
Abstract
Ecological restoration aims at supporting biodiversity and ecosystem services, and urban greening is a great opportunity to achieve this goal. This is facilitated by species-rich seed mixtures based on local provenances, which are designed for certain nutrient and moisture regimes based on functional plant traits. Such grassland mixtures might be cultivated on crushed waste bricks, which would be a new component of water-holding urban substrates. Thus, we studied the effects of brick quantity and quality, acid pre-treatment of bricks, soil type and moisture on biomass of designed seed mixtures. Three greenhouse experiments were conducted, with substrates consisting of different brick ratios (5% vs. 30%), brick types (clean production waste vs. demolition material), and brick treatments (acid vs. control) tested on three trait-based mixtures and a non-regional commercial standard mixture. The trait-based mixtures included information on specific leaf area, seed mass and grass-to-legume ratio. There were no negative effects of demolition bricks, soil texture and moisture on grassland biomass. Acid-treated clean porous bricks improved biomass production of the standard and intermediate mixtures, while the effect was minimal with demolition bricks. Designed seed mixtures had a biomass similar to the standard mixture under dry conditions but did not benefit from high moisture like the standard mixture. In conclusion, waste bricks are a useful additive for urban restoration substrates to save raw material, and specifically designed regional mixtures can replace commercial grassland types on these substrates.
Similar content being viewed by others
Introduction
Re-using organic or mineral wastes as planting substrate is a promising way of recycling. This has been shown for construction waste, bricks, coal gangue, paper ash pellets and sewage sludge (Du et al. 2020; Liu et al. 2019; Molineux et al. 2015; Naeth and Wilkinson 2014). In China, 2,400 million tons of construction waste were produced in 2015 (Liu et al. 2019), and in Germany 215 million tons in 2016 (Kreislaufwirtschaft 2019). Waste materials have some potential to substitute soil in restoration practice, and thus to reduce the consumption of raw material, and to save costs (Walsh et al. 2018). In this study, we focus on crushed waste bricks which are available in large quantities, for example about 10 million tons in Germany in the 2014 (Umweltbundesamt 2019). Waste accumulates during brick production and through demolition of buildings.
Brick waste as a component of restoration substrates would be a valuable alternative to using waste bricks as bulk material for construction work, while demolition bricks with remains of mortar and plaster are rarely used as substrate component (Umweltbundesamt 2019). Bricks have a high water-holding capacity, while the pH of about 10 is rather high, increasing the pH of the substrate to 7–9 (Hitchmough et al. 2001; Molineux et al. 2009). A treatment with phosphoric acid might reduce the pH value and improve soil fertility. Thus, crushed clean bricks of up to 80% are used for green roofs (Molineux et al. 2015; Stovin et al. 2015), or in small quantities as planting substrate in gardens and landscaping. However, brick-augmented substrates with varying water holding capacity and different nutrient concentrations have not been tested for grassland restoration in comparison to standard topsoil (Molineux et al. 2009; Nagase and Dunnett 2010).
Crushed waste bricks as component of planting substrates can be used for landfill and quarry restoration as well as for road verges and urban greening, often resulting in ‘novel ecosystems’ (Hobbs et al. 2006; Kowarik 2011), while they might benefit biodiversity and ecosystem services (Bowman et al. 2017). Urban greening could accommodate considerable amounts of brick-augmented substrate to support semi-natural grasslands that otherwise have declined, for example in Central Europe (Poschlod et al. 2005; Wesche et al. 2012). So far, grass-dominated seed mixtures are used for urban greening that result in species-poor swards with few resources for pollinators (Hefter et al. 2010). Most seeds are of non-regional origin, i.e. they do not derive from local provenances. Therefore, these seed mixtures are less supportive for native biodiversity (Durka et al. 2017), while seeds for restoration should be regionally adapted to benefit long-term restoration success (Bucharova et al. 2019).
A topical issue in restoration ecology is reassembling plant communities that simultaneously foster biodiversity and specific ecosystem processes (de Bello et al. 2010; Funk et al. 2008). Biodiversity could benefit from species pools of semi-natural grasslands, with a high proportion of regional provenance herbs. However, for urban greening no specific seed mixtures have been tested that would fit brick-augmented substrates. Development of such mixtures could be assisted by recent progress in trait-based community ecology (Suding et al. 2008) that promotes functional restoration (Laughlin 2014).
The classic approach of this method is to adopt a candidate list of species deemed suitable by expert opinion. However, this approach is not general and therefore not transferable and comparable to less familiar conditions. In that respect, a suitable quantitative approach could be trait-based models sensu Laughlin (2014). Trait-based species selection can be transferred to new conditions, and outcomes can be more easily compared (Suding et al. 2008). However, unspecific regional seed mixtures suffer from environmental sorting which reduces seeding efficiency (Freitag et al. 2021), while using designed seed mixtures could reduce mortality risks of seedlings, leading to lower costs, higher functional outcomes and improved restoration success (Laughlin et al. 2017). More specifically, seed mixtures for brick-augmented substrates must be adapted to high pH and variable soil moisture. For example, species with a high specific leaf area (SLA) have high transpiration and high growth rates (Poorter et al. 2009). However, the response of SLA to environmental factors must be monitored, as it tends to be higher on nitrogen-rich soils and with increased moisture (Ordoñez et al. 2009; Poorter et al. 2009). Another promising functional trait is seed mass which tends to be higher under adverse site conditions (Westoby et al. 2002). On productive soils, graminoids are a major component (Feßel et al. 2016), whereas legumes are facilitative for non-legume plants (Erktan et al. 2018).
