
Abstract
Porcine parvovirus 1 (PPV1) mainly induces severe reproductive failure in pregnant swine, and causes huge economic losses to the swine industry. Cell apoptosis induced by PPV1 infection has been identified the major cause of reproductive failure. However, the molecular mechanism was not fully elucidated. In this study, the potential mechanism of PPV1 induced apoptosis in PK-15 cells was investigated. Our results showed that PPV1 induced apoptosis in PK-15 cells. Further studies revealed toll-like receptor 2 (TLR2) was involved in the PPV1-mediated apoptosis. TLR2 siRNA significantly decreased the apoptosis. Finally, our study showed NF-κB was activated by TLR2 during PPV1-induced apoptosis. The activation of NF-κB signaling was demonstrated by the phosphorylation of p65, p65 nuclear translocation and degradation of inhibitor of kappa B α (IκBα). Together, these results provided evidence that the recognition between PPV1 and PK-15 cells was mainly through TLR2, and then induction of the NF-κB signaling pathway activation, which further induces apoptosis. Our study could provide information to understand the molecular mechanisms of PPV1 infection.
Similar content being viewed by others

Introduction
Parvoviruses are small, non-enveloped, and single-stranded DNA viruses in the family Parvoviridae (Cao et al. 2017). To date, seven porcine parvovirus (PPV) genotypes (PPV1 to PPV7) have been discovered in swine (Li et al. 2021). Classical PPV1 is an important pathogen causing reproductive failure in sows, characterized by early embryonic death, fetal mummification, stillbirths, infertility and delayed return to estrus (Zhang et al. 2015; Zhou et al. 2017). Unlike PPV1, the research of novel PPVs (PPV2-PPV7) is limited to virus isolations, and the pathogenesis of PPV2-PPV7 is unclear (Li et al. 2021). Once pregnant sows infect with PPV1, the virus can across placental barrier and destroy their embryos or fetuses. PPV1 primarily replicates in rapidly dividing cells, which can not induce mitosis. A series of ultrastructural changes after PPV1 infection can be observed, such as nuclear swelling, rapid lactate dehydrogenase and free viral DNA releasing, all point toward apoptosis and necrosis as the main form during virus infection (Meszaros et al. 2017a). Zhang et al. agreed that the apoptosis induced by PPV1 is considered to be the key factor of cell death and tissue damage, which are important pathogenic factors of reproductive failure in sows (Zhang et al. 2019). However, the molecular mechanism of PPV1 inducing apoptosis remains unclear.
Nuclear factor-kappa B (NF-κB) is a highly conserved protein complex that involves in inflammation, immunity and cancer (Taniguchi and Karin 2018). Research showed that NF-κB is activated and subsequent “proinflammatory cytokine storm” causes serious damage after virus infection. Inhibitors blocking the activity of NF-κB signaling pathway provide a potential therapeutic target to treat virus infection. Dexamethasone, an NF-κB inhibitor, has succeeded in treating COVID-19 for reducing the incidence of death (Davies et al. 2021). Some selected small molecule NF-κB inhibitors are effective at reducing adjuvant-induced inflammation, and act as immune potentiators for enhancement of vaccine adjuvants (Moser et al. 2020).Upon viral infection, pattern recognition receptors (PPRs) recognize pathogen-associated molecular pattern (PAMPs). The compounds engage TIR-domain-containing adaptor inducing interferon-β (TRIF), myeloid differentiation primary response gene 88 (MyD88), or mitochondria antiviral-signaling protein (MAVS). Then these adaptor molecules recruit TNF receptor-associated factor (TRAF) and transforming growth factor β-activated kinase 1 (TAK1) to active NF-κB (Zhao et al. 2015).
Toll-like receptors (TLRs), a well-characterized cluster of prototype PRRs, play a crucial role in the initiation of innate cellular immune responses. TLRs bind to components of pathogens, and produce cytokine through activation of signaling pathways (Vandevenne et al. 2010). TLRs and their downstream signaling pathways play crucial roles in host innate and adaptive immune responses (Gray et al. 2006). However, little is known about the molecular mechanisms of PPV1-induced TLRs/NF-κB activation.
