates extracellularly before it penetrates the target cells [45]. The present study demonstrated that systemic IgG responses were highest in chickens vaccinated with either JOL2068 or JOL2077 strain compared to the PBS control group. Consequently, the mortality rate was lowest in both JOL2068 and JOL2077 vaccinated chickens compared to the chickens that received PBS only. The induction of immune responses at mucosal surfaces serves as the first line defense against pathogens that enter into systemic circulation through mucosal surfaces. Importantly, the presence of antigen-specific secretory (s) IgA antibodies in the intestinal mucus acts as an immunological barrier, thereby preventing the adherence of Salmonella bacteria to the intestinal lining and subsequent penetration into deeper tissues [46]. In the present study, we observed that the sIgA levels were highest in JOL2068 and JOL2077 vaccinated chickens. Consequently, the lower bacterial load was observed in spleen and liver of chickens vaccinated with either JOL2068 or JOL2077 strain. These findings clearly suggest that SG can be exploited to deliver viral antigens to elicit carrier and virus-specific humoral and cell mediated protective immune responses.
In conclusion, we show that SG delivering IBV-specific S1 protein can elicit efficient both SG and S1-specific humoral and cell-mediated immunity and can offer dual protection against FT and IB in chickens. Other studies have shown that efficacious live attenuated IB vaccines have protected chickens against clinical signs and mortality and increased significant resistance in birds to infection [40]. Our results clearly show that vaccination with SG-based IB vaccine induced efficient protection against IBV virulent challenge as evidenced by insignificant inflammation in trachea, lungs, kidneys and bursa and by efficient containment of viral replication in tracheal tissues. Further studies are warranted to investigate the protective efficacy of JOL2077 vaccine in both broiler and young layer chickens.
References
Cook JKA, Jackwood M, Jones RC (2012) The long view: 40 years of infectious bronchitis research. Avian Pathol 41:239–250
Colvero LP, Villarreal LYB, Torres CA, Brañdo PE (2015) Assessing the economic burden of avian infectious bronchitis on poultry farms in Brazil. Rev Sci Tech 34:993–999
Jackwood MW (2012) Review of infectious bronchitis virus around the world. Avian Dis 56:634–641
Ignjatović J, Sapats S (2000) Avian infectious bronchitis virus. Rev Sci Tech 19:493–508
Ruch TR, Machamer CE (2011) The hydrophobic domain of infectious bronchitis virus E protein alters the host secretory pathway and is important for release of infectious virus. J Virol 85:675–685
Yamada Y, Liu DX (2009) Proteolytic activation of the spike protein at a novel RRRR/S motif is implicated in furin-dependent entry, syncytium formation, and infectivity of coronavirus infectious bronchitis virus in cultured cells. J Virol 83:8744–8758
Cavanagh D (1983) Coronavirus IBV: structural characterization of the spike protein. J Gen Virol 64:2577–2583
Moore KM, Jackwood MW, Hilt DA (1997) Identification of amino acids involved in a serotype and neutralization specific epitope with in the s1 subunit of avian infectious bronchitis virus. Arch Virol 142:2249–2256
Cavanagh D (2007) Coronavirus avian infectious bronchitis virus. Vet Res 38:281–297
Wang L, Junker D, Collisson EW (1993) Evidence of natural recombination within the S1 gens of infectious bronchitis virus. Virology 192:710–716
Jia W, Karaca K, Parrish CR, Naqi SA (1995) A novel variant of avian infectious bronchitis virus resulting from recombination among three different strains. Arch Virol 140:259–271
Lin S-Y, Chen H-W (2017) Infectious bronchitis virus variants: molecular analysis and pathogenicity investigation. Int J Mol Sci 18:E2030
Shivaprasad HL (2000) Fowl typhoid and pullorum disease. Rev Sci Tech 19:405–424
Arora D, Kumar S, Jindal N, Narang G, Kapoor PK, Mahajan NK (2015) Prevalence and epidemiology of Salmonella enterica serovar Gallinarum from poultry in some parts of Haryana, India. Vet World 8:1300–1304