The aim of this study is to test the effects of brick-augmented substrates on contrasting seed mixtures under different moisture regimes. We established three full-factorial experiments to address the following questions:
-
1.
How do grassland seed mixtures respond to substrates with different brick quantity and quality, and which effect have crushed bricks pre-treated with phosphorous acid?
-
2.
Does a high brick ratio reduce negative drought effects on grassland biomass?
-
3.
Do seed mixtures respond differently to soil moisture regimes on brick-based substrates?
Material and methods
Species and trait selection and community design
As test communities for assessing the quality of different brick-augmented substrates, we used lowland meadows growing on nutrient-rich moderately moist soil in Central Europe (Leuschner and Ellenberg 2018; Oberdorfer and Müller 1983). For the design of the seed mixtures, we excluded species with a rooting depth >1 m (Kutschera and Lichtenegger 1982, 1992; Landolt and Bäumler 2010) and those that were not commercially available. From the remaining pool of 41 herbaceous species, twelve forbs, three legumes, and seven grasses were selected (Appendix, Table A1; the composition of the 20 species was randomly chosen for each plot.
We constrained the communities to certain forb-grass-legume ratios, to two traits (SLA, seed mass) and the Ellenberg R value. Three opposing seed mixture types were designed based on the pre-determined species compositions by adjusting the weight proportion of the seeded species. For the ‘vigorous’ type, we opted for high SLA, low seed mass, high grass ratio and low legume ratio (Table 1), the ‘robust’ type had opposing values, and the ‘intermediate’ type was in between. In addition, the mean Ellenberg R value was set to 7 for all mixtures to account for the relatively high pH of brick-augmented substrates. The used community-weighted means of the intermediate mixture were based on communities described by Oberdorfer and Müller (1983), as also seen in commercial mixtures of the company Krimmer (Appendix, Table A2).
To calculate the proportion of each species on the entire seed mass, the community-weighted means and the randomly selected species compositions were put into the function ‘Select’ of the package 'Select' (Laughlin 2014). Trays were seeded with a density of 4 g m−2 as recommended by Kirmer (2019). These designed seed mixtures were tested against a commercial standard mixture for non-agricultural grassland with 17 species and a density of 20 g m−2 (Regel-Saatgut-Mischung RSM 7.1.2 ‘landscape lawn with forbs’ DIN 18917 2018). The R indicator values of plant species were taken from Ellenberg et al. (2001); the traits were available for 98–99% of the species from the TRY database (Kattge et al. 2020).
Experimental design
Brick-augmented substrates were tested in three experiments at the Greenhouse Laboratory Centre Dürnast, Technical University of Munich (WGS 84 (lat, lon): 48.40583, 11.69151). All treatment combinations were grown in plastic trays (50 cm × 30 cm × 6 cm). The loamy soil was provided by the company Wurzer Umwelt, and the brick rubble was production waste from modern, porous bricks (38% intra-particle pore space) of the company Leipfinger-Bader, while the demolition bricks produced in the 1960s have a lower rate of porosity (27% intra-particle pore space). The porosity of modern bricks is increased by adding sawdust to the raw material, which then burns in the kiln, leaving air voids. The demolition bricks were cleaned and sorted but still had remains of mortar and plaster, and all bricks were crushed to a fraction 4–16 mm.
The substrates with 5% bricks had pH values of 7.4 ± 0.1, and the ones with 30% pH 7.1 ± 0.5 (Table 2). The organic matter was reduced by bricks amounting in Experiments 1 and 2 to 5.4–8.4% and 0.9–3.9% in Experiment 3 (Table 2). Phosphate was significantly increased by acid-treated demolition bricks (134–548 mg 100 g−1), but not by acid-treated clean bricks (15 mg 100 g−1) (Table 2).
Experiments 1 and 2 started at the end of May 2019 and ran for 14 weeks in a semi-open greenhouse with a glass roof but with wire mesh walls so that climatic conditions were similar to outside conditions, with an average temperature of 19 ± 6 °C (www.dwd.de, accessed 27.02.2020; Appendix, Fig. A4). Experiment 3 started in mid-January 2020 and lasted 13 weeks in a heated closed greenhouse with a temperature of 20–22 °C during the day and a light supply of 70 μmol m−2 for 12 h d−1 (Appendix, Fig. A4). In Experiments 1 and 2, the trays were watered every day for the first four weeks from above, and after plant establishment they were watered from beneath. To induce a moisture gradient, water amount was the same per watering, but frequencies differed between blocks: every second day, daily, twice a day, or thrice a day. For Experiment 3, plants were watered on demand (every 5–8 days) to avoid water stress. In all experiments, non-seeded species were identified every second week and removed.