In the present study, TLR2 was verified to be involved in PPV1-induced apoptosis using RNA interference (RNAi). Subsequently, the activation of NF-κB signaling pathway during PPV1 infection was confirmed by dual-luciferase reporter test, indirect immunofluorescence assay (IFA) and western blotting. These results may provide valuable information on the host cell-virus interactions for PPV1 and other swine reproductive failure viral infections.
Materials and methods
Cells and virus
The PK-15 cell line (ATCC: CCL-33) was used to propagate PPV1. PK-15 cells were cultured in minimum eagle’s medium (MEM, Gibco, USA) supplemented with 10% fetal bovine serum (FBS, Gibco, USA) at 37 °C in a humidified atmosphere containing 5% CO2. The PPV1 HN-4 strain was isolated from the lung tissue of an embryonic death piglet in Henan, China (Zhou et al. 2017). The cytopathic effect (CPE) and immunofluorescence assay were observed in PK-15 cells after five blind passages of PPV1. The TCID50 was 10–5.1/0.1 mL.
Inhibitor and antibodies
The NF-κB inhibitors BAY 11-7082 was purchased from Beyotime company (Beyotime, China) and dissolved in Dimethyl sulfoxide (DMSO, Solarbio, China) to a stored concentration of 100 μM. The final working concentration of NF-κB inhibitor BAY 11-7082 is 0.5 μM and 5 μM. Monoclonal antibodies against phosphorylated p65, IκBα and IKKα were purchased from Cell Signaling Technology (CST, USA). Monoclonal antibodies against TBP and β-actin were purchased from Proteintech (Proteintech, China).
Apoptosis analysis
Annexin V/PI double staining was used to detect phosphatidylserine valgus to evaluate apoptosis. In the early stage of cell apoptosis, phosphatidylserine in the inner side of the cell is everted to the outer membrane and can bind with Annexin V- Fluorescein isothiocyanate (FITC). In the late stage of apoptosis, propidium iodide (PI) can bind to DNA through the damaged cell membrane. According to Annexin V-FITC/PI apoptosis decision kit (Solarbio, China), the cells infected with PPV1 were stained, and then the apoptosis was detected by flow cytometry (Beckman Coulter, USA). Cell apoptosis was analyzed using CytExpert software (Beckman Coulter, USA).
NF-κB luciferase assays
PK-15 cells were seeded in 24-well plates. Cell (reached approximately 70% - 80% confluence) were used to transfection experiment. Briefly, NF-κB reporter plasmid pNF-κB-luc (125 ng) and indicated plasmids pRL-TK (25 ng) were added to 100 μL of Opti-MEM® Reduced serum medium (ThermoFisher Scientific, USA). Then 0.5 μL of PLUSTM reagent (ThermoFisher Scientific, USA) was added, and incubated for 5 min at room temperature. 1.5 μL of Lipofectamine®LTX reagent (ThermoFisher Scientific, USA) was subsequently added to the mixture, and incubated for an additional 25 min at room temperature. This mixture was added to cells and incubated at 37 °C. 24 h later, cells were infected with PPV1 (MOI = 0.1/1/10). Cell extracts were collected, and the luciferase activities were measured using the Dual-Luciferase Reporter assay system according to the manufacturer's protocols (Promega, USA). Values for each sample were normalized using the renilla luciferase values. For each experiment, firefly luciferase activity was divided by renilla luciferase activity to correct for differences in transfection efficiency. The resultant ratios were normalized to fold change value obtained from PK-15 infected with PPV1.
Western blotting analysis
PK-15 cells were cultured in 60 mm dishes and were infected with PPV1 (MOI = 1). At the indicated times, cytoplasmic and nuclear protein were extracted separately using Cytoplasmic and Nuclear Protein Extraction Kit (TDY, China). The total protein was prepared by adding lysis buffer in cells. The cell extracts were separated by SDS-PAGE and electroblotted onto a polyvinylidene difluoride (PVDF) membrane (Millipore, USA). Then the membranes were incubated with specific primary antibodies, followed by incubation with appropriate horseradish peroxidase (HRP) conjugated secondary antibodies. Signals were detected using an ECL chemiluminescence detection kit (Millipore, USA) and quantitated using Image J software.
