
Abstract
Legume cover crops can play a valuable role in maintaining and increasing soil quality and nitrogen availability, but are infrequently grown in the Upper Midwest due to short growing seasons with minimal management windows; cold, wet springs; and harsh winters. This study was performed to assess the viability of winter annual legume species in northern climates as a potential source of nitrogen (N) fertility to a 75-day sweet corn (Zea mays convar. saccharata var. rugosa) cash crop in Lamberton and Grand Rapids, MN in 2016 and 2017. Treatments included medium red clover (Trifolium pratense), two cold-hardy ecotypes of hairy vetch (Vicia villosa Roth), a cereal rye-hairy vetch biculture (Secale cereale L., Vicia villosa Roth), cereal rye as a non-legume control, and a fallow weed-free control. Legumes were split into rhizobia inoculated and non-inoculated treatments. Inoculation had no effect on nodulation, biomass production, or N fixation likely due to competition with endogenous rhizobia strains. The rye monoculture and biculture produced the most biomass at all site-years averaging 7.7 and 7.0 Mg ha−1 respectively while the two vetch ecotypes averaged 4.5 and 3.9 Mg ha−1. Both vetch ecotypes contributed among the most nitrogen in all site-years, contributing up to 211 kg N ha−1 from aboveground biomass. Data from natural abundance isotopic approaches indicate that 75% of vetch tissue N in Grand Rapids and 59% of vetch tissue N in Lamberton was derived from atmospheric N fixation, with equal or higher percent fixation of vetch in biculture at all site-years. More studies should be performed to better understand controls on N fixation of legume cover crops in cold climates.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Nitrogen is often a limiting nutrient in organic cropping systems (Berry et al. 2002). Organic farmers may employ legumes as a way to provide substantial N through biological nitrogen fixation (BNF) and delivery of N to soils through incorporation of nitrogen-rich plant biomass (Fageria et al. 2005). Legumes grown as cover crops may convey additional benefits, such as diversification (Tiemann et al. 2015), stabilization of plant–microbe metabolic cycles (Blagodatskaya et al. 2014), erosion control (Basche et al. 2016), nutrient management (Bergtold et al. 2012; Finney et al. 2016; Syswerda et al. 2012), weed suppression (Leavitt et al. 2011), disruption of pest cycles (Ratnadass et al. 2012), increased water infiltration (Baumhardt et al. 2015), and temperature stabilization (Basche et al. 2016).
Growing cool-season legumes such as winter annuals may deliver these benefits while enhancing soil N stocks, with evidence suggesting that winter annual legume cover crops can accumulate enough biomass nitrogen to maintain a warm-season cash crop in various environments (Fageria et al. 2005; Parr et al. 2011; Tonitto et al. 2006). A meta-analysis showed no difference in yield between legume cover crops and conventional fallow with application of recommended N when the legumes netted more than 110 kg N ha−1 in aboveground biomass (Tonitto et al. 2006). While winter annual legumes have been successful in other more southern regions of the U.S, the Upper Midwest presents unique challenges, such as: (1) short growing seasons that limit establishment and biomass production; (2) extreme minimum temperatures and variable snow cover leading to winter kill; and (3) wet springs that limit field working conditions. Despite these challenges, legume cover crops have been successfully managed in more northern climates for nitrogen delivery to a cash crop (Griffin et al. 2000; Lawson et al. 2012), yet we lack information about the capacity of legumes to provide supplemental fixed N to cash crops in cold climates.
In addition, cold temperatures have varying effects on the legume-rhizobia mutualism responsible for successful BNF and higher tissue N. Some temperate legume species may experience decreased nodulation and BNF in cold climates, while other arctic species can fix comparable amounts of N to temperate legumes, suggesting successful adaptation (Bordeleau and Prévost 1994). Nodulation of the legume cover crop hairy vetch has been shown to be reduced in temperatures common to the Upper Midwest (Perkus 2018).