In total, 128 trays on eight tables were used in Experiment 1, and 64 in Experiment 2. Both experiments were full-factorial and had a split-plot design with a randomized complete block design on the plot level; plots were re-randomized after seven weeks. On the block level (= table), there was the treatment ‘moisture’ with two replicates, and on the plot level (= tray), Experiment 1 had three treatments with eight replicates. The treatments were four seed mixtures (standard, robust, intermediate, vigorous), two brick ratios of the substrate by volume (5%, 30%), and with or without acid treatment of the bricks with phosphoric acid (concentration of acid 0.3 mol kg−1, added in concrete mixer with a retention time of 4 min); the bricks were clean production waste. In Experiment 2, the treatments on plot level were two seed mixtures (robust, vigorous), two brick ratios (5%, 30%), and two brick types (clean production waste, demolition bricks with plaster and mortar); all bricks were treated with phosphoric acid.
Experiment 3 had a full-factorial completely randomized design and was re-randomized three times with in total 72 trays. We established four treatments with three replicates: three soil textures (sandy, medium, loamy), two brick ratios (5%, 30%), and two substrate densities (low, high), and with or without pelletized activated carbon (1 t ha−1). For the soil treatment, loam was mixed with 20, 50 or 80% quartz sand 0/4 (Table 2; Appendix Table A3).
Establishment of species ranged from 0 to 100% and was 73 ± 8% for species of the standard seed mixture, and 53–54 ± 5% for the designed regional seed mixtures in Experiments 1 and 2, and 71 ± 5% in Experiment 3 (Appendix Table A1). The species Onobrychis viciifolia emerged but could not establish and died.
Measurements and data analysis
The substrates were analyzed for grain size distribution, pH, nutrients and organic ratio (Table 2; Appendix Figs. A1, A2 and A3). In all experiments, grassland biomass was collected as a fitness indicator (Younginger et al. 2017) cut at 1 cm aboveground, while the fringe with a distance of 4 cm to the plastic edge was skipped to avoid edge effects. Then, the biomass was dried at 65 °C for three days and immediately weighed.
Due to right-skewed data distribution, biomass values were log-transformed. For Experiments 1 and 2, linear mixed-effects models were calculated with the random effect ‘block’ (= table) and the maximum likelihood method. For Experiment 3, a linear model was calculated. If the results were ‘statistically clear’ p < 0.05 (sensu Dushoff et al. 2019), we calculated contrasts with multiple comparisons and corrected them with the Tukey method. Uncertainties of the effects were expressed as standard error of the mean (SE).
The analyses were performed in R, version 4.0.2 (Team 2020) and with the packages ‘Select’ for community calculations (Laughlin et al. 2018), ‘lme4’ for liner mixed-effects models (Bates et al. 2015), ‘MuMIn’ for Pseudo-R2 values (Barton 2019), ‘DHARMa’ for model evaluation (Hartig 2020), ‘emmeans’ for calculating contrasts (Lenth 2020), and ‘ggeffects’ (function ‘ggemmeans’) to extract coefficients for the graphs (Breheny and Burchett 2017; Lüdecke 2018).
Results
Experiment 1 showed that two seed mixtures had increased biomass on substrate with acid-treated crushed bricks (interaction: χ2 (3) = 12.2, p = 6.9e–03; Fig. 1), i.e. the standard mixture (+39 ± 13%, compared to 5% bricks and no acid; t1,155 = –3.6, padj = 2.2e–03) and the intermediate mixture (+28 ± 12%, t1,155 = –2.7, padj = 3.8e–02). Furthermore, the vigorous and the robust mixtures showed an effect of <5%. Seed mixtures were differently affected by moisture regime (interaction: χ2 (9) = 25.7, p = 2.3e–03; Fig. 5): the designed mixtures performed worse under moist conditions than the standard seed mixture (standard vs. intermediate: –29 ± 6%, t1,155 = 3.8, padj = 1.4e–03), but not under dry conditions (–1 ± 9%, t1,155 = 0.1, padj = 9.9e–01). Under all moisture regimes, the designed seed mixtures were not clearly different from the standard mixture under dry conditions: for example, robust seed mixture under medium moist conditions (–30 ± 16%, t1,23.4 < –1.6, padj (Dunnett) = 6.5e–01). Furthermore, the biomass of the designed mixes did not differ statistically clear under the different moisture regimes (all t1,155 <|1.8|, padj > 2.6e–01). Pseudo-R2 values of the model for Experiment 1 were R2marginal = 0.48, R2conditional = 0.71.

Effects of acid treatment and brick ratio on biomass of four seed mixtures (cf. Table 1). For example, the effect within the standard mixture (5% control vs. 30% acid) +39 ± 13% SE, and within the intermediate mixture (5% control vs. 30% acid) +28 ± 12% SE (linear mixed-effects model: interaction: p = 6.9e–03). Shown are the estimated marginal means and the corresponding confidence intervals 95%. Letters indicate differences with p < 0.05 within each seed mixture. Data from Experiment 1 with new clean bricks
Experiment 2 revealed that demolition bricks with a ratio of 30% had no effect on biomass (χ2 (1) = 0.6, p = 4.4e–01, Fig. 2). Though, clean bricks reduced biomass by 7 ± 6% and demolition bricks by 25 ± 4% compared to the group with 5% clean bricks. Under any moisture regime, brick addition did not have a statistically clear influence on biomass production (interaction: χ2 (3) = 7.1, p = 6.8e–02; Fig. 3): the brick effect reached from –8 ± 6% (dry) to +13 ± 7% (moist). Pseudo-R2 values of the model for Experiment 2: R2marginal, = 0.28, R2conditional, = 0.65.

