
Abstract
The abundance of domesticable mammals in Eurasia facilitated its early transition from hunter–gatherer to agricultural economies, with dramatic consequences for human history. This paper empirically examines the origins of these biogeographical advantages and finds that the extinction of large mammals during the past 100,000 years was a decisive force in the evolution of mammal domestication. In Eurasia’s domestication cradles, humans had sufficient incentives to continually practice herd management as a hunting strategy to prevent the depletion of their vital common resources. These strategies changed some targeted species and made them more receptive to human domination. The absence of these conditions (human incentive and animal receptivity) in other regions resulted in the paucity of domestication. The paper presents the most comprehensive empirical analysis of the origins of animal domestication and the roots of global inequalities to date and unearths a critical channel for the influence of deep history on comparative economic development.






Similar content being viewed by others


Notes
Galetti (2018) consider the impact of large mammals and their extinctions on the ice-age environment. Gamble (2013) presents an overview of climate change patterns over the past 7 million years. Koch and Barnosky (2006) and Mann (2019) provide excellent literature reviews on the causes of the Late Quaternary Extinctions. See Diamond (1997), Vigne (2011, 2015) and Zeder (2012a, 2012b) for analyses of the process of animal domestication. Section 2 discusses this literature.
Coevolution refers to a situation in which two or more species reciprocally affect each other’s evolution. The importance of coevolution for domestication was first proposed by Rindos (1984).
For example, in Africa, the cradle of humanity other animals coevolved with hominins for around 7 million years, and the LQE was negligible. Meanwhile, Homo sapiens’ occupation of the New World, where animals evolved in isolation, resulted in mass extinctions. Eurasia’s extinctions were comparatively mild, with more severe extinctions occurring at higher latitudes—regions that were occupied by hominins later during the ice ages. Section 2 discusses these issues further.
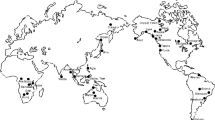
See Fig. 7 for the location of animal domestication centers.
The Pleistocene is the most recent period of repeated glaciations, lasting from approximately 2.58 million to 11,700 years ago. Polar ice caps appeared for the first time at the beginning of this geological epoch.
Laland (2015) discusses different approaches to biological evolution.
For example, Ashraf and Galor’s (2013) justification of their empirical strategy is that “distances along prehistoric human migration routes from Africa have no direct effect on economic development during the Common Era” (p. 15).
Domestication results in morphological changes, including docility, floppy ears, a rounder skull, reduced size, variation in coat color, and shorter birth intervals (Larson and Fuller 2014).
East Africa is the widely accepted origin of those Homo sapiens who colonized most of the planet in the Late Pleistocene (e.g., Nielsen 2017).
During the earlier out-of-Africa migration events, which started possibly as early as 185 ka, Homo sapiens did not colonize Eurasia, probably because of the widespread presence of Neanderthals in western Asia at that time (Lohse and Frantz 2014).
For this spatial weight matrix, the p-value from the Moran test of spatial independence is 0.00, which leads to the rejection of the null hypothesis that errors are independently and identically distributed.
For the probit model, F is the standard normal c.d.f.; the c.d.f. of complementary log–log model (\(C(y_i)=1-exp\{-exp(y_i)\}\)) is asymmetric around 0 (Cameron and Trivedi 2009).
For example, hunter–gatherers in southwest Asia were managing the core livestock species (the pig, sheep, goat, and cow) around 11 ka. In other regions, herd management becomes visible in archeological records generally later, for example, around 7 ka in Central Asia and 6 ka in South America. Larson et al. (2014, online appendix) provide data on the timing of change hunter–gatherers’ subsistence strategies based on the best available information as of 2014.
Consequently, the literature attributes the variation in the severity of the LQE to the differences in the extent of hominin–fauna coevolution and climate change (Sect. 2.3). Furthermore, if humans’ efforts to manage species were a cause of the LQE, extinctions should have been more severe in the lower latitudes of Asia, where most animal species were domesticated.
Here culture broadly defined as “large body of practices, techniques, heuristics, tools, motivations, values, and beliefs that we all acquire \(\cdots\) mostly by learning from other people” (Henrich 2016, p. 3).
