
Abstract
Heat shock proteins act as molecular chaperones that have preferentially been transcribed in response to severe perturbations of the cellular homeostasis such as heat stress. Here the traits respiration rate (RR), rectal temperature (RT), pack cell volume (PCV) and the individual heat tolerance coefficient (HTC) were recorded as physiological responses on heat stress (environmental temperatures) in Bos taurus (crossbred Holstein Friesian; HF) and B. indicus (Thai native cattle: White Lamphun; WL and Mountain cattle; MT) animals (n = 47) in Thailand. Polymorphisms of the heat shock protein 90-kDa beta gene (HSP90AB1) were evaluated by comparative sequencing. Nine single nucleotide polymorphisms (SNP) were identified, i.e. three in exons 10 and 11, five in introns 8, 9, 10 and 11, and one in the 3′UTR. The exon 11 SNP g.5082C>T led to a missense mutation (alanine to valine). During the period of extreme heat (in the afternoon) RR and RT were elevated in each of the three breeds, whereas the PCV decreased. Mountain cattle and White Lamphun heifers recorded significantly better physiologic parameters (p < 0.05) in all traits considered, including or particularly HTC than Holstein Friesian heifers. The association analysis revealed that the T allele at SNP g.4338T>C within intron 3 improved the heat tolerance (p < 0.05). Allele T was exclusively found in White Lamphun animals and to 84% in Mountain cattle. Holstein Friesian heifers revealed an allele frequency of only 18%. Polymorphisms within HSP90AB1 were not causative for the physiological responses; however, we propose that they should at least be used as genetic markers to select appropriate breeds for hot climates.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Global warming and the proposed climate change are likely to become the major threats to the sustainability of livestock production systems in the future. Simulations of different climate scenarios suggest regional rises in temperature. In addition, the intensity and duration of heat waves will dramatically increase (Gaughan et al. 2010). The changes will cause pertinent heat stress to livestock visible as predicted by, e.g. a reduced feed consumption rate (Bernabucci et al. 1999), a decreased milk production (Sharma et al. 1988) and a lower reproductive success rate (Cavestany et al. 1985).
The physiological mechanisms of heat stress regulation are known to be identical in Bos taurus and B. indicus cattle. Temperatures above the comfort zone, a high relative humidity and an intensive solar radiation alone or together with a low wind speed are the primary factors that increase an individual’s heat load and finally cause (heat) stress (Mader et al. 2006). B. indicus is, however, generally better adapted to heat stress (Beatty et al. 2006): several studies report that the detrimental effects of heat stress on production traits are of a lesser extent (Gaughan et al. 1999; Gaughan et al. 2010). Mammals respond to heat stress with an evolutionary old and conserved adaptive cellular system. It is characterized by the transcriptional activation and accumulation of a set of proteins known as heat shock proteins (HSP). Isoforms of these proteins are categorized into families with respect to their molecular weight, i.e. HSP27, HSP60, HSP70, HSP90 and HSP110/104 (Kregel 2002). Proposed functions of HSPs are microfilament stabilization (HSP27), refolding of proteins and prevention of aggregation of denatured proteins (HSP60), regulation of steroid hormone receptors and protein translocation (HSP90) and protein folding (HSP110/104). HSP70 proteins are molecular chaperons. HSP27 and HSP70 proteins have antiapoptotic and HSP60 proteins proapoptotic effects (Kregel 2002).
Ninety-kilodalton heat shock proteins (Hsp90) act as important molecular chaperones that are constitutively expressed as a consequence of heat or stress induction (Chen et al. 2006). Two major cytoplasmatic Hsp90 isoforms are constituted by gene duplication: the inducible Hsp90α and the constitutive Hsp90β form. The contribution of Hsp90 isoforms to various cellular processes including signal transduction, protein folding, protein degradation, cell survival and morphological evolution has extensively been studied (Csermely et al. 1998).
The ‘trait’ heat tolerance is a quantitative trait (Gaughan et al. 2010; Li et al. 2011; Liu et al. 2011). Several studies aimed to find the link between phenotypes and genotypes. A quantitative trait locus (QTL) study in Drosophila melanogaster (Morgan and Mackay 2006) mapped heat stress resistance to a genomic region on chromosome 3 containing amongst other genes the positional candidate gene HSP83, which is the ortholog to the mammalian HSP90 gene family (Marcos-Carcavilla et al. 2010). In sheep, polymorphisms within another Hsp90 gene, the HSPAA1, were investigated. A single nucleotide polymorphism (SNP) located at position −660 in the 5′flanking region was associated with different thermal conditions (Marcos-Carcavilla et al. 2010). An SNP at nucleotide position 2789 within ATP1A1 messenger RNA is known to be associated with heat tolerance traits in dairy cows (Liu et al. 2010; Liu et al. 2011). Effects of the SNP g.1524G>A, g.3494T>C and g.6601G>A within HSP70A1A affect thermo-tolerance in Chinese Holstein cattle (Li et al. 2011). However, there have been no reports of genetic variations in bovine HSP90 genes and heat tolerance.