Immunofluorescence assay (IFA)
PK-15 cells were seeded in 96-well culture plate. At approximately 50%–60% confluence, cells were infected with PPV1 (MOI = 1) for 1.5 h. Maintenance medium (MEM containing 2% FBS) was added for 36 h. Then PK-15 cells were fixed with 4% paraformaldehyde for 30 min, permeabilized with 3% Triton X-100, and blocked with 3% BSA (Sigma, USA) for 30 min. Cells were stained with NF-κB p65 mouse monoclonal antibody (5G6) (TDY, China) for 1 h at room temperature. Cells were washed with PBS and incubated with goat anti-mouse secondary antibodies conjugated with Alexa-fluor 594 (Life Technologies, USA) for 1 h. Nuclei were stained with 4′-6-diamidino-2-phenylindole-dihydrochloride (DAPI, Invitrogen, USA) for 5 min at room temperature. Images were acquired by fluorescence microscope (Zeiss, Germany).
The RNAi analysis
Two short interfering RNA (siRNA) sequences targeting the porcine TLR2 (siRNA210 and siRNA662) and one irrelevant interference sequence (negative control, Table 1) were designed separately (based on GenBank: NM_213761.1). Sequences were synthesized by GenePharma Company. Cells were transfected with siRNA using ExFect®2000 Transfection Reagent (Vazyme, China) according to the manufacturer’s instructions. RNAs were extracted from cells after 24 h, 36 h and 48 h post-transfection. Protein was collected at 48 h post-transfection.
Real-time quantitative reverse transcription polymerase chain reaction (RT-qPCR)
RT-qPCR was used to detect of TLR2 and NF-κB signal molecule during PPV1 infection. The cells were collected and analyzed at different time points after PPV1 infection. Total RNA was extracted from PK-15 cells using TRIzol reagent (Vazyme, China). cDNAs were reversed by Strand cDNA Synthesis Kit (Vazyme, China). RT-qPCR was performed using Power SYBR Green PCR Master Mix (Vazyme, USA). β-actin was used as the internal control, and all gene expressions were normalized to β-actin. The primers used are listed in Table 2.
Statistical analysis
All experiments were performed in triplicate. Data are presented as mean ± standard deviation (SD). P values were calculated using Student’s t test or one-way analysis of variance (ANOVA). * 0.01 < p < 0.05, ** p < 0.01.
Results
PPV1 infection induces apoptosis in PK-15 cells
To explore whether PPV1 infection could induce apoptosis, PK-15 cells were infected with different MOIs (0.1 and 1) of PPV1. After 36 h, the cells were stained with Annexin V-FITC/PI, and then analyzed by flow cytometry. The results showed that the percentage of apoptotic cells in PPV1 infected groups were significantly higher than that in the mock group. At the MOI of 0.1 and 1, apoptotic cells induced by PPV1 increased about 2 and 4.1-fold, respectively ( . 1).

PPV1 infection induces apoptosis in PK-15 cells. PK-15 cells were infected with PPV1 at MOI of 0.1 and 1 for 36 h. Mock-infected PK-15 cells were used as controls. PPV1-induced phosphatidylserine eversion detected by annexin V-FITC/PI staining and flow cytometry (a) and presented as a percentage of apoptotic cells (b), respectively. Results are expressed as mean ± SD from three independent experiments. *0.01 < p < 0.05, **p < 0.01
TLR2 is involved in PPV1-induced apoptosis
To determine whether TLR2 involved in PPV1-induced apoptosis, RT-qPCR and RNAi analyses were used. mRNA expression of TLR2 had a significant peak at 6 hpi and 12 hpi in PPV1-infected cells comparing to uninfected cells (Fig. 2a). To further understand the role of TLR2, TLR2 was silenced by RNAi. We designed two specific siRNA sequence, siRNA 210 and siRNA 662. As shown in Fig. 2b and c, siRNA 210 significantly decreased the mRNA and protein expression level of TLR2. We next investigated whether TLR2 was involved in PPV1 infection. mRNA of TLR2 was significantly reduced in PPV1-infected cells after blocking the expression of TLR2 (Fig. 2d). siRNA 210 treated cells were infected with PPV1 (MOI = 1), and stained with an Annexin V-FITC/PI kit. Results showed that the percentage of apoptotic cells were significantly reduced in PPV1-infected TLR2 knockdown cells, which suggested TLR2 was involved in PPV1-mediated apoptosis (Fig. 3a and b).