Legume seeds are commonly inoculated with a host-specific rhizobia species and strain to optimize BNF. However, inoculation of legumes with highly effective rhizobia often fails when transferred to the field due to competition with existing endogenous rhizobia (Streeter 1994; Triplett and Sadowsky 1992). The success of the inoculant in the field may depend on genetic specificity (Mothapo et al. 2013a; Thies et al. 1992), environment (Pandey et al. 2017; Parr et al. 2011), soil pH (Wongphatcharachai et al. 2015), and competition with endogenous strains (Brockwell et al. 1995; Kosslak et al. 1983; Thies et al. 1992). Plant available soil N in the form of nitrate is also well-known to inhibit nodulation (Streeter 1985). Little is known about the ability of inoculant rhizobia to compete with the twin challenges of competition with existing rhizobia and extremely cold and often widely fluctuating spring and fall temperatures, the time when cool season legumes are required to be most productive.
The productivity of legume cover crops in terms of nitrogen accumulation is dependent on and amplified by the amount of biomass produced (Unkovich et al. 2010; Wortman and Dawson 2015). Hairy vetch (Vicia villosa Roth), a common cover crop, has been shown to produce between 2–8 Mg ha−1 of biomass across a range of climates (Cook et al. 2010; Parr et al. 2011; Poffenbarger et al. 2015a, b; Teasdale et al. 2004), though productivity in more northern regions is limited (Leavitt et al. 2011). Hairy vetch biomass N in warmer climates may reach 217 kg N ha−1 (Parr et al. 2011), but as much as 168 kg N ha−1 has been documented as far north as Wisconsin (Stute and Posner 1995).
Grass-legume bicultures, such as cereal rye and hairy vetch (Secale cereale L.-Vicia villosa Roth), may provide an advantage by both providing and retaining N resources in agroecosystems (Ranells and Wagger 1997; White et al. 2016) and have been shown to increase productivity, total nitrogen, and nitrogen fixation over legume monocultures (Poffenbarger et al. 2015a, b; Sainju et al. 2005; Schipanski and Drinkwater 2012). Brainard et al. (2012) found that vetch monocultures in Michigan contained on average 58% of tissue N from BNF (%Ndfa; N derived from the atmosphere), with vetch-cereal rye biculture containing higher at 70% Ndfa.
Studies evaluating N fixation potential in cold climates are limited, and legume-rhizobia relationships from crops that have historically been grown in warmer climates is underexplored in the Upper Midwest. The specific objectives of this study were to (1) evaluate winter rye, medium red clover, two ecotypes of hairy vetch, and a rye-vetch biculture as cover crops for N productivity; (2) determine the effect of inoculation on legume nodulation, total N, and %Ndfa; and (3) elucidate relationships between soil residual N, nodulation, and cover crop productivity in a sweet corn production system at two sites in Minnesota. Sweet corn was chosen as a short-season crop relevant to Minnesota, which produces more sweet corn than any other U.S. state (NASS—Quick Stats). We hypothesize that cereal rye, commonly grown in the Upper Midwest as a cover crop, will produce more biomass than, but similar amounts of N to legume cover crops. We further hypothesize that endogenous rhizobia strains will compete with inoculants for nodule occupancy, but that legumes will contribute fixed nitrogen regardless of rhizobia source.
Methods
Site descriptions and climate
This study was conducted at two sites in 2015–2016 and 2016–2017 on certified organic land in Grand Rapids, MN (LAT: 47.243347, LONG: − 93.492622) and Lamberton, MN (LAT: 44.239366, LONG: − 95.309855). The North Central Research and Outreach Center in Grand Rapids, MN is composed of a well-drained loamy silt sand over sand superactive Haplic Glossudalf and is considered atypical farmland with 100–120 frost-free days annually (NRCS Web Soil Survey 2017). The research site was certified organic in 2015 and had recently been converted to cropland from an apple orchard characterized by rhizomatous quackgrass (Agropyron repens) and broadleaf weeds. Snow cover was comparable in both years of the study, but 2016–2017 experienced an early fall 410 mm snowfall and hard freeze with minimum temperatures of − 10 °C that subsequently melted and was followed by a fall freeze–thaw cycle. The Southwest Research and Outreach Center in Lamberton, MN research site was located on a portion of the Elwell Agroecology Farm, characterized by a calcium carbonate fine silt superactive, mesic Typic Hapludoll considered prime farmland with 140–180 frost-free days annually (NRCS Web Soil Survey 2017). The research site was certified organic in 1993 and was characterized by common agricultural broadleaf weeds, primarily Canada thistle (Cirsium arvense) and lamb’s quarters (Chenopodium album). Lamberton 2015–2016 experienced growing degree days (GDD) on par with the site average since 1961 while 2016–2017 was warmer than average in the fall and close to average in the spring (Fig. 1). Precipitation in both years was higher than the 30-year average in the spring and fall (Fig. 2). Additional soil and climate characteristics are shown in Table 1.