No statistically clear effect of brick type and brick ratio (linear mixed-effects model: interaction: p = 4.4e–01). The effect of brick type on biomass (5% clean bricks vs. 30%), i.e. clean bricks 7 ± 6% SE, and demolition bricks −25 ± 4% SE. Shown are the estimated marginal means and the corresponding confidence intervals 95%. Data from Experiment 2 with the seed mixtures ‘vigorous’ and ‘robust’; all bricks were treated with acid

The effect of brick ratio on biomass under different moisture regimes was not statistically clear (linear mixed-effects model: interaction: p = 6.8e–02). The effect ranged from −8 ± 6% SE (dry) to +13 ± 7% SE (moist) Shown are the estimated marginal means and the corresponding confidence intervals 95%; data from Experiment 1 with new clean bricks
Experiment 3 showed that brick ratio had under any soil texture no effect on biomass: (F2,59 = 0.1, p = 8.6e–01; Fig. 4; R2adj = 0.41): for loamy substrate +23 ± 20% (t1,59 = 1.3, p = 2.1e–01) and for sandy substrate +8 ± 18% (t1,59 = 0.4, p = 6.6e–01) (Fig. 5).

No effects of soil type and brick ratio on biomass of the intermediate seed mixture (linear model: interaction: p = 8.6e–01). The effect ranged from +8 ± 18% SE (sand) to plus +23 ± 20% SE (loam). Shown are the estimated marginal means and the corresponding confidence intervals 95%. Data from Experiment 3 where the seed mixture ‘intermediate’ and the brick type ‘demolition’ were compared

Effect of soil moisture on biomass of the four seed mixtures (cf. Table 1), i.e. dry (standard vs. intermediate mixture) 1 ± 9% SE, and moist −29 ± 6% SE (linear mixed-effects model: interaction: p = 2.3e–03). Shown are the estimated marginal means and the corresponding confidence intervals 95%. Different letters indicate differences with p < 0.05 within each moisture regime. Data from Experiment 1 with new clean bricks
Discussion
Acid-treated clean porous crushed bricks had a positive effect on the standard and the intermediate seed mixture, and no negative effect on the other two mixtures. There was no longer a positive effect if demolition bricks were used but also no negative effect. A brick ratio of up to 30% did not reduce the drought effect by sandy soils or few watering. The old, less porous bricks had no negative effect on biomass production. The trait-based seed mixtures were similar productive compared to standard seed mixtures under dry conditions. There was no difference in biomass among the contrastingly designed mixtures.
Effects of brick ratio, brick type and acidic pre-treatment on vegetation
To our knowledge, this is the first study that systematically tested an acid treatment of crushed bricks for planting substrates. We expected a positive effect of this treatment by lowering the pH level and by increasing phosphorus concentration. Although, the pH level was not affected significantly by acid in Experiment 1, and phosphorus was increased especially in substrates with demolition bricks in Experiment 2 (Table 2). This higher phosphorus content of demolition bricks may be the result of the reaction of phosphorus acid with the adherent mortar producing a higher amount of available Calciumphosphate compounds. In any substrate, plant-available phosphorus was present in sufficient quantities of more than 40 mg per 100 g soil (Table 2). Moreover, the substrate which had a positive effect on biomass production in Experiment 1 (acid-treated clean bricks), had a slightly increased amount of phosphate compared to the substrates with acid-treated demolition bricks which had no strong positive effect on biomass production (Table 2).
Nevertheless, we could observe in Experiment 1 a statistically clear acid effect on biomass for substrates with clean bricks and the seed mixtures intermediate (+28%) and standard (+39%). However, this effect could not be reproduced in Experiment 3, with acid-treated demolition bricks especially when added to sandy soil. The main differences were an even lower moisture than the dry conditions in Experiment 1 with nutrient-poor soils. Experiment 2 showed a significant negative effect of demolition bricks compared to 5% clean bricks. This fits to the results of Hitchmough et al. (2001), who found a negative effect of brick-augmented substrate (50% demolition brick ratio) on biomass of seedlings of calcareous grassland species (−80–90%). The differences of demolition brick substrate to clean brick substrate were the pH value (6.2 vs. 7.7), the phosphate amount (600 vs. 67 mg 100 g−1), and the amount of potassium oxide (14 vs. 40 mg 100 g−1). However, in Experiment 3, demolition bricks had no longer a negative effect.
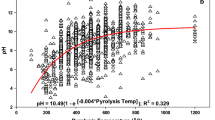
The pH value of bricks used in our experiments (clean: 8.2, demolition: 6.9) was lower than in other studies (Hitchmough et al. 2001; Molineux et al. 2009). Adding the non-acid treated bricks to soil resulted in pH values of the substrates of 7.5, which is similar to the substrate with only 5% bricks (pH 7.4). The pH can be reduced to around 8 by adding 15–20% compost (Ondoño et al. 2016), while adding 25% compost did not further change the pH value (Molineux et al. 2009). Hitchmough et al. (2001) could not reduce the pH value by adding 50% sand with a pH of 6.8 to demolition bricks. We observed a maximum reduction of the pH value by 0.4 after adding 30% demolition bricks to sandy soil. The sandy substrate still contained 14% topsoil and 0.9% organic matter in contrast to the substrate of Hitchmough et al. (2001). This suggests that organic material can buffer the pH of bricks very efficiently.