For example, Mann (2019) argue that rapid and frequent climate changes during the ice ages created highly transient ecosystems that favored ecophysiological traits associated with large body size. The loss of these advantages within the stable climate regime of the Holocene lowered the extinction thresholds (the point at which a species experiences an abrupt change in density or number) of large mammalian species and made them vulnerable to proximate causes of extinction (e.g., humans). Thus, the shift in the fabric of the ecosystems during the Pleistocene–Holocene transition might be responsible for the LQE and the emergence of herd management among Homo sapiens.
The most widely used definition of megafauna is large mammals weighing more than 44\(_{kg}\). In Sandom (2014)’s data, 154 out of 177 species (or 87%) weigh more than 44\(_{kg}\).
These are Bolivia, Chile, China, Egypt, France, Indonesia, Iran, Iraq, Israel, Jordan, Kazakhstan, Lebann, Myanmar, Pakistan, Peru, Russia, Syria, Turkey, Ukraine, and United Arab Emirates. Figure 7 presents the locations of these countries.
For SAR models, the autocorrelation parameter (below the table) decreases from 0.8 in column 5 to 0.2 in column 6 and 7, which indicates that the set of independent variables considerably weaken the spatial correlation.
The values of the size and axis variable for Eurasia are, respectively, 54.76 (in millions of km\(^2\)) and 2.4, which are higher than those of other continents.
The sample size in these regressions decreases to 91 countries for which data on domesticable animals and the severity of extinction are simultaneously available.
The dependent variable is a binary variable that takes the value of 1 for animal domestication centers, and 0 otherwise.
The autocorrelation parameter is not statistically significant in columns 4 and 5, which indicates that spatial clustering is not a concern here.
Mean precipitation over the period 1961–1990 in the centers of domestication of Africa and southwest Asia is 5 and 30 (in mm per month), respectively.
Mean precipitation over the period 1961–1990 for non-achievers of Asia, Europe, and South Asia’s domestication centers is 82, 69, and 128 (in mm per month), respectively.
References
Acemoglu D, Robinson JA (2012) Why nations fail. Crown Business, New York
Anselin L (1988) Spatial econometrics: methods and models. Springer, Dordrecht
Ang JB (2013) Institutions and the long-run impact of early development. J Dev Econ 105:1–18
Ang JB (2019) Agricultural legacy and individualistic culture. J Econ Growth 24:397–425
Arbatli CE et al (2020) Diversity and conflict. Econometrica 88(2):727–797
Ashraf Q, Galor O (2013) The “Out of Africa” hypothesis, human genetic diversity, and comparative economic development. Am Econ Rev 103(1):1–46
Ashraf Q, Galor O (2018) The macrogenoeconomics of comparative economic development. J Econ Lit 56(3):1119–1155
Bartlett LJ et al (2016) Robustness despite uncertainty: regional climate data reveal the dominant role of humans in explaining global extinctions of Late Quaternary megafauna. Ecography 39:152–161
Bar-Yosef O (2011) Climatic fluctuations and early farming in West and East Asia. Curr Anthropol 52(Suppl. 4):S175–S193
Bettinger R, Richerson P, Boyd R (2009) Constraints on the development of agriculture. Curr Anthropol 50(5):627–631
Bond G et al (1999) The North Atlantic’s 1–2 kyr climate rhythm: relation to Heinrich events, Dansgaard/Oeschger cycles and The Little Ice Age. In: Clark PU, Webb RS, Keigwin LD (eds) Mechanisms of global change at millennial time scales. Geophysical monograph, vol 112. American Geophysical Union, Washington, pp 35–58
Borcan O, Olsson O, Putterman L (2018) State history and economic development: evidence from six millennia. J Econ Growth 23:1–40
Bove V, Gokmen G (2018) Genetic distance, trade, and the diffusion of development. J Appl Econom 33:617–623
Cameron CA, Trivedi PK (2009) Microeconometrics using stata. Stata Press, College Station