TLR2 is involved in the PPV1-mediated NF-κB pathway. (a) PK-15 cells were infected with PPV1 at MOI of 1, and cell lysates were harvested at the indicated time points. The mRNA expression levels of TLR2 were analyzed by RT-qPCR. (b) and (c) Silencing efficiency of siRNA targeting TLR2 were detected by RT-qPCR and western blotting. PK-15 cells were transfected with negative control siRNA or TLR2 siRNA. Total RNA was extracted, and the mRNA levels of TLR2 were measured by RT-qPCR. Cell lysates were harvested and analyzed by western blotting. (d) PK-15 cells were transfected with negative control siRNA or TLR2 specific siRNAs, then infected with PPV1 at an MOI of 1. The mRNA expression levels of TLR2 was analyzed by RT-qPCR. Results are expressed as mean ± SD from three independent experiments. *0.01 < p < 0.05, **p < 0.01

TLR2 is involved in apoptosis induced by PPV1 infection. PK-15 cells were infected with PPV1 at MOI of 1 and transfected with siRNA (150 pmol) or siNegative (150 pmol) as indicated. At 36 hpi, the percentage of apoptotic cells was detected by flow cytometry analysis using an Annexin V-FITC/PI apoptosis detection kit. Results are expressed as mean ± SD from three independent experiments. *0.01 < p < 0.05, **p < 0.01
PPV1 infection active NF-κB in PK-15 cells
To investigate whether NF-κB signaling was activated during PPV1 infection, PK-15 cells were co-transfected with pNF-κB-luc and pRL-TK, and infected with various MOIs of PPV1 later. The luciferase activity was no statistically significant difference between the PPV1 (MOI = 0.1) infected and control cells. While the luciferase activity was significantly increased about 1.8 and 2.1-fold at MOI of 1 and 10, respectively. To further confirm NF-κB was activated by PPV1 infection, PPV1 were inactivated with UV. Data showed that PPV1 pretreated with UV did not activate NF-κB signaling (Fig. 4a). mRNA level of NF-κB was also decreased when PK-15 cells treated with siRNA 210 (Fig. 4b).

PPV1 infection promotes NF-κB activation by TLR2 in PK-15 cells. (a) PK-15 cells were co-transfected with pRL-TK and pNF-κB-luc expression plasmids for 24 h. Cells were mock treated or stimulated with different MOI (0.1, 1, or 10) of PPV1 or inactivated PPV1 for 24 h, and cell lysates were analyzed for dual-luciferase reporter assays. The ratios were normalized to fold change value. (b) PK-15 cells were transfected with negative control siRNA or TLR2 specific siRNAs, then infected with PPV1 at an MOI of 1. The mRNA expression levels of NF-κB were analyzed by RT-qPCR. Results are expressed as mean ± SD from three independent experiments. *0.01 < p < 0.05, **p < 0.01
PPV1 infection activates NF-κB associating with p65 and IκBα axis
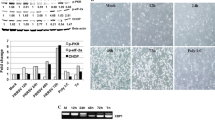
NF-κB signaling was activated in PK-15 cells upon PPV1 infection demonstrating by the phosphorylation of p65 (p-p65), p65 nuclear translocation and degradation of IκBα. The expression of p65 phosphorylated at serine 536 was analysed by western blotting in the indicated time. Compared to the mock cells, the expression levels of total p-p65 increased after PPV1 initial infection, peaked at 6 h, follow by a decrease (Fig. 5a). The translocation of p-p65 to nucleus was significantly increased from 3 h post-infection (hpi) to 24 hpi (Fig. 5b). Instead, the p-p65 in the cytoplasm was decreased (Fig. 5c).