The objective of this study was to record physiological parameters along with heat stress, to search for sequence variants in HSP90AB1 and to describe putative associations between them in three cattle breeds used in Northern Thailand.
Materials and methods
Experimental animals
Forty-seven clinically healthy not lactating females between 12 and 18 months were randomly selected and kept at the experimental farm of the Chiang Mai University in Thailand. The animals had different genetic backgrounds and belonged to the indigenous B. indicus breeds White Lamphun (WL; n = 17) and Mountain cattle (MT; n = 16). Fourteen crossbreds (Holstein Friesian (HF) × Thai native breed) were further used. As the proportion of Holstein Friesian blood was individually between 82.8% and 98.4%, these animals are further regarded as Holstein Friesian heifers. The cattle were kept in groups according to the animal welfare rules at the experimental farm under natural conditions. They were fed ad libitum on seasonal grass, rice straw and fresh water.
Physiological parameters
Respiratory rate (diaphragm movements per minute) (RR) and rectal temperature (RT) (°C) were measured in the morning (8.00 a.m.) and in the afternoon (2.00 p.m.). In addition, blood samples were collected according to the recommendation of the manufacturer in microhematocrit capillaries to measure the pack cell volume (PCV) using a hematocrit centrifuge at 10000 rpm for 5 min (HAEMATOKRIT-210; Hettich, Germany). The measurements and the sample collection were performed 2 weeks per month for four consecutive months (September to December, i.e. from the end of the rainy season until the middle of the winter season) to achieve eight observations per animal. The outdoor temperature and the relative humidity (RH) (%) were recorded daily during the experiment.
Earlier heat tolerance experiments led to the development of a formula (Rhoad 1944) to calculate an individual’s heat tolerance coefficient (HTC). This formula, also known as the Iberia heat tolerance test for cattle, is as follows:
where HTC is the heat tolerance coefficient, ART is the average rectal temperature, 38.3 is the physiological bovine body temperature, 10 is a correction factor to convert deviations in body temperature to a unit basis and 100 is the perfect efficiency in maintaining temperature at 38.3°C. The index of HTC was calculated for each cow to assess its heat adaptability.
Molecular genetics
Genomic DNA was extracted for all experimental animals from whole blood (9-ml vials containing EDTA) and/or from small pieces of tissue taken from the ears with a modified salting out method according to Sambrook et al. (1989) and Miller et al. (1988). Twelve DNA samples (four HF, four WL and four MT) of Thai heifers and 12 samples (four German Holstein Friesians, four Holstein Reds and four Charolais) of the DNA repository at the Institute of Veterinary Medicine in Göttingen (Germany) were randomly chosen to screen for SNP by comparative sequencing.
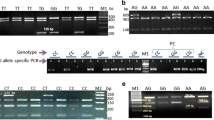
Six primer combinations were created based on the publicly available bovine HSP90AB1 gene sequence (Acc. No. NW001494158). The PCR products cover exons 2 to 12 (Table 1). PCR was carried out using 50 to 100 ng of genomic DNA, 0.2 mM dNTPs, 40 pM of each primer and 2.5 U of Taq DNA polymerase in 1× PCR buffer in a final volume of 25 μl. The PCR profile used was: 35 cycles at 94°C for 30 s, a primer specific annealing temperature (see Table 1) for 30 s and an extension period of 30 s at 72°C with an initial denaturation for 2 min at 94°C and a final extension at 72°C for 5 min. PCR reactions were performed using the Biometra T-Gradient thermocycler (Biometra, Germany). To check fragment integrity PCR products were separated on 1% agarose gels. PCR products were then purified with the QIAquick PCR Purification Kit (Qiagen, Germany). The purified PCR products were directly sequenced using the BigDye™ Terminator Cycle Sequencing Ready Reaction Kit® (Applied Biosystems, Germany) on an automated DNA sequencer (ABI-PRISM 3100® capillary analyzer; Applied Biosystems, Germany). The sequenced data were analyzed and manually checked using the software suite DNASTAR LasergeneTM 6® (DNASTAR, Inc., Germany).