Cohen MN (1977) The food crisis in prehistory. Yale University Press, New Haven
Cohen MN (2009) Introduction: rethinking the origins of agriculture. Curr Anthropol 50(5):591–595
Comin D, Easterly W, Gong E (2010) Was the wealth of nations determined in 1000 BC? Am Econ J Macroecon 2:65–97
Cook JC (2015) The natural selection of infectious disease resistance and its effect on contemporary health. Rev Econ Stat 97:742–757
Crosby AM (1986) Ecological imperialism: the biological expansion of Europe, 900–1900. Cambridge University Press, Cambridge
Daniele V, Di Ruggiero A (2017) The roots of global inequality: the role of biogeography and genetic diversity. J Dev Stud 57:1584–1602
Darmofal D (2015) Spatial analysis for the social sciences. Cambridge University Press, Cambridge
Desmet K, Ignacio O-O, Romain W (2017) Culture, ethnicity, and diversity. Am Econ Rev 107(9):2479–2513
Dennell R (2017) Pleistocene hominin dispersal, naive faunas, and social networks. In: Boivin N, Crassard R, Petraglia M (eds) Human dispersal and species movement: from prehistory to the present. Cambridge University Press, Cambridge, pp 29–61
Depetris-Chauvin E, zak. (2020) The origins of the division of labor in pre-industrial times. J Econ Growth 25:297–340
Diamond J (1997) Guns, germs, and steel: the fates of human societies. Norton and Company, New York
Dow GK, Reed CG (2011) Stagnation and innovation before agriculture. J Econ Behav Organ 77:339–350
Dow GK, Reed CG (2015) The origins of sedentism: climate, population, and technology. J Econ Behav Organ 119:56–71
Dow GK, Reed CG, Olewiler N (2009) Climate reversals and the transition to agriculture. J Econ Growth 14:27–53
Ertan A, Fiszbein M, Putterman L (2016) Who was colonized and when? A cross-country analysis of determinants. Eur Econ Rev 83:165–184
Flannery KV (1969) Origins and ecological effects of early domestication in Iran and the Near East. In: Ucko PJ, Dimbleby GW (eds) The domestication and exploitation of plants and animals. Duckworth, London, pp 73–100
Galetti M et al (2018) Ecological and evolutionary legacy of megafauna extinctions. Biol Rev 93:845–862
Gamble C (2013) Settling the Earth, the archaeology of deep human history. Cambridge University Press, Cambridge
Gorodnichenko Y, Roland G (2017) Culture, institutions, and the wealth of nations. Rev Econ Stat 99(3):402–416
Hansen CW (2013) The diffusion of health technologies: cultural and biological divergence. Eur Econ Rev 64:21–34
Hemming SR (2004) Heinrich events: massive late Pleistocene detritus layers of the North Atlantic and their global climate imprint. Rev Geophys 42:1005
Henn B et al (2012) The great human expansion. Proc Natl Acad Sci 109(44):17758–17764
Henrich J (2016) The secret of our success: how culture is driving human evolution, domesticating our species, and making us smarter. Princeton University Press, Princeton
Koch PL, Barnosky AD (2006) Late Quaternary extinctions: state of the debate. Annu Rev Ecol Evol Syst 37:215–250
Laland K et al (2015) The extended evolutionary synthesis: its structure, assumptions and predictions. Proc R Soc Biol Sci 282:20151019
Larson G, Fuller DQ (2014) The evolution of animal domestication. Annu Rev Ecol Evol Syst 45:115–136
Larson G et al (2014) Current perspective and the future of domestication studies. Proc Natl Acad Sci 111:6139–6146
Lewis ME (2017) Carnivore guilds and the impact of hominin dispersals. In: Boivin N, Crassard R, Petraglia M (eds) Human dispersal and species movement: from prehistory to the present. Cambridge University Press, Cambridge, pp 29–61
Lohse K, Frantz LAF (2014) Neandertal admixture in Eurasia confirmed by maximum-likelihood analysis of three genomes. Genetics 196:1241–1251
Long JA, Stoy PC (2013) Quantifying the periodicity of Heinrich and Dansgaard–Oeschger events during marine oxygen isotope stage 3. Quatern Res 79:413–423