PPV1 infection activates NF-κB associating with p65 and IκBα axis. PK-15 cells were mock treated or stimulated with PPV1 (MOI = 1). Cells were harvested at 0, 3, 6, 12, 24, 36, 48 and 60 hpi. The total protein (a), cytoplasmic protein (b), and nuclear protein (c) were subjected to western blotting analysis with antibodies against phosphorylated p65, β-actin, and TBP, respectively. (d) After 36 hpi, cells were fixed and permeated for IFA. Red: p65 protein; Blue: DAPI; magnification, × 200. (e) The cells lysates were processed for western blotting analysis with antibodies against IκBα and IKKα. All experiments were performed in triplicate and repeated three times. *0.01 < p < 0.05, **p < 0.01
To investigate the nuclear translocation of p65, PK-15 cells were infected with PPV1 (MOI = 1) and fixed at 36 hpi. P65 was observed with red fluorescence following stimulation with PPV1, and located on nuclear and cytoplasm. The amounts of p65 translocated into nuclei was more than uninfected cells, indicating PPV1 contributes p65 nuclear translocation (Fig. 5d).
Normally, NF-κΒ binds to the inhibitor of NF-κΒ (IκΒ) to form an inactivated compound, and sterically block the function of NF-κΒ in plasma. Therefore, we used an anti-IκΒα antibody to analyse IκΒ expression. As shown in Fig. 5e, there was a significant decrease in IκBα protein levels at 12, 36, 48 and 60 hpi in PPV1 infected PK-15 cells. IκB kinase (IKK) triggers the phosphorylation and degradation of IκB. The results showed IKKα increased gradually since 3 hpi, and peaked at 12 hpi, and then reduced.
PPV1 induces apoptosis by activating the NF-κB signaling
NF-κB signaling plays a key role in the regulation of apoptosis. Whether the apoptosis of PK-15 cells induced by PPV1 infection was investigated. PK-15 cells were treated with different concentrations (0.5 μM and 5 μM) of NF-κB inhibitor BAY 11-7082 for 0.5 h, and then infected with PPV1 (MOI = 1). The percentage of apoptotic cells were remarkably reduced in BAY 11-7082 treated PPV1-infected cells (Fig. 6). These data indicated that PPV1 infection induces apoptosis by activating the NF-κB signaling.

PPV1 infection induces apoptosis by activating the NF-κB pathway. PK-15 cells were infected with PPV1 (MOI = 1) and treated with NF-κB inhibitor BAY 11-7082 (0.5 μM or 5 μM) as indicated. Mock-infected PK-15 cells were used as controls. After 36 h, the cells were subjected to apoptosis analysis. Results are expressed as mean ± SD from three independent experiments. *0.01 < p < 0.05, **p < 0.01
Discussion
PPVs have been widely spread and become a serious threat to the swine herds. PPV1 is one of the major causes of reproductive failure in swine worldwide, and the economic loss was high in the 1960s (Streck and Truyen 2020). For the particular clinical symptoms and affected tissues/organs of PPV2 to PPV7 are not determined, the pathogenesis of novel PPVs is unclear (Vargas-Bermudez et al. 2021). Many researches have shown that PPV1 induced host cell apoptosis causing cell death, leading to sow reproductive failure (Zhang et al. 2019). The strain virulence of PPV1 is defined by the severity of sow reproductive failure. In the present study, we revealed that PPV1 induced the apoptosis of PK-15 cells by activating TLR2 and NF-κB signaling pathways (Fig. 7).

The signal pathway of apoptosis mediated by PPV1. As indicated, TLR2-mediated NF-κB signaling pathway is involved in PPV1-induced apoptosis in PK-15 cells
Apoptosis is a programmed cell death, and plays an important role in viral infection. Viruses have evolved a variety of strategies to interfere with the process of apoptosis. Some viruses are able to suppress apoptosis for viral replication and survival, while apoptosis might also facilitate viral egress (Kaminskyy and Zhivotovsky 2010). Human parvovirus B19 infection, inducing anemia and nonimmune hydrops fetalis, are the result of apoptosis in human erythroid progenitor cells and fetal tissues (Chen and Qiu 2010). Cell apoptosis contributes to the tissue damage of canine parvovirus infection in dogs (Bauder et al. 2000). Thus, apoptosis is one of the factors that cause diseases during parvovirus infection. Our results also demonstrated PPV1 induces apoptosis in PK-15 cells. The ratio of apoptotic cells reached 50% in PPV1 YL strain-infected PK-15 cells (Zhang et al. 2015). However, Meszaros et al. showed the number of apoptotic cells induced by PPV1 strain Kresse was less than 14%, and necrosis was the main form of cell death PPV1 infection induced necrosis or apoptosis depending on the viral strain and cell type (Meszaros et al. 2017b). And the components of PPV1 that induce apoptosis should be further studied.