Statistical analysis
Gene diversity, allele and genotype frequencies and their accordance with or deviation from the Hardy–Weinberg law were determined by POPGENE 1.31 (Yeh et al. 1999) and GenAlEx 6.3 (Peakall and Smouse 2006). For each trait, association analyses via regression on individual SNP genotypes, a repeated gene substitution MIXED model and least square means (SAS Inst., Inc., Cary, NC, USA) were performed. In the first step any possible association between an SNP within the HSP90AB1 gene and a trait was analyzed using stepwise regression analysis. In the second step a gene substitution model was used to analyze breed specific effects and those of significant SNPs driven from the stepwise regression. The following model was applied:
where Y ij is the phenotypic value of heat associated traits, μ is the overall mean, B i is the fixed effect of ith breed, b k is the regression coefficient on the number of copies of significant alleles of HSP90AB1 gene and k is the number of significant SNPs of the HSP90AB1 gene. X ij presents the copies of alleles of significant SNPs within HSP90AB1, and e ij is the random error. The sire effect was not included in the statistical model. Significance level of differences among genotype groups was determined at p < 0.05.
Results and discussion
Polymorphism screen and population genetics parameters
Nine novel polymorphisms − SNP01 to SNP09 (three in exons, five in introns and one in the 3′UTR) − covering 5,664 bp of the bovine HSP90AB1 were detected by comparative sequencing of 24 animals representing the six breeds. SNP07 led to a missense mutation (alanine to valine); the further SNPs proved to be silent. Allele and genotype frequencies are displayed in Table 2. Fixed allele frequencies were predominantly found in the HF heifers, and the most balanced distribution of alleles over all data displayed the Mountain cattle. A close to 1:1 ratio of alleles was only found for four SNPs (SNP04, SNP06 and SNP09 in Mountain cattle, and SNP08 in White Lamphun). The calculated genetic heterozygosity based on allele frequencies was low in Holstein Friesian (0.071) but high in Thai native cattle (0.326 for Mountain cattle and one of 0.307 for White Lamphun). The data support therefore a higher genetic diversity of Thai native cattle as proposed before by the Department of Livestock Development in Thailand (Boonyanuwat et al. 2005). The authors calculated mean heterozygosities between 0.415 and 0.565 after analysis of 30 STS loci in four Thai indigenous cattle (including White Lamphun).
Physiological parameters and their associations with HSP90AB1 sequence variants
Associations between sequence variants within HSP90AB1 and physiological parameters were analyzed. Earlier, others have considered RR, PCV, RT and HTC as parameters to evaluate the heat stress/tolerance of cattle (Beatty et al. 2006; Liu et al. 2010; Liu et al. 2011). We elaborated the parameters further to define the traits AM-RR (respiratory rate in the morning), PM-RR (respiratory rate in the afternoon), AM-PCV (blood pack cell volume in the morning), PM-PCV (blood pack cell volume in the afternoon), AM-RT (rectal temperature in the morning) and PM-RT (rectal temperature in the afternoon). In addition, all recorded observations to calculate average values for RR, PCV and RT (ARR, APCV and ART) were used. During the experimental time an averaged hot and humid climate of 22°C and 94% RH in the morning increased to 34°C and 68% RH in the afternoon. During the extreme heat in the afternoon, RR and RT traits were elevated in animals of all of the breeds, whereas all traits corresponding to the PCV decreased compared to the morning values. Mountain cattle and White Lamphun heifers recorded significantly better physiologic parameters (p < 0.05) in all traits considered, including or particularly HTC than Holstein Friesian heifers (98.38 and 96.85 compared to 95.28) (Table 4). Table 3 summarizes the effects of the SNP on the physiological parameters using the stepwise regression analysis. To determine which of the nine SNPs were associated with the traits, a forward stepwise regression analysis was conducted (α = 0.05 for inclusion and 0.05 exclusion). For five (SNP03, SNP06, SNP07, SNP08 and SNP09) of the nine SNPs, an effect on at least one of the ten traits was computed. SNP03 was the most frequently associated DNA variation (effects on eight of the ten traits) with as well the highest significance thresholds. Effects of miscellaneous SNP on a trait were evident, except for the RR traits.