Growing degree days (GDD) in each study environment throughout the cover crop growing season (a) and sweet corn growing season (b). GDD = (Tmax + Tmin)/2 − Tbase, where Tmax is maximum daily temperature, Tmin is minimum daily temperature, and Tbase is the base temperature of the organism, set at 4 °C. Midwestern Regional Climate Center

Monthly average precipitation (millimeters) at Grand Rapids, MN (GR) and Lamberton, MN (LAMB) during the period of September–August (2015–2016 and 2016–2017) and 30-year average precipitation at both locations during same time period. At both locations, fall 2016 was unseasonably wet. LAMB received 133.6 and 71.6 mm of precipitation, compared to 30-year state averages of 77.05 mm and 48.15 mm in September and October, respectively. In September 2016, GR received 141.99 mm, compared to a 30-year state average of 78.18 mm. LAMB received above average spring rainfall in both years, with 141.22 mm and 151.89 mm of precipitation in 2016 and 2017 respectively, compared to a 30-year average of 97.54. In 2016, GR was unseasonably dry during July, receiving 56.13 mm compared to a 30-year average of 104.09 mm. Midwestern Regional Climate Center
Experimental design
The experiment was a randomized complete block design with 4 replications. There were six treatments consisting of 4 legume cover crop treatments, including: (1, 2) two common cold-hardy ecotypes of hairy vetch (Vicia villosa Roth), variety not stated (V1 and V2); (3) medium red clover (Trifolium pratense L., CLO); (4) a cereal rye-hairy vetch biculture (Secale cereale L.-Vicia villosa Roth, MIX); (5) a non-legume control of cereal rye (Secale cereale L., RYE); and (6) a fallow control (noCC). In year one (2015–2016), four additional treatments were planted including an inoculated and a non-inoculated treatment in all legume-containing plots, totaling ten treatments. In year two (2016–2017), all legume cover crop treatments were inoculated. At Grand Rapids in 2016–2017, V1 was dropped due to insufficient seed quantities. Plot locations did not overlap from 2015–2016 to 2016–2017.
Cultural practices
Inoculation
For inoculated treatments in 2015–2016 and all legume seeds in 2016–2017, vetch seeds were inoculated with a peat-based inoculum of Rhizobium leguminosarm biovar. viceae (Verdesian LifeSciences, Cary, NC) by creating a 4:1 sucrose slurry that coated the seeds before applying the inoculant at the recommended rate. Seeds were allowed to dry and kept in a cool, dark place until planting, approximately 1 week. Medium red clover seeds were pre-inoculated with Sinorhizobium meliloti (Monsanto BioAg, St. Louis, MO) at the recommended rate. Both inocula are OMRI certified.
Cover crop planting
Cover crops were planted with a 3- or 6-row JP-6 Jang seeder (Mechanical Transplanter, Holland, MI) at Grand Rapids and with a Marliss drill (Remlinger Manufacturing Company, Kalida, OH) at Lamberton in late August or early September (see Table 2). Plots were 3.0 m × 7.6 m and planted lengthwise with rows running N-S and planted according to recommended seeding rates (see Table 3). Treatments in Grand Rapids received overhead irrigation as-needed during fall establishment. Treatments in Lamberton received no irrigation.
Cover crop termination and corn planting
Cover crops were terminated in late May/early June at both locations to ensure adequate development of sweet corn, a common horticultural crop grown in Midwestern cropping rotations for the processing industry typically following soybean or canning peas. Staggered planting from early May through late June is typical to meet processing contracts in this region (O’Rourke and Hutchison 2001), so delaying cover crop termination for increased crop maturity and biomass inputs may risk sweet corn production and crop insurance (Processing Sweet Corn: Iowa, Minnesota, Wisconsin 2017). In Grand Rapids, cover crops were terminated with a Toro Z Master Z253 riding mower with a custom fit expanded metal cover in 2015–2016 (The Toro Company, Bloomington, MN) and a FM62-H flail mower in 2016–2017 (Woodmaxx Power Equipment Ltd., Akron, NY). Residue was raked evenly over the plots and was immediately incorporated in 2015–2016. In 2016–2017, residue was allowed to dry for 7 d prior to incorporation with a TG-60-YK tiller (King Kutter Inc., Winfield, AL). Cover crops in Lamberton were terminated with a 90 M flail mower (Loftness, Hector, MN) and allowed to dry for 3 d in 2015–2016 and 4 d in 2016–2017 prior to direct incorporation by a JD-235 tandem folding disk (Deere & Co., Moline, IL).