Do bricks reduce drought effects?
Crushed bricks should improve substrates by their high water-holding capacity (UBA 2018). High water holding capacity of the substrate is important for vegetation during drought periods (Farrell et al. 2012; Molineux et al. 2015). For brick-augmented roof substrates, water holding capacity seems to be more important than pH differences for species establishment (Molineux et al. 2015).
Our results showed no effect on biomass after brick addition under dry conditions in Experiment 1, which suggests no significant difference in water holding capacity between the used loamy soil and bricks. The available water capacity is affected by soil type, storage density and humus amount (Bodner et al. 2015). Therefore, we tested soil types from loam to sand under dry conditions in Experiment 3, and found that there is no difference with or without brick addition. This would mean that bricks can substitute different soil types without a significant decrease in vegetation biomass. For drought resistance, the humus ratio seems to be more important (Graceson et al. 2014), and that humus is mixed in and not lying on top of the substrate (Schröder and Kiehl 2021). An advantage of demolition bricks is that they could emaciate humid soil and keeps water holding capacity compared to an emaciation by gravel. Nevertheless, the water holding capacity of the substrate results from the available water capacity and the substrate depth (Bodner et al. 2015). Further experiments should be conducted in the field with deeper substrates.
Effect of moisture on designed seed mixtures
In ecological restoration, seed mixtures are wanted that simultaneously improve biodiversity and ecosystem services (Bowman et al. 2017). Here, a trait-based approach allows greater generality for species compositions and more predictive power (Shipley et al. 2016). Designing trait-based seed mixtures could make seeding more efficient since general seed mixtures are filtered by environmental conditions like soil fertility. To our knowledge, these are the first results on a variety of species compositions and more than ten species to test trait-based models for restoration seed mixtures (cf. Laughlin et al. 2017; Yannelli et al. 2018). This shows that it is possible to establish species-rich seed mixtures that are resilient to dry or moist conditions. However, we could not create different outcomes under varying moisture regimes for the different seed mixtures in Experiment 1. The reason could be that the designed differences in community-weighted means were too low. Finding appropriate traits and mean trait values is a key challenge for trait-based restoration (Laughlin et al. 2017). In future, specified traits should be reduced and remaining traits discriminated or explicitly diversified with the function, and (long-term) field experiments would help with upscaling. First, the established vegetation did not sufficiently represent the seed mixture because some species would have emerged only after more than 13 weeks. An extension of the greenhouse experiment was not possible, because the trays were completely rooted. Second, different seed weights might not lead to the same proportions in dominance.
In comparison to the non-regional standard seed mixture, the designed seed mixtures had a similar biomass under dry conditions, although they were sown at lower density (4 g m−2 vs. 20 g m−2). Lower seed density especially of grasses improves establishment of all species also with lower competition ability in early life stages and therefore foster biodiversity (Dickson and Busby 2009). For practical amplification, it is not necessary to reach a maximum of productivity to keep management costs low, but coverage should be enough to fulfill ecosystem functions like soil erosion. In our experiments, most designed seed mixtures approached the biomass of the standard mixture under dry conditions. These findings suggest that regional seed mixtures with a lower sowing density provide a sufficient quality for urban greening.
Conclusion
The greenhouse experiments revealed that grassland communities develop on brick-augmented substrates under different soil moisture without any negative effects on biomass. The results were consistent both for a standard seed mixture and three trait-based mixtures of regional origin. Further, the results suggest that crushed bricks could be successfully added to varying soil types, and a pre-treatment of the bricks with phosphorous acid seems unnecessary for community establishment. Thus, waste bricks are a promising component of new restoration substrates. However, to check for the large-scale effect of the water holding capacity of these novel substrates, field experiments are necessary.
Availability of data and material
Bauer M, Krause M, Heizinger V & Kollmann J (2022) Data and code for Bauer et al. (2022) Urban Ecosystems (v1.1.0) [Dataset]. – Zenodo. https://doi.org/10.5281/zenodo.6477717.