PPV1 induces apoptosis by multiple pathways. Zhang et al. demonstrated PPV1 infection induces apoptosis by activation of p53 and mitochondria-mediated pathway (Zhang et al. 2015). Zhao and Zhang suggested PPV1 induces apoptosis via the ROS/mitochondrial pathway (Zhang et al. 2019; Zhao et al. 2016). Data showed factors or signaling pathways activated by inflammation are involved in the regulation of cell apoptosis, and apoptosis may be the result of inflammation (Yang et al. 2015). Our previous research indicated that PPV1 infection activates inflammatory cytokine production, and causes severe inflammation (Zhou et al. 2017). TLRs play a critical role in immune response. Our data showed the mRNA expression of TLR2 was increased after PPV infection, suggested TLR2 was involved in the process. Ma et al. proved TLR4 signaling pathway was involved in PPV1 infection-induced apoptosis (Ma et al. 2020). We speculated TLR2 and TLR4 might play synergistic roles in PPV1 infection.
Specific PRRs may activate apoptotic by NF-κB signaling pathways. TLR2 is a major TLRs that involves in the mediation of NF-κB signaling. NF-κB, one of the most important molecules, is essential for inflammatory responses. In our study, NF-κB luciferase reporter assay showed that higher infective doses of PPV1 caused greater activation of the NF-κB promoter. However, infection with UV-inactivated PPV1 had no impact on NF-κB promoter activity, indicating that the PPV1-mediated activation of NF-κB activity is dependent on viral replication in a dose-dependent manner. These results are consistent with the findings of Cao et al. (2017), which suggested that PPV1 infection enhanced expression of NF-κB-regulated genes in a viral dose-dependent manner, as compared with mock-infected cells. siRNA targeting TLR2 decreased the expression clearly suggested TLR2 involved in NF-κB signaling pathway during PPV1 infection.
NF-κB family consists of homodimers/heterodimers of five subunits, and p65 is the main subunit of the canonical pathway of NF-κB signaling that is potentially targeted by viruses (Rahman and McFadden 2011). Researches showed viruses induce phosphorylation of p65 at serine 276 and serine 536, and the phosphorylation status of p65 affects the transcriptional activity of NF-κB (Liu et al. 2007). Our results showed p65 localized to nucleus after PPV1 infection, which suggested that p65 was involved in activating NF-κB signaling pathway. In most cell types, NF-κB dimers are retained in the cytoplasm by interaction with specific inhibitors known as IκBs. After the degradation of the IκB subunit, NF-κB is then released and translocated to the nucleus. We determined the changes of IκB during PPV1 infection. The decrease of IκBα subunits was observed at 36 hpi. Meanwhile, a decrease in IKKα expression levels was observed at 36 hpi, corresponding with the trend of IκBα degradation.
In conclusion, our study reveals that PPV1 infection activates NF-κB signaling pathway through TLR2 to induce apoptosis in PK-15 cells. These findings provide a theoretical basis for further understanding the pathogenesis of PPV1 and provide a therapeutic approach for PPV1.