Malhi Y et al (2016) Megafauna and ecosystem function from the Pleistocene to the Anthropocene. Proc Natl Acad Sci 113(4):838–846
Mann DH et al (2019) Climate-driven ecological stability as a globally shared cause of Late Quaternary megafaunal extinctions: the plaids and stripes hypothesis. Biol Rev 94:328–352
Martin SP (1984) Prehistoric overkill: the global model. In: Martin PS, Klein RG (eds) Quaternary extinctions: a prehistoric revolution. University of Arizona Press, Tucson, pp 354–403
Milner AM et al (2016) Vegetation responses to abrupt climatic changes during the last interglacial complex (marine isotope stage 5) at Tenaghi Philippon, NE Greece. Quat Sci Rev 154:169–181
Nielsen R et al (2017) Tracing the peopling of the world through genomics. Nature 541:302–310
North DC, Thomas RP (1977) The first economic revolution. Econ Hist Rev 30(2):229–241
Hibbs DA, Olsson O (2004) Geography, biogeography and why some countries are rich and others are poor. Proc Natl Acad Sci 101(10):3715–3720
Olsson O, Hibbs DA (2005) Biogeography and long-run economic development. Eur Econ Rev 49(4):909–938
Ordonez A, Williams JW (2013) Climatic and biotic velocities for woody taxa distributions over the last 16,000 years in eastern North America. Ecol Lett 16:773–781
Olsson O, Paik C (2016) Long-run cultural divergence: evidence from the Neolithic Revolution. J Dev Econ 122:197–213
Ostrom E (1990) Governing the commons: the evolution of institutions for collective action. Cambridge University Press, New York
Petraglia M (2017) Hominins on the move: an assessment of anthropogenic shaping of environments in the Palaeolithic. In: Boivin N, Crassard R, Petraglia M (eds) Human dispersal and species movement: from prehistory to the present. Cambridge University Press, Cambridge, pp 90–118
Price TD, Bar-Yosef O (2011) The origins of agriculture: new data, new ideas an introduction to supplement 4. Curr Anthropol 52(Suppl. 4):S163–S174
Putterman L (2008) Agriculture, diffusion, and development: ripple effects of the Neolithic Revolution. Economica 75:729–748
Putterman L, Weil DN (2010) Post-1500 population flows and the long-run determinants of economic growth and inequality. Q J Econ 125(4):1627–82
Ramachandran et al (2005) Support from the relationship of genetic and geographic distance in human populations for a serial founder effect originating in Africa. In: Proceedings of the National Academy of Sciences 102(44):15942
Riahi I (2017) Colonialism and genetics of comparative development. Econ Hum Biol 27:55–73
Riahi I (2020a) How hominin dispersals and megafaunal extinctions influenced the birth of agriculture. J Econ Behav Organ 174:227–250
Riahi I (2020b) Animals and the prehistoric origins of economic development. Eur Rev Econ Hist. https://doi.org/10.1093/ereh/heaa016
Rindos D (1984) The origins of agriculture: an evolutionary perspective. Academic Press, Orlando
Sandom C et al (2014) Global Late Quaternary megafauna extinctions linked to humans, not climate change. Proc R Soc 281:2013–3254
Smith Bruce D (2007) Niche construction and the behavioral context of plant and animal domestication. Evol Anthropol 16:188–199
Smith Vernon L (1975) The primitive hunter culture, Pleistocene extinction, and the rise of agriculture. J Polit Econ 83:727–755
Spolaore E, Wacziarg R (2009) The diffusion of development. Q J Econ 124(2):469–529
Spolaore E, Wacziarg R (2013) How deep are the roots of economic development? J Econ Lit 51(2):325–369
Spolaore E, Wacziarg R (2016) War and relatedness. Rev Econ Stat 98(5):925–939
Stiner MC (2001) Thirty years on the broad spectrum revolution and paleolithic demography. Proc Natl Acad Sci 98(13):6993–6996
Steiper ME, Young NM (2006) Primate molecular divergence date. Mol Phylogenet Evol 41(2):384–394
Stuart AJ (2014) Late Quaternary megafaunal extinctions on the continents: a short review. Geol J 50:338–363
Strahler AN, Stahler AM (1992) Modern physical geography. Wiley, New York