References
Barton K (2019) MuMIn:multi-model inference. R package version 1.43.15. www.CRAN.R-project.org/package=MuMIn
Bates D, Mächler M, Bolker BM, Walker S (2015) Fitting linear mixed-effects models using lme4. J Stat Softw 67:1–48. https://doi.org/10.18637/jss.v067.i01
Bodner G, Nakhforoosh A, Kaul H-P (2015) Management of crop water under drought: a review. Agron Sustain Dev 35:401–442. https://doi.org/10.1007/s13593-015-0283-4
Bowman DMJS, Garnett ST, Barlow S, Bekessy SA, Bellairs SM, Bishop MJ, Bradstock RA, Jones DN, Maxwell SL, Pittock J, Toral-Granda MV, Watson JEM, Wilson T, Zander KK, Hughes L (2017) Renewal ecology: conservation for the Anthropocene. Restor Ecol 25:674–680. https://doi.org/10.1111/rec.12560
Breheny P, Burchett W (2017) Visualization of regression models using visreg. R J 9:56–71. https://doi.org/10.32614/RJ-2017-046
Bucharova A, Bossdorf O, Hölzel N, Kollmann J, Prasse R, Durka W (2019) Mix and match: regional admixture provenancing strikes a balance among different seed sourcing strategies for ecological restoration. Conserv Genet 20:7–17. https://doi.org/10.1007/s10592-018-1067-6
Bundesanstalt für Geowissenschaften und Rohstoffe, (ed) (2005) Bodenkundliche Kartieranleitung, 5th edn. Schweizerbart, Stuttgart
de Bello F, Lavorel S, Díaz S, Harrington R, Cornelissen JHC, Bardgett RD, Berg MP, Cipriotti P, Feld CK, Hering D, Martins da Silva P, Potts SG, Sandin L, Sousa JP, Storkey J, Wardle DA, Harrison PA (2010) Towards an assessment of multiple ecosystem processes and services via functional traits. Biodivers Conserv 19:2873–2893. https://doi.org/10.1007/s10531-010-9850-9
Dickson TL, Busby WH (2009) Forb species establishment increases with decreased grass seeding density and with increased forb seeding density in a Northeast Kansas, U.S.A., experimental prairie restoration. Restor Ecol 17:597–605. https://doi.org/10.1111/j.1526-100X.2008.00427.x
DIN 18917:2018-07. Vegetation technology in landscaping. Turf and seeding (18917:1973-10).
Du T, Wang D, Bai Y, Zhang Z (2020) Optimizing the formulation of coal gangue planting substrate using wastes: The sustainability of coal mine ecological restoration. Ecol Eng 143. https://doi.org/10.1016/j.ecoleng.2019.105669
Durka W, Michalski SG, Berendzen KW, Bossdorf O, Bucharova A, Hermann J-M, Hölzel N, Kollmann J (2017) Genetic differentiation within multiple common grassland plants supports seed transfer zones for ecological restoration. J Appl Ecol 54:116–126. https://doi.org/10.1111/1365-2664.12636
Dushoff J, Kain MP, Bolker BM (2019) I can see clearly now: Reinterpreting statistical significance. Methods Ecol Evol 8:12. https://doi.org/10.1111/2041-210X.13159
Ellenberg H, Weber EW, Düll R, Wirth V, Werner W (2001) Zeigerwerte von Pflanzen in Mitteleuropa, 3rd edn. Scripta Geobotanica, vol 18. Erich Goltze, Göttingen
Erktan A, McCormack ML, Roumet C (2018) Frontiers in root ecology: recent advances and future challenges. Plant Soil 424:1–9. https://doi.org/10.1007/s11104-018-3618-5
Farrell C, Mitchell RE, Szota C, Rayner JP, Williams N (2012) Green roofs for hot and dry climates: Interacting effects of plant water use, succulence and substrate. Ecol Eng 49:270–276. https://doi.org/10.1016/j.ecoleng.2012.08.036
Feßel C, Meier IC, Leuschner C (2016) Relationship between species diversity, biomass and light transmittance in temperate semi-natural grasslands: is productivity enhanced by complementary light capture? J Veg Sci 27:144–155. https://doi.org/10.1111/jvs.12326
Freitag M, Klaus VH, Bolliger R, Hamer U, Kleinebecker T, Prati D, Schäfer D, Hölzel N (2021) Restoration of plant diversity in permanent grassland by seeding: Assessing the limiting factors along land-use gradients. J Appl Ecol 58:1681–1692. https://doi.org/10.1111/1365-2664.13883
Funk JL, Cleland EE, Suding KN, Zavaleta ES (2008) Restoration through reassembly: plant traits and invasion resistance. Trends Ecol Evol 23:695–703. https://doi.org/10.1016/j.tree.2008.07.013
Graceson A, Monaghan J, Hall N, Hare M (2014) Plant growth responses to different growing media for green roofs. Ecol Eng 69:196–200. https://doi.org/10.1016/j.ecoleng.2014.03.067
Hartig F (2020) DHARMa:residual diagnostics for hierarchical (multi-level/mixed) regression models. R package version 0.2.7. www.CRAN.R-project.org/package=DHARMa
Hefter I, Jünger G, Baasch A, Tischew S (2010) Gebietseigenes Wildpflanzensaatgut in Begrünungs- und Renaturierungsvorhaben fördern: Aufbau eines Spenderflächenkatasters und Informationssystems. Naturschutz Landschaftsplanung 42:333–340
Hitchmough J, Kendle T, Paraskevopoulou AT (2001) Seedling emergence, survival and initial growth of forbs and grasses native to Britain and central/southern Europe in low productivity urban “waste” substrates. Urban Ecosyst 5:285–308. https://doi.org/10.1023/A:1025643929335