Lee S-K, Chon J-W, Song K-Y, Hyeon J-Y, Moon J-S, Seo K-H (2013) Prevalence, characterization, and antimicrobial susceptibility of Salmonella Gallinarum isolated from eggs produced in conventional or organic farms in South Korea. Poult Sci 92:2789–2797
Matsuda K, Chaudhari AA, Lee JH (2011) Evaluation of safety and protection efficacy on cpxR and lon deleted mutant of Salmonella Gallinarum as a live vaccine candidate for fowl typhoid. Vaccine 29:668–674
Pei Z, Jiang X, Yang Z, Ren X, Gong H, Reeves M, Sheng J, Wang Y, Pan Z, Liu F, Wu J, Lu S (2015) Oral delivery of a novel attenuated salmonella vaccine expressing influenza a virus proteins protects mice against H5N1 and H1N1 viral infection. PLoS One 10:e0129276
Kim JH, Hajam IA, Lee JH (2018) Oral immunization with a novel attenuated Salmonella Typhimurium encoding influenza HA, M2e and NA antigens protects chickens against H7N9 infection. Vet Res 49:12
Schepens B, De Vlieger D, Saelens X (2018) Vaccine options for influenza: thinking small. Curr Opin Immunol 53:22–29
Desin TS, Köster W, Potter AA (2013) Salmonella vaccines in poultry: past, present and future. Expert Rev Vaccines 12:87–96
Jiao H, Pan Z, Yin Y, Geng S, Sun L, Jiao X (2011) Oral and nasal DNA vaccines delivered by attenuated Salmonella enterica serovar Typhimurium induce a protective immune response against infectious bronchitis in chickens. Clin Vaccine Immunol 18:1041–1045
Ignjatovic J, Galli L (1994) The S1 glycoprotein but not the N or M proteins of avian infectious bronchitis virus induces protection in vaccinated chickens. Arch Virol 138:117–134
Kant A, Koch G, van Roozelaar DJ, Kusters JG, Poelwijk FAJ, van der Zeijst BAM (1992) Location of antigenic sites defined by neutralizing monoclonal antibodies on the S1 avian infectious bronchitis virus glycopolypeptide. J Gen Virol 73:591–596
Collisson EW, Pei J, Dzielawa J, Seo SH (2000) Cytotoxic T lymphocytes are critical in the control of infectious bronchitis virus in poultry. Dev Comp Immunol 24:187–200
Hajam IA, Lee JH (2017) An influenza HA and M2e based vaccine delivered by a novel attenuated Salmonella mutant protects mice against homologous H1N1 infection. Front Microbiol 8:872
Hajam IA, Lee JH (2017) Preexisting Salmonella -specific immunity interferes with the subsequent development of immune responses against the Salmonella strains delivering H9N2 hemagglutinin. Vet Microbiol 205:117–123
Bradford MM (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72:248–254
Hajam IA, Kim JH, Lee JH (2018) Incorporation of membrane-anchored fl agellin into Salmonella Gallinarum bacterial ghosts induces early immune responses and protection against fowl typhoid in young layer chickens. Vet Immunol Immunopathol 199:61–69
Larsen J, Lund O, Nielsen M (2006) Improved method for predicting linear B-cell epitopes. Immunome Res 2:2
Yrlid U, Svensson M, Håkansson A, Chambers BJ, Ljunggren HG, Wick MJ (2001) In vivo activation of dendritic cells and T cells during Salmonella enterica serovar Typhimurium infection. Infect Immun 69:5726–5735
Van TTH, Lin Y-C, Van TNN, Nguyen TQ, Le TTH, Do TH, Truong NH, Coloe PJ, Smooker PM (2013) Salmonella as a vaccine vector for influenza virus. Procedia Vaccinol 7:23–27
Rahman M, Uyangaa E, Woo Y (2012) Oral co-administration of live attenuated Salmonella enterica serovar Typhimurium expressing chicken interferon-α and interleukin-18 enhances the alleviation of clinical signs caused by respiratory infection with avian influenza virus H9N2. Vet Microbiol 157:448–455
Raggi LG, Lee GG (1965) Lack of correlation between infectivity, serologic response and challenge results in immunization with an avian infectious bronchitis vaccine. J Immunol 94:538–543
Gelb J, Nix WA, Gellman SD, Gellman SD (1998) Infectious bronchitis virus antibodies in tears and their relationship to immunity. Avian Dis 42:364–374
Toro H, Fernandez I (1994) Avian infectious bronchitis: specific lachrymal IgA level and resistance against challenge. Zentralbl Veterinarmed B 41:467–472