Abbreviations
- DAPI:
-
4′-6-Diamidino-2-phenylindole-dihydrochloride
- FITC:
-
Fluorescein isothiocyanate
- hpi:
-
Hour post infection
- HRP:
-
Horseradish peroxidase
- IFA:
-
Immunofluorescence assay
- IκB:
-
Inhibitory kappa B
- IKK:
-
IκB kinase
- NF-κB:
-
Nuclear factor-kappa B
- PAMPs:
-
Pathogen-associated molecular patterns
- PI:
-
Propidium iodide
- p-p65:
-
Phosphorylation of p65
- PPV:
-
Porcine parvovirus
- PRRs:
-
Pattern-recognition receptors
- PVDF:
-
Polyvinylidene difluoride
- RNAi:
-
RNA interference
- TLRs:
-
Toll-like receptors
References
Bauder B, Suchy A, Gabler C, Weissenböck H (2000) Apoptosis in feline panleukopenia and canine parvovirus enteritis. J Vet Med Series B 47:775–784. https://doi.org/10.1111/j.1439-0450.2000.00411.x
Cao L, Chen J, Wei Y, Shi H, Zhang X, Yuan J, Shi D, Liu J, Zhu X, Wang X, Cui S, Feng L (2017) Porcine parvovirus induces activation of NF-κB signaling pathways in PK-15 cells mediated by toll-like receptors. Mol Immunol 85:248–255. https://doi.org/10.1016/j.molimm.2016.12.002
Chen AY, Qiu J (2010) Parvovirus infection-induced cell death and cell cycle arrest. Future Virol 5:731–743. https://doi.org/10.1016/j.molimm.2016.12.002
Davies DA, Adlimoghaddam A, Albensi BC (2021) The effect of COVID-19 on NF-κB and neurological manifestations of disease. Mol Neurobiol 58(8):4178–4187. https://doi.org/10.1007/s12035-021-02438-2
Gray P, Dunne A, Brikos C, Jefferies CA, Doyle SL, O’Neill LA (2006) MyD88 adapter-like (Mal) is phosphorylated by Bruton’s tyrosine kinase during TLR2 and TLR4 signal transduction. J Biol Chem 281:10489–10495. https://doi.org/10.1074/jbc.M508892200
Jin X, Yuan Y, Zhang C, Zhou Y, Song Y, Wei Z, Zhang G (2020) Porcine parvovirus nonstructural protein NS1 activates NF-kappaB and it involves TLR2 signaling pathway. J Vet Sci 21:e50. https://doi.org/10.4142/jvs.2020.21.e50
Kaminskyy V, Zhivotovsky B (2010) To kill or be killed: how viruses interact with the cell death machinery. J Intern Med 267:473–482. https://doi.org/10.1111/j.1365-2796.2010.02222.x
Li J, Xiao Y, Qiu M, Li X, Li S, Lin H, Li X, Zhu J, Chen N (2021) A systematic investigation unveils high coinfection status of porcine parvovirus types 1 through 7 in China from 2016 to 2020. Microbiol Spectr 9(3):e0129421. https://doi.org/10.1128/Spectrum.01294-21
Liu P, Jamaluddin M, Li K, Garofalo RP, Casola A, Brasier AR (2007) Retinoic acid-inducible gene I mediates early antiviral response and toll-like receptor 3 expression in respiratory syncytial virus-infected airway epithelial cells. J Virol 81:1401–1411. https://doi.org/10.1128/JVI.01740-06
Ma X, Guo Z, Zhang Z, Li X, Liu Y, Zhao L, Wang X (2020) Ferulic acid protects against porcine parvovirus infection-induced apoptosis by suppressing the nuclear factor-κB inflammasome axis and toll-like receptor 4 via nonstructural protein 1. Evid Based Complement Alternat Med 2020:3943672. https://doi.org/10.1155/2020/3943672
Meszaros I, Olasz F, Csagola A, Tijssen P, Zadori Z (2017a) Biology of porcine parvovirus (ungulate parvovirus 1). Viruses 9(12):393. https://doi.org/10.3390/v9120393
Meszaros I, Toth R, Olasz F, Tijssen P, Zadori Z (2017b) The sat protein of porcine parvovirus accelerates viral spreading through induction of irreversible endoplasmic reticulum stress. J Virol 91(16):e00627-e717. https://doi.org/10.1128/JVI.00627-17
Moser BA, Escalante-Buendia Y, Steinhardt RC, Rosenberger MG, Cassaidy BJ, Naorem N, Chon AC, Nguyen MH, Tran NT, Esser-Kahn AP (2020) Small molecule NF-κB inhibitors as immune potentiators for enhancement of vaccine adjuvants. Front Immunol 11:511513. https://doi.org/10.3389/fimmu.2020.511513