In preparation for sweet corn planting, fields were disked in Grand Rapids by an IH 485 disk (Case IH, Racine, WI) and disked and dragged in Lamberton by a JD-235 and an HR 3004 D rotary power harrow (Kuhn, Brodhead, WI), respectively. ‘Luscious’ variety organic sweet corn (Johnny’s Selected Seeds, Fairfield, ME) was seeded at both locations at 86,487 seeds ha−1. Corn was planted by a model 90 single-row seed drill (HEGE Equipment Inc., Colwich, KS) at Grand Rapids and a 4-row precision cone planter with twin finger units (Kincaid, Haven, KS) at Lamberton. Four 7.6-m rows were seeded per plot.
Corn management
At early stages of development, corn was cultivated with a field cultivator with S-tines in both locations. Corn was weeded by hand-hoeing every two weeks beginning three weeks after planting and continued until canopy closure.
Nitrogen recommendations for sweet corn in medium-to-high N soils in Minnesota following oats, rye, or wheat is approximately 135 kg ha−1 (Rosen and Eliason 2005). Corn was sidedressed with 56 kg ha−1 8-2-4 composted turkey litter and feather meal (Sustane, Cannon Falls, MN) at 6–8 wk after planting to ensure for a measurable harvest after a pilot study indicated that additional N was necessary for any corn production.
Sampling and analysis
Biomass sampling
Biomass was sampled 0–2 d prior to termination at all site-years. Four 0.1 m2 quadrats were randomly cut to ground level and pooled from each plot. Biomass was separated by cover crop species or weeds, dried for 2–7 d at 60 °C, and weighed to generate dry weight per hectare. All plant material was ground through a 2 mm sieve and further pulverized with a 2010 Geno/Grinder ball grinder (SPEX SamplePrep, Metuchen, NJ) using a 5 mm diameter stainless steel ball bearing (Craig Ball Sales, Seaford, DE) for 3–10 min at 1500 rpm and stored at room temperature prior to elemental analysis.
Nodule sampling and rhizobia characterization
To determine inoculant effectiveness, three legume plants were randomly selected at both sites from inoculated and uninoculated plots containing a legume species in May 2016 in Lamberton and June 2016 in Grand Rapids, prior to cover crop termination. Plant roots were carefully dug from a 300-mm radius from the plant stem measuring approximately 300 mm deep, lightly shaken to remove loose soil from the root ball, and gently washed in a bucket of water. Roots were wrapped in a moist paper towel and stored in an air-tight Ziploc® bag at 4 °C for up to 1 week. Nodules were removed from the root, counted, and dried at 60 °C before determining nodule dry mass. Three nodules were removed from each plant and stored separately for molecular characterization.
Molecular analysis of nodule occupants comparing inoculated and uninoculated treatments was used to determine rhizobia source as either native soil or inoculant. The three nodules previously removed from each plant were surface sterilized, crushed, and streaked on YMA media plates supplemented with congo red for rhizobia selection and isolation. Single isolated strains were then subjected to 16S rRNA gene sequencing using standard approaches. Rhizobia (R. leguminosarum) DNA sequences were obtained for the small subunit ribosomal RNA (16S rRNA) gene of 117 rhizobial strains isolated from four legume hosts (V1, V2, MIX and CLO) in Lamberton, MN and 124 in Grand Rapids.