HSP90AB1 SNP and RR traits
An increased RR is an important thermoregulatory response to heat stress. It aids in heat dissipation via evaporative cooling (Hammond et al. 1996; Beatty et al. 2006). Thus, a low RR may indicate an improved thermo-tolerance. Using the stepwise regression model, we report associations between SNP03 and SNP06 on RR traits approved by p < 0.05 or better. The possession of one T allele at SNP06 increased the AM-RR by 3.24 times/min (p < 0.0001), whereas one copy of the T allele at SNP03 lowered the PM-RR (−2.68 times/min) as well as the ARR (−3.10 times/min). The allele frequency distributions indicate a high frequency of the detrimental SNP06 T allele (0.96) in Holstein Friesian heifers but elevated frequencies of the supportive T allele at SNP03 in the indigenous Thai cattle. Contrary to that, the gene substitution model (Table 4) suggests that this observation is primarily breed-specific: White Lamphun has lower RR compared to Mountain cattle resp. Holstein Friesian for all three traits that also differ significantly (p < 0.05) except for AM-RR. For this trait no significant differences between Mountain cattle and White Lamphun do exist. In literature, there is no evidence that under physiological conditions the respiratory capacity to handle heat is superior in zebu cattle. The proportion of evaporation was roughly similar for Brahman, Holstein, Jersey and Brown Swiss. Heat stress, however, enhances the evaporative heat loss via respiration in European breeds (Seif et al. 1979; Gaughan et al. 1999; Gaughan et al. 2010) indicating more sophisticated heat loss mechanisms in less-adapted breeds to higher temperatures (Hansen 2004).
HSP90AB1 SNP and PCV traits
SNP03 resp. SNP07 were significantly (p < 0.001 resp. p < 0.05) associated with the PCV traits. In any case, the presence of T allele provided an advantage of 2.62 to 4.50% (Table 3). In turn, the gene substitution model proved that again only the breed attributed significantly to effects on PCV traits. Mountain cattle heifers revealed the highest percentage of recorded PCV, whereas Holstein Friesian heifers showed the lowest and White Lamphun cattle represented medium values. The T allele frequencies are in fact not the highest in the Mountain cattle. Thus, we assume that these two SNPs might act rather as markers than as causative sequence variations for PCV traits. Putative physiological differences of B. taurus resp. B. indicus cattle to continuous heat and humidity were investigated previously by Beatty et al. (2006). The authors propose that the increased water consumption under higher temperatures will lead to an increased total blood volume and a decrease in PCV. We did not measure the total blood volume, the water intake − and also not the water output as urine − to assure this observation but conclude that Mountain cattle heifers consume less water to keep the homeostasis compared to the other two breeds.
HSP90AB1 SNP and RT traits
Most associations between SNPs within HSP90AB1 and traits were recorded for rectal temperature. These effects were highly significant but at the same time also inconsistent as well. In total, nine putative SNP effects existed. The stepwise regression analysis revealed effects of the T allele on the trait AM-RT: SNP06 accounts for a temperature raise of 1.10°C, and a decrease of 0.99°C is at the same time caused by a T allele at SNP09. The T and C allele frequency at the loci SNP06 and SNP09 is identical in Holstein Friesian and White Lamphun but not the ones for SNPs 07 and 08 that are physically lying in between. As the SNP positions are only 428 bp away from each other, recombination events in this gene area are possible. The estimated regression coefficient decreased for the traits PM-RT resp. ART in the presence of a T allele at SNP03 by 0.28°C (p = 0.006) resp. by 0.11°C (p = 0.019). A T allele at SNP07 was associated with a temperature increase of 0.23°C (p = 0.012) resp. 0.16°C (p = 0.009). Preferred HTCs are associated with the T allele at SNP03, and a detrimental effect on this trait comes from variant T at SNP08 (Table 3). The gene substitution model (Table 4) finally proved that only RT traits depend on the breed and the investigated SNP.
Do HSP90AB1 SNPs contribute to heat stress/heat tolerance in Thai cattle breeds?
Thailand is located on the Indi–China peninsula. The climate is monsoonal tropical that remains hot and humid throughout the year. The average temperature is about 29°C, ranking in Bangkok (capital city) from 35°C in April to 17°C in December (MFA 2011). White Lamphun and the Mountain cattle are the most prominent native cattle breeds in Northern Thailand. They are rather fertile animals, tolerant towards a poor food quality and also towards internal and external parasites (Rattanaronchart 1998). The breeds are well adapted to the environment, but there were very few studies to prove this both with phenotypic and genetic data.