Hobbs RJ, Arico S, Aronson J, Baron JS, Bridgewater P, Cramer VA, Epstein PR, Ewel JJ, Klink CA, Lugo AE, Norton D, Ojima D, Richardson DM, Sanderson EW, Valladares F, Vila M, Zamora R, Zobel M (2006) Novel ecosystems: theoretical and management aspects of the new ecological world order. Global Ecol Biogeogr 15:1–7. https://doi.org/10.1111/j.1466-822x.2006.00212.x
Kattge J, Bönisch G, Díaz S, Lavorel S, Prentice IC, Leadley P, … Wirth C (2020) TRY plant trait database - enhanced coverage and open access. Glob Chang Biol 26:119–188. https://doi.org/10.1111/gcb.14904
Kirmer A (2019) Vegetationstechnik der Renaturierung im Offenland. In: Kollmann J, Kirmer A, Tischew S, Hölzel N, Kiehl K (eds) Renaturierungsökologie. Springer, Berlin, Heidelberg, pp 53–70
Kowarik I (2011) Novel urban ecosystems, biodiversity, and conservation. Environ Pollut 159:1974–1983. https://doi.org/10.1016/j.envpol.2011.02.022
Kreislaufwirtschaft B (2018) Mineralische bauabfälle monitoring 2016. Bericht zum Aufkommen und zum Verbleib mineralischer Bauabfälle im Jahr 2016. Berlin. https://kreislaufwirtschaft-bau.de/Download/Bericht-11.pdf. Accessed 6 Sept 2019
Kutschera L, Lichtenegger E (1982) Wurzelatlas mitteleuropäischer Grünlandpflanzen: Band 1 Monocotyledoneae. Gustav Fischer, Stuttgart, New York
Kutschera L, Lichtenegger E (1992) Wurzelatlas mitteleuropäischer Grünlandpflanzen: Band 2 Pteridophyta und Dicotyledoneae (Magnoliopsida). Teil 1 Morphologie, Anatomie, Ökologie, Verbreitung, Soziologie, Wirtschaft. Gustav Fischer, Stuttgart, Jena, New York
Landolt E, Bäumler B (2010) Flora indicativa: Ökologische Zeigerwerte und biologische Kennzeichen zur Flora der Schweiz und der Alpen. Haupt, Bern, Genève
Laughlin DC (2014) Applying trait-based models to achieve functional targets for theory-driven ecological restoration. Ecol Lett 17:771–784. https://doi.org/10.1111/ele.12288
Laughlin DC, Strahan RT, Huffman DW, Sánchez Meador AJ (2017) Using trait-based ecology to restore resilient ecosystems: historical conditions and the future of montane forests in western North America. Restor Ecol 25:S135–S146. https://doi.org/10.1111/rec.12342
Laughlin DC, Chalmandrier L, Joshi C, Renton M, Dwyer JM, Funk JL (2018) Generating species assemblages for restoration and experimentation: A new method that can simultaneously converge on average trait values and maximize functional diversity. Methods Ecol Evol 9:1764–1771. https://doi.org/10.1111/2041-210X.13023
Lenth R (2020) Emmeans:estimated marginal means, aka least-sqares means. R package version 1.4.4. www.CRAN.R-project.org/package=emmeans
Leuschner C, Ellenberg H (2018) Ecology of Central European non-forest vegetation: Coastal to alpine, natural to man-made habitats, 6th edn. Vegetation Ecology of Central Europe, vol 2. Springer, Cham
Liu X, Liu L, Leng P, Hu Z (2019) Feasible and effective reuse of municipal sludge for vegetation restoration: physiochemical characteristics and microbial diversity. Sci Rep 9:879
Lüdecke D (2018) Ggeffects: tidy data frames of marginal effects from regression models. J Open Source Softw 3:1–5. https://doi.org/10.21105/joss.00772
Molineux CJ, Fentiman CH, Gange AC (2009) Characterising alternative recycled waste materials for use as green roof growing media in the U.K. Ecol Eng 35:1507–1513. https://doi.org/10.1016/j.ecoleng.2009.06.010
Molineux CJ, Gange AC, Connop SP, Newport DJ (2015) Using recycled aggregates in green roof substrates for plant diversity. Ecol Eng 82:596–604. https://doi.org/10.1016/j.ecoleng.2015.05.036
Naeth MA, Wilkinson SR (2014) Establishment of restoration trajectories for upland tundra communities on diamond mine wastes in the Canadian Arctic. Restor Ecol 22:534–543. https://doi.org/10.1111/rec.12106
Nagase A, Dunnett N (2010) Drought tolerance in different vegetation types for extensive green roofs: effects of watering and diversity. Landsc Urban Plan 97:318–327. https://doi.org/10.1016/j.landurbplan.2010.07.005
Oberdorfer E, Müller T (eds) (1983) Süddeutsche Pflanzengesellschaften, 2nd edn. Fischer, Stuttgart, New York
Ondoño S, Martínez-Sánchez JJ, Moreno JL (2016) The composition and depth of green roof substrates affect the growth of Silene vulgaris and Lagurus ovatus species and the C and N sequestration under two irrigation conditions. J Environ Manage 166:330–340
Ordoñez JC, van Bodegom PM, Witte J-PM, Wright IJ, Reich PB, Aerts R (2009) A global study of relationships between leaf traits, climate and soil measures of nutrient fertility. Global Ecol Biogeogr 18:137–149. https://doi.org/10.1111/j.1466-8238.2008.00441.x
Poorter H, Niinemets U, Poorter L, Wright IJ, Villar R (2009) Causes and consequences of variation in leaf mass per area (LMA): a meta-analysis. New Phytol 182:565–588. https://doi.org/10.1111/j.1469-8137.2009.02830.x