Surovell TA et al (2016) Test of Martin’s overkill hypothesis using radiocarbon dates on extinct megafauna. Proc Natl Acad Sci 113:886–891
Vigne J-D (2011) The origins of animal domestication and husbandry: a major change in the history of humanity and the biosphere. CR Biol 334:171–181
Vigne J-D (2015) Early domestication and farming: what should we know or do for a better understanding? Anthopozoologia 50(2):123–150
Weisdorf JL (2005) From Foraging to Farming: Explaining the Neolithic Revolution. J Econ Surv 19:561–586
Zeder MA (2012) The domestication of animals. J Anthropol Res 68:161–90
Zeder MA (2012) The broad spectrum revolution at 40: resource diversity, intensification, and an alternative to optimal foraging explanations. J Anthropol Archaeol 31:241–264
Zeder MA (2015) Core questions in domestication research. Proc Natl Acad Sci 112:3191–3198
Zeder MA (2016) Domestication as a model system for niche construction theory. Evol Ecol 30:325–348
Acknowledgements
I am very grateful to Gregory K. Dow and Sanders Korenman for their help and support throughout this project. I thank Daron Acemoglu, Anna D’Souza, Frank Heiland, Louis Putterman, Melinda A. Zeder, the participants of brown-bag seminars at the Marxe School of Public and International Affairs (Spring 2019), and the participants of the Growth Lab seminars at the Department of Economics in Brown University (December 2019) for comments and suggestions on different drafts of the paper.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Appendices
Appendix 1
See Figs. 7, 8, 9, 10 and Tables 5, 6, 7, 8.

Source: Larson and Fuller (2014)
Distribution of the wild ancestors of domesticable animals.

Some key events in hominin history

Late Quaternary extinctions

Ecological diversity
Appendix 2: Data descriptions and resources
1.1 Absolute latitude
The absolute value of a country’s geodesic centroid latitude. Source: The CIA’s World Factbook.
1.2 Climate-c1 and climate-c2
The first two principal components of mean temperature anomaly, mean precipitation velocity, mean temperature, and mean precipitation. The first two components cumulatively explain 84% of these four measures. The following table presents the principal component analysis of the measures of climate.
The principal components of climate variables | ||||
Component variables | PC1 | PC2 | PC3 | PC4 |
Eigenvalue | 2.7 | 1.08 | 0.39 | 0.26 |
Proportion | 0.57 | 0.27 | 0.10 | 0.07 |
Mean temperature anomaly | − 0.51 | 0.51 | 0.05 | 0.70 |
Mean precipitation velocity | 0.47 | 0.54 | 0.70 | − 0.10 |
Mean temperature | 0.54 | − 0.43 | 0.08 | 0.72 |
Mean precipitation | 0.48 | 0.51 | − 0.72 | 0.02 |
1.3 Continent axis
Continental major axis is a rough measure of the East-West orientation of the major landmasses and is obtained by dividing each continent’s distance in longitudinal degrees between the eastern and westernmost points with its North-West distance in latitudinal degrees. Source: Hibbs and Olsson (2004) and Olsson and Hibbs (2005).
1.4 Continent size
The total area (in millions of km2) of a continent to which a country belongs. Total areas of continents are 30, 55, 7.69, 43, millions of km2 for Africa, Eurasia, Australia, and the Americas, respectively. Source: The World Wide Web.
1.5 Domesticable animals (Larson and Fuller)
The number of the wild progenitors of animal species, with traits that were suitable for domestication, that were prehistorically native to modern-day countries. Domesticable animals weighing more than 10 kg are Sus scrofa (wild boar), Ovis aries (sheep), Capra hircus (goat), Bos taurus (cow), Equus caballus (horse), Camelus dromedarius (Arabian camel), Camelus bactrianus (Bactrian camel), Equus asinus (donkey), Bubalus bubalis (water buffalo), Bos gaurus (gaur), Bos javanicus (Bali cattle), Bos grunniens (yak), Lama glama and Vicugna pacos (llama and alpaca), Rangifer tarandus (reindeer), and Canis familiaris (Dog). Figure 7 presents the geographic distribution of these species .Source: Larson and Fuller (2014).