Rahman MM, McFadden G (2011) Modulation of NF-κB signalling by microbial pathogens. Nat Rev Microbiol 9:291–306. https://doi.org/10.1038/nrmicro2539
Streck AF, Truyen U (2020) Porcine parvovirus. Curr Issues Mol Biol 37:33–46. https://doi.org/10.21775/cimb.037.033
Taniguchi K, Karin M (2018) NF-κB, inflammation, immunity and cancer: coming of age. Nat Rev Immunol 18:309–324. https://doi.org/10.1038/nri.2017.142
Vandevenne P, Sadzot-Delvaux C, Piette J (2010) Innate immune response and viral interference strategies developed by human herpesviruses. Biochem Pharmacol 80:1955–1972. https://doi.org/10.1016/j.bcp.2010.07.001
Vargas-Bermudez DS, Rendon-Marin S, Ruiz-Saenz J, Mogollón D, Jaime J (2021) The first report of porcine parvovirus 7 (PPV7) in Colombia demonstrates the presence of variants associated with modifications at the level of the VP2-capsid protein. PLoS ONE 16(12):e0258311. https://doi.org/10.1371/journal.pone.0258311
Yang Y, Jiang G, Zhang P, Fan J (2015) Programmed cell death and its role in inflammation. Mil Med Res 2:12. https://doi.org/10.1186/s40779-015-0039-0
Zhang H, Huang Y, Du Q, Luo X, Zhang L, Zhao X, Tong D (2015) Porcine parvovirus infection induces apoptosis in PK-15 cells through activation of p53 and mitochondria-mediated pathway. Biochem Biophys Res Commun 456:649–655. https://doi.org/10.1016/j.bbrc.2014.12.011
Zhang J, Fan J, Li Y, Liang S, Huo S, Wang X, Zuo Y, Cui D, Li W, Zhong Z, Zhong F (2019) Porcine parvovirus infection causes pig placenta tissue damage involving nonstructural protein 1 (NS1)-induced intrinsic ROS/mitochondria-mediated apoptosis. Viruses 11(4):389. https://doi.org/10.3390/v11040389
Zhao J, He S, Minassian A, Li J, Feng P (2015) Recent advances on viral manipulation of NF-κB signaling pathway. Curr Opin Virol 15:103–111. https://doi.org/10.1016/j.coviro.2015.08.013
Zhao X, Xiang H, Bai X, Fei N, Huang Y, Song X, Zhang H, Zhang L, Tong D (2016) Porcine parvovirus infection activates mitochondria-mediated apoptotic signaling pathway by inducing ROS accumulation. Virol J 13:26. https://doi.org/10.1186/s12985-016-0480-z
Zhou Y, Jin X, Jing Y, Song Y, He X, Zheng L, Wang Y, Wei Z, Zhang G (2017) Porcine parvovirus infection activates inflammatory cytokine production through toll-like receptor 9 and NF-κB signaling pathways in porcine kidney cells. Vet Microbiol 207:56–62. https://doi.org/10.1016/j.vetmic.2017.05.030
Funding
This work was supported by the National Natural Sciences Foundation of China (31772722, U1404326); the Program for Science & Technology Innovation Talents in Universities of Henan Province (15HASTIT028), and the Program for Key Scientific Research in Universities of Henan Province (18A230001).
Author information
Authors and Affiliations
Contributions
Lu Xia and Linqing Wang designed and funded the study; Menglong Xu and Xiaohui Jin performed the experiments; Menglong Xu and Chi Zhang analyzed the results and drafted the manuscript; Hang Liao, Pingli Wang, Yong Zhou and Yue Song participated in correcting the manuscript. All the authors have reviewed the final version of the manuscript and approve it for publication.
Corresponding authors
Ethics declarations
Conflict of interest
The authors have no relevant financial or non-financial interests to disclose.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Menglong Xu and Xiaohui Jin contributed equally to this work and shared first authorship.
Rights and permissions
About this article

Cite this article
Xu, M., Jin, X., Zhang, C. et al. TLR2-mediated NF-κB signaling pathway is involved in PPV1-induced apoptosis in PK-15 cells. Vet Res Commun 47, 397–407 (2023). https://doi.org/10.1007/s11259-022-09954-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11259-022-09954-z