Poschlod P, Bakker JP, Kahmen S (2005) Changing land use and its impact on biodiversity. Basic Appl Ecol 6:93–98. https://doi.org/10.1016/j.baae.2004.12.001
R Core Team (2020) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. www.R-project.org
Schröder R, Kiehl K (2021) Testing standard growth substrates for establishing native dry sandy grassland species on extensive green roofs in Northern Germany. Basic Appl Ecol 56:181–191. https://doi.org/10.1016/j.baae.2021.07.010
Shipley B, de Bello F, Cornelissen JHC, Laliberté E, Laughlin DC, Reich PB (2016) Reinforcing loose foundation stones in trait-based plant ecology. Oecologia 180:923–931. https://doi.org/10.1007/s00442-016-3549-x
Stovin V, Poë S, De-Ville S, Berretta C (2015) The influence of substrate and vegetation configuration on green roof hydrological performance. Ecol Eng 85:159–172. https://doi.org/10.1016/j.ecoleng.2015.09.076
Suding KN, Lavorel S, Chapin FS, Cornelissen JHC, Díaz S, Garnier E, Goldberg D, Hooper DU, Jackson ST, Navas M-L (2008) Scaling environmental change through the community-level: a trait-based response-and-effect framework for plants. Global Change Biol 14:1125–1140. https://doi.org/10.1111/j.1365-2486.2008.01557.x
Umweltbundesamt (2018) Ziegel: Factsheet. www.umweltbundesamt.de/sites/default/files/medien/3521/dokumente/factsheet_ziegel_fi.pdf. Accessed 6 Nov 2019
Walsh D, Glass K, Morris S, Zhang H, McRae I, Anderson N, Alfieri A, Egendorf SP, Holberton S, Owrang S, Cheng Z (2018) Sediment exchange to mitigate pollutant exposure in urban soil. J Environ Manag 214:354–361. https://doi.org/10.1016/j.jenvman.2018.03.013
Wesche K, Krause B, Culmsee H, Leuschner C (2012) Fifty years of change in Central European grassland vegetation: Large losses in species richness and animal-pollinated plants. Biol Conserv 150:76–85. https://doi.org/10.1016/j.biocon.2012.02.015
Westoby M, Falster DS, Moles AT, Vesk PA, Wright IJ (2002) Plant ecological strategies: some leading dimensions of variation between species. Annu Rev Ecol Syst 33:125–159. https://doi.org/10.1146/annurev.ecolsys.33.010802.150452
Yannelli FA, Karrer G, Hall R, Kollmann J, Heger T (2018) Seed density is more effective than multi-trait limiting similarity in controlling grassland resistance against plant invasions in mesocosms. Appl Veg Sci 21:411–418. https://doi.org/10.1111/avsc.12373
Younginger BS, Sirová D, Cruzan MB, Ballhorn DJ (2017) Is biomass a reliable estimate of plant fitness? Appl Plant Sci 5:1600094
Acknowledgements
We thank Joachim Schattauer and Robert Hansel and their team of the Greenhouse Laboratory Centre Dürnast for technical help. We are thankful for the help with brick preparation, experimental set up and measuring, by Simon Schneider, Holger Paetsch, Jakob Huber, Janosch Fiedler and Hakan Tolbas. We are grateful to the company Umwelt Wurzer for their extensive support of the entire project and especially for the supply of the substrates. Administrative support by Fabian Glaab und Julius Greim (RAS, Regensburg) is acknowledged. We thank two anonymous reviewers for really helpful comments. The project was funded by the Federal Ministry for Economic Affairs and Energy within the ZIM program (Zentrales Innovationsprogramm Mittelstand).
Funding
Open Access funding enabled and organized by Projekt DEAL. The project was funded by the Federal Ministry for Economic Affairs and Energy by the ZIM program (Zentrales Innovationsprogramm Mittelstand).
Author information
Authors and Affiliations
Contributions
MB and JK designed the study. All authors designed the substrates, and MB and JK the regional seed mixtures. MB set up the experiments, conducted data sampling, performed the statistical analyses, and wrote the manuscript. VH pre-treated demolition bricks, and MK developed and conducted the acid treatments. MB wrote the manuscript, and JK, MK and VH substantially contributed to various revisions.
Corresponding author
Ethics declarations
Ethics approval
We approve to comply the ethical standards.
Consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Highlights
• Grassland mixtures grow successfully on substrates augmented with crushed bricks.
• Biomass production is unaffected by brick quantity and quality.
• Performance of trait-based seed mixtures matches commercial standard mixtures under dry conditions.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Bauer, M., Krause, M., Heizinger, V. et al. Using crushed waste bricks for urban greening with contrasting grassland mixtures: no negative effects of brick-augmented substrates varying in soil type, moisture and acid pre-treatment. Urban Ecosyst 25, 1369–1378 (2022). https://doi.org/10.1007/s11252-022-01230-x
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11252-022-01230-x