1.6 Domesticable animals (Olsson and Hibbs)
The number of domesticable animal species weighing more than 44 kg that were prehistorically native to the territories that now belong to modern-day countries. These species are Sus scrofa (wild boar), Ovis aries (sheep), Capra hircus (goat), Bos taurus (cow), Equus caballus (horse), Camelus dromedarius (Arabian camel), Camelus bactrianus (Bactrian camel), Equus asinus (donkey), Bubalus bubalis (water buffalo), Bos gaurus (gaur), Bos javanicus (Bali cattle), Bos grunniens (yak), Lama glama and Vicugna pacos (llama and alpaca), and Rangifer tarandus (reindeer). Source: Hibbs and Olsson (2004) and Olsson and Hibbs (2005).
1.7 Elevation (mean)
The mean elevation of a country in km above sea level, calculated using geospatial elevation data reported by the G-ECON project at a 1-degree resolution. The measure is thus the average elevation across the grid cells within a country. Source: Ashraf and Galor (2013).
1.8 Elevation (standard deviation)
The standard deviation of elevation across the grid cells within a country in km above sea level, calculated using geospatial elevation data reported by the G-ECON project at a 1-degree resolution. Source: Ashraf and Galor (2013).
1.9 Ecological diversity
A country-level measure indicating the extent of variation in countries’ physical geography as defined by Köppen–Geiger climate zones. These zones are tropical rainforest climate (Af), monsoon variety of tropical rainforest climate (Am), tropical savannah climate (Aw), steppe climate (BS), desert climate (BW), mild human climate with no dry season (Cf), mild human climate with a dry season (Cs), mild human climate with a dry winter (Cw), snowy-forest with a dry winter (DW), snowy-forest with a most winter (Df), polar ice climate (E), and highland climate (H). Let \(s_i^K\in [0,1]\) denotes the share of the land area of country i in each of the 12 Köppen–Geiger climate zones \(K\in \{Af,Am,Aw,\ldots ,H\}\). The value of zero for \(s_i^K\) indicates that country i’s land area excludes climate zone K, and the value of one indicates that the entire land area of country i is only in one climate zone. Let \(var(s_i^K)\) denotes the sample variance of \(s_i^K\) for country i. When most of the landmass of a country is in only a few climate zones, \(s_i^K\)s are dispersed and, therefore, \(var(s_i^K)\) are high. For example, for countries such as Qatar, Kuwait, Egypt, and Oman, \(s^{BW}\)=1, which means that their entire land areas are in the desert climate (BW). For these countries \(var(s_i^K)\)=0.08, which is the highest variance of \(s_i^K\) in the sample of 134 countries. On the other hand, when the landmass of a country includes many climate zones, \(s_i^K\)s are less dispersed and \(var(s_i^K)\) is relatively small. For example, China’s landmass includes nine climate zones and \(var(s_{China}^K)\)=0.01, which is the smallest value for \(var(s_i^K)\) in the entire sample. The measure of ecological diversity of country i is defined as \(di=1/ var(s_i^K)\) for \(K\in \{Af,Am,Aw,\cdots ,H\}\). Thus, if country j has more diversity in its physical geography than country i, \(var(s_j^K)< var(s_i^K)\) and \(d_i>d_j\). The measure of ecological diversity (di) is rescaled to 0 and 1, with 0 and 1 indicating the lowest and the highest ecological diversity, respectively. Summary statistics for the share of the landmass of countries in Köppen–Geiger climate zones (\(s_i^K\)), \(var(s_i^K)\), and di are presented in Table 8. Figure 10 presents the global variation in the normalized measure of ecological diversity. The original source of the Köppen–Geiger climate zones map is Strahler and Stahler's (1992) Modern Physical Geography. The share of the land area of countries in Köppen–Geiger climate zones is obtained online from: https://www.pdx.edu/econ/country-geography-data.