Here in no case, the values of the investigated physical parameters were pathological, but the data clearly underline a superior performance of Mountain cattle and White Lamphun compared to the B. taurus individuals. There are several physiological mechanisms to cope with heat stress (i.e. sweating, high respiratory rate, rising rectal temperature above critical thresholds, increased water consumption, reduced metabolic rate and a decreased dry matter intake) that at the same time reveal a negative impact on the production and reproduction performance of the cattle (West 2003; Hansen 2004; Beatty et al. 2006). It is well described that all of these physiological responses are substantially enhanced in B. taurus compared to B. indicus (Hammond et al. 1996; Collier et al. 2008). In addition, there is also ample evidence that the basal metabolic rate of B. indicus is generally lower compared to B. taurus (Gaughan et al. 1999; Hansen 2004; Gaughan et al. 2010). Clearly, low metabolic rates are consequences of reduced or low performance traits such as growth rate and milk yield. Thus, low producing cattle (livestock) reveal an increased heat tolerance (Reid et al. 1991). However, to finally cope well with heat stress further factors including housing, nutrition, health status, age and body condition have to be considered (Gaughan et al. 2010).
Conclusions
The association analysis revealed that the T allele at SNP g.4338T>C (SNP03) improved the heat tolerance (p < 0.05) of the animals. Allele T was exclusively found in White Lamphun animals and to 84% in Mountain cattle. Holstein Friesian heifers revealed an allele frequency of only 18%. The study indicates breed specific physiological responses to heat stress. Polymorphisms within HSP90AB1 were not causative for the physiological responses; however, the results propose that this gene is an attractive candidate for heat tolerance and should at least be used as a genetic marker to select appropriate breeds for hot climates.
References
Beatty, D.T., Barnes, A., Taylor, E., Pethick, D., McCarthy, M., Maloney, S.K., 2006. Physiological responses of Bos taurus and Bos indicus cattle to prolonged, continuous heat and humidity. Journal of Animal Science, 84, 972–985.
Bernabucci, U., Bani, P., Ronchi, B., Lacetera, N., Nardone, A., 1999. Influence of short- and long-term exposure to a hot environment on rumen passage rate and diet digestibility by Friesian heifers. Journal of Dairy Science, 82, 967–973.
Boonyanuwat, K., Suwanmajo, S., Khumsung, V., 2005. Genetic diversity for sustainable of Thai indigenous cattle of Department of Livestock Development. Department of Livestock Development, Accessed on 03.03.2011. http://www.dld.go.th/research-AHD/research/Webpage/2548/48(3)-0406-290.pdf.
Cavestany, D., el-Wishy, A.B., Foote, R.H., 1985. Effect of season and high environmental temperature on fertility of Holstein cattle. Journal of Dairy Science, 68, 1471–1478.
Chen, B., Zhong, D., Monteiro, A., 2006. Comparative genomics and evolution of the HSP90 family of genes across all kingdoms of organisms. BMC Genomics, 7, 156.
Collier, R.J., Collier, J.L., Rhoads, R.P., Baumgard, L.H., 2008. Invited review: genes involved in the bovine heat stress response. Journal of Dairy Science, 91, 445–454.
Csermely, P., Schnaider, T., Soti, C., Prohaszka, Z., Nardai, G., 1998. The 90-kDa molecular chaperone family: structure, function, and clinical applications. A comprehensive review. Pharmacology & Therapeutics, 79, 129–168.
Gaughan, J.B., Mader, T.L., Holt, S.M., Josey, M.J., Rowan, K.J., 1999. Heat tolerance of Boran and Tuli crossbred steers. Journal of Animal Science, 77, 2398–2405.
Gaughan, J.B., Mader, T.L., Holt, S.M., Sullivan, M.L., Hahn, G.L., 2010. Assessing the heat tolerance of 17 beef cattle genotypes. International Journal of Biometeorology, 54, 617–627.
Hammond, A.C., Olson, T.A., Chase, C.C., Jr., Bowers, E.J., Randel, R.D., Murphy, C.N., Vogt, D.W., Tewolde, A., 1996. Heat tolerance in two tropically adapted Bos taurus breeds, Senepol and Romosinuano, compared with Brahman, Angus, and Hereford cattle in Florida. Journal of Animal Science, 74, 295–303.
Hansen, P.J., 2004. Physiological and cellular adaptations of zebu cattle to thermal stress. Animal Reproduction Science, 82–83, 349–360.
Kregel, K.C., 2002. Heat shock proteins: modifying factors in physiological stress responses and acquired thermotolerance. Journal of Applied Physiology, 92, 2177–2186.