Thompson G, Mohammed H, Bauman B, Naqi S (1997) Systemic and local antibody responses to infectious bronchitis virus in chickens inoculated with infectious bursal disease virus and control chickens. Avian Dis 41:519–527
Cook KA, Otsuki K, da Silva Martins NR, Ellis MM, Huggins MB (1992) The secretory antibody response of inbred lines of chicken to avian infectious bronchitis virus infection. Avian Pathol 21:681–692
Chhabra R, Forrester A, Lemiere S, Awad F, Chantrey J, Ganapathy K (2015) Mucosal, cellular, and humoral immune responses induced by different live infectious bronchitis virus vaccination regimes and protection conferred against infectious bronchitis virus Q1 strain. Clin Vaccine Immunol 22:1050–1059
Seo SH, Collisson EW (1998) Cytotoxic T lymphocyte responses to infectious bronchitis virus infection. Adv Exp Med Biol 440:455–460
Raj GD, Jones RC (1997) Cross-reactive cellular immune responses in chickens vaccinated with live infectious bronchitis virus vaccine. Avian Pathol 26:641–649
O’Garra A, Arai N (2000) The molecular basis of T helper 1 and T helper 2 cell differentiation. Trends Cell Biol 10:542–550
Boehm U, Klamp T, Groot M, Howard JC (1997) Cellular responses to interferon-γ. Annu Rev Immunol 15:749–795
Estes DM, Brown WC (2002) Type 1 and type 2 responses in regulation of Ig isotype expression in cattle. Vet Immunol Immunopathol 90:1–10
Moore KW, O’Garra A, de Waal Malefyt R, Vieira P, Mosmann TR (1993) Interleukin-10. Annu Rev Immunol 11:165–190
Collins FM (1974) Vaccines and cell-mediated immunity. Bacteriol Rev 38:371–402
Pasetti MF, Simon JK, Sztein MB, Levine MM (2011) Immunology of gut mucosal vaccines. Immunol Rev 239:125–148
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
The work was designed by JHL and IAH. Experiments and sampling were done by IAH and JK. Manuscript draft preparation and statistical analysis was done by IAH. All authors read and approved the final manuscript.
Acknowledgements
This work was supported by Korea Institute of Planning and Evaluation for Technology in Food, Agriculture, and Forestry (IPET) through Agri-Bio industry Technology Development Program, funded by Ministry of Agriculture, Food and Rural Affairs (MAFRA) (Grant Number. 118055-03).
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Author information
Authors and Affiliations
Corresponding author
Additional files
Additional file 1.
In silico prediction of linear B cell epitopes and antigenicity of partial S1 amino acid sequence of IBV. Linear B-cell epitopes and antigenicity of SI protein were predicted by the BepiPred program, which assigns a score to each individual amino acid in a specific sequence. (A) Prediction of linear B cell epitopes. (B) Prediction of antigenicity using Kolaskar & Tongaonkar Antigenicity method.
Additional file 2.
Immunoblot analysis of S1 antigen expressed in JOL2077. The JOL2077 vaccine strain was grown in LB broth to mid-logarithmic phase and then the culture supernatant was collected and subjected to Western blot analysis using polyclonal IBV-specific antibody. Lane M, protein molecular weight (catalog#, P8500, GenDEPOT, USA); lane 1, JOL2077 culture supernatant; lane 2, JOL2068 culture supernatant as negative control.
Additional file 3.
Flow cytometric analysis of immune responses (A) Gating of lymphocytes in flow cytometric analysis of PBMCs isolated from the vaccinated and control chickens. The debris and dead cells were excluded by gating based on the forward and the side scatter. (B) Representative flow cytometry scatter dot plots for CD3+CD4+ and CD3+CD8+ T cell populations of vaccinated and control chicken groups.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Hajam, I.A., Kim, J. & Lee, J.H. Oral immunization with a novel attenuated Salmonella Gallinarum encoding infectious bronchitis virus spike protein induces protective immune responses against fowl typhoid and infectious bronchitis in chickens. Vet Res 49, 91 (2018). https://doi.org/10.1186/s13567-018-0588-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13567-018-0588-9