1.10 Extinction
This variable refers to the percentage of known extinct animal species (larger than 10 kg) to the total number of extinct and extant animal species from the Late Pleistocene to early/middle Holocene (132,000–1000 years BP) for each country. Sandom (2014) calculates the proportion of extinct large mammals for a total of 177 taxonomically accepted animal species in the literature. They estimate these extinctions for Taxonomic Databases Working Group (TDWG) countries. With some exceptions, TDWG regional classification corresponds to countries’ current borders. The exceptions are large countries such as Russia, China, USA, Brazil, Australia, India, Argentina, Mexico, South Africa, and Chile, for which data are available at the within-country level. For these countries, I calculate the average rate of extinction at the country-level. Within-country variation in the severity of the extinction for the large countries is very limited and averaging the measure of extinctions the country-level does not affect the main results. Source: Sandom (2014).
1.11 Genetic diversity
The expected heterozygosity (genetic diversity) of a given country as predicted by (the extended sample definition of) migratory distance from East Africa (i.e., Addis Ababa, Ethiopia). This measure is calculated by applying the regression coefficients obtained from regressing expected heterozygosity on migratory distance at the ethnic group level, using the worldwide sample of 53 ethnic groups from the HGDP-CEPH Human Genome Diversity Cell Line Panel. This measure is normalized to 0–1 range with 0 and 1 indicating the lowest and highest genetic diversity, respectively. Source: Ashraf and Galor (2013).
1.12 Migratory distance from East Africa
The great circle distance from Addis Ababa (Ethiopia) to the country’s modern capital city along a land-restricted path forced through one or more of five intercontinental waypoints. Distances are calculated using the Haversine formula and are measured in units of 1000 km. The geographical coordinates of the intercontinental waypoints are from Ramachandran et al. (2005), while those of the modern capital cities are from the CIAs World Factbook. Source: Ashraf and Galor (2013).
1.13 Precipitation velocity
A measure of displacement of mean annual precipitation between the Last Glacial Maximum (21 ka) and present-day climate (1950-2000). Climate velocity indicates the speed at which species will need to migrate in order to stay in the same enveloped climatic condition. The velocity of climate change is generally calculated as the temporal trend divided by spatial gradient in a climate variable such as temperature or precipitation. More specifically, climate velocity = \(\Delta\) (climate variable)/year \(\div\) \(\Delta\) (climate variable)/km (Ordonez and Williams, 2013). Mean precipitation velocity between the Last Glacial Maximum (the LGM, 21 ka) and present-day climate (1950–2000) is calculated from the WorldClim’s data at 2.5 arcminutes and is presented in absolute value and is standardized to fall between 0 and 1, with 0 and 1 representing the lowest and the highest velocity, respectively. Source: Sandom (2014).
1.14 Terrain roughness
The degree of terrain roughness of a country, calculated using geospatial surface undulation data reported by the G-ECON project at a 1-degree resolution, which is based on more spatially disaggregated elevation data at a 10-min resolution. The measure is thus the average degree of terrain roughness across the grid cells within a country. Source: Ashraf and Galor (2013).
1.15 Temperature anomaly
A measure of the magnitude of climate change between the Last Glacial Maximum (21 ka) and present-day climate (1950–2000). The anomaly in temperature means a departure from a reference value. A positive anomaly indicates that the observed measure was higher than the reference value, while a negative anomaly indicates that the observed measure was lower than the reference value. Mean temperature anomaly between the Last Glacial Maximum (the LGM, 21 ka) and present-day climate (1950–2000) is calculated from the WorldClim’s data at 2.5 arcminutes. This measure is constructed by averaging records from the MIROC3 and CCSM models and is presented in absolute value and is standardized to fall between 0 and 1, with 0 and 1 representing the lowest and the highest anomaly, respectively. Source: Sandom (2014).
1.16 Temperature and precipitation
The intertemporal average monthly temperature and precipitation of a country in, respectively degrees Celsius and mm per month, over the 1961–1990 time period, calculated using geospatial average monthly data for this period reported by the G-ECON project at a 1-degree resolution. The measure is thus the spatial mean of the intertemporal average monthly temperature and precipitation across the grid cells within a country. Source: Ashraf and Galor (2013).
See Table 9.
Rights and permissions
About this article

Cite this article
Riahi, I.A. Why Eurasia? A probe into the origins of global inequalities. Cliometrica 16, 105–147 (2022). https://doi.org/10.1007/s11698-021-00222-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11698-021-00222-9