Li, Q., Han, J., Du, F., Ju, Z., Huang, J., Wang, J., Li, R., Wang, C., Zhong, J., 2011. Novel SNPs in HSP70A1A gene and the association of polymorphisms with thermo tolerance traits and tissue specific expression in Chinese Holstein cattle. Molecular Biology Report, 38, 2657–2663.
Liu, Y.X., Zhou, X., Li, D.Q., Cui, Q.W., Wang, G.L., 2010. Association of ATP1A1 gene polymorphism with heat tolerance traits in dairy cattle. Genetics and Molecular Research, 9, 891–896.
Liu, Y., Li, D., Li, H., Zhou, X., Wang, G., 2011. A novel SNP of the ATP1A1 gene is associated with heat tolerance traits in dairy cows. Molecular Biology Report, 38, 83–88.
Mader, T.L., Davis, M.S., Brown-Brandl T., 2006. Environmental factors influencing heat stress in feedlot cattle. Journal of Animal Science, 84, 712–719.
Marcos-Carcavilla, A., Mutikainen, M., Gonzalez, C., Calvo, J.H., Kantanen, J., Sanz, A., Marzanov, N.S., Perez-Guzman, M.D., Serrano, M., 2010. A SNP in the HSP90AA1 gene 5′ flanking region is associated with the adaptation to differential thermal conditions in the ovine species. Cell Stress Chaperones, 15, 67–81.
MFA, 2011. Ministry of Foreign Affairs: about Thailand. www.mfa.go.th. Accessed on 03.03.2011.
Miller, S.A., Dykes, D.D., Polesky, H.F., 1988. A simple salting out procedure for extracting DNA from human nucleated cells. Nucleic Acids Research, 16, 1215.
Morgan, T.J., Mackay, T.F., 2006. Quantitative trait loci for thermotolerance phenotypes in Drosophila melanogaster. Heredity, 96, 232–242.
Peakall, R.O.D., Smouse, P.E., 2006. GENALEX 6: genetic analysis in Excel. Population genetic software for teaching and research. Molecular Ecology Notes, 6, 288–295.
Rattanaronchart, S., 1998. White Lamphun cattle conservation and development project: a brief of 15 years experience. Department of Animal Science, Faculty of Agriculture, Chiang Mai University, Chiang Mai, Thailand.
Reid, C.R., Bailey, C.M., Judkins, M.B., 1991. Metabolizable energy for maintenance of beef-type Bos taurus and Bos indicus x Bos taurus cows in a dry, temperate climate. Journal of Animal Science, 69, 2779–2786.
Rhoad, A.O., 1944. The Iberia heat tolerance test for cattle. Tropical Agriculture, 21, 162–164.
Sambrook, J., Frisch, E.F., Maniatis, T., 1989. Molecular cloning: a laboratory manual. Cold Spring Harbor Laboratory, New York.
Seif, S.M., Johnson, H.D., Lippincott, A.C., 1979. The effects of heat exposure (31 degrees C) on Zebu and Scottish Highland cattle. International Journal of Biometeorology, 23, 9–14.
Sharma, A.K., Rodriguez, L.A., Wilcox, C.J., Collier, R.J., Bachman, K.C., Martin, F.G., 1988. Interactions of climatic factors affecting milk yield and composition. Journal of Dairy Science, 71, 819–825.
West, J.W., 2003. Effects of heat-stress on production in dairy cattle. Journal of Dairy Science, 86, 2131–2144.
Yeh, F.C., Yang, R., Bozle, T., 1999. POPGENE version 1.31, MS Windows-based freeware for population genetic analysis. University of Alberta and Center for International Forestry Research.
Acknowledgements
R. Charoensook is a fellow of the Katholischer Akademischer Ausländer-Dienst (KAAD). We gratefully acknowledge the funding support for this project provided by the University of Göttingen to C. Knorr.
Open Access
This article is distributed under the terms of the Creative Commons Attribution Noncommercial License which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This is an open access article distributed under the terms of the Creative Commons Attribution Noncommercial License (https://creativecommons.org/licenses/by-nc/2.0), which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
About this article
Cite this article
Charoensook, R., Gatphayak, K., Sharifi, A.R. et al. Polymorphisms in the bovine HSP90AB1 gene are associated with heat tolerance in Thai indigenous cattle. Trop Anim Health Prod 44, 921–928 (2012). https://doi.org/10.1007/s11250-011-9989-8
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11250-011-9989-8