A phylogenetic tree was developed showing the relationship of the isolated strains based on partial 16S rRNA gene sequences (550 bp). Phylogenetic analysis was conducted in MEGA, version 7. The evolutionary history was inferred using the Neighbor-Joining method, with optimal tree with the sum of branch length = 0.26513004. Evolutionary distances were computed using the Maximum Composite Likelihood method with units of the number of base substitutions per site. Codon positions included were 1st + 2nd + 3rd + Noncoding. All positions containing gaps and missing data were eliminated. R. leguminosarum bv. vicae, trifolii, and phaseolii were included in the BLAST search as outgroups.
Soil sampling and soil extractable N measurement
Eight to ten 25 mm diameter soil cores were taken per plot at the time of biomass sampling to 150 mm depth and aggregated and homogenized in a bucket. Soil samples were stored in air-tight Ziploc® bags to retain field moisture, kept at 4 °C until analysis for no longer than 7 days, then passed through a 2 mm sieve prior to analysis. To determine extractable N, 40 mL 1 M KCl was added to a 10 g subsample of field moist soil and shaken for 1 h at 240 rpm. Samples were removed from the shaker and allowed to settle for 1 h, and the extract was filtered through #1 Whatman qualitative paper filters (GE Healthcare Life Sciences, Marlborough, MA) and frozen at − 20 °C until analysis. Total extractable N was determined using Shimadzu TOC-L/TN analyzer (standalone model, Shimadzu, Kyoto, Kyoto Prefecture, Japan).
Corn harvest
Of the four 7.6 m rows seeded per plot, two 3 m sections of corn were selected for harvest from the two innermost rows. A 3 m pole was placed between the rows and all corn harvested from either side. Corn ears were immediately husked and categorized as marketable or unmarketable. Marketable corn was classified as being at least 170 mm in length and displaying less than 8% damage or immaturity modified from Griffin et al. (2000) and USDA standards (USDA AMS, n.d.). A fresh weight was obtained for each category for each plot.
Nitrogen-15 natural abundance determination of percentage of nitrogen derived from atmosphere
Nitrogen-15 natural abundance calculations
The percent nitrogen derived from the atmosphere (%Ndfa) is determined using three values: the δ15N from the field-grown legume species (δ15Nfix), the δ15N from a non-fixing reference crop accessing the same soil N pool (δ15Nref), and the δ15N from the legume species grown in the absence of substrate nitrogen and therefore entirely dependent on biological nitrogen fixation (B) where δ15N is the natural abundance of the 15N isotope in parts per thousand relative to atmospheric N2 (Unkovich et al. 2008). The formula to determine %Ndfa can be expressed as follows (Shearer and Kohl 1986):
In this study, an average of the δ15N from the reference plot of cereal rye in each block and the weeds within each plot was used as δ15Nref.
Element analysis and Nitrogen-15 natural abundance sample preparation
Ground plant biomass was homogenized and subsampled into tin capsules (EA Consumables, Pennsauken, NJ) at 10 mg ± < 0.040 mg in 2015–2016 and 10 mg ± < 0.010 mg in 2016–2017. Samples were analyzed using an Elementar vario PYRO cube EA-IRMS CNS analyzer (Elementar, Langenselbold, Germany) for carbon and nitrogen content and isotopic fractionation of 15N.
Isotope fractionation determination
Legume species were grown in a GCW-15 growth chamber (Environmental Growth Chambers, Chagrin Falls, OH) at the University of Minnesota to determine B values with native rhizobia after preliminary analysis suggested that field-grown legumes were nodulated by endemic strains. Seeds were surface sterilized according to protocol modified from Somasegaran and Hoben (1994). Sterilized seeds were added to a sterile Petri dish and allowed to germinate for 16 days in a growth chamber.
Germinated seedlings were transferred to sterile Magenta units™ (bioWORLD, Dublin, OH) containing N-free medium composed of 50/50 v/v vermiculite and coarse sand. Three seedlings were added to each unit, which were thinned to one after 7 d. Seedlings were fertilized with dilute N-free nutrient solution (Broughton and Dilworth 1971) via cotton wick method as described by Rodriguez et al. 2008. At the time of transplant, seeds were inoculated with 1 mL rhizobia suspension containing four isolates, comprised of two representative strains from each field site, specific to the legume accession grown in a non-inoculated field treatment.
Seedlings were grown in the growth chamber until harvest in 3 replicated blocks. For the first 34 d, the plants were under the following conditions due to growth chamber error: 0800–1800 at 90 μmol intensity, 22 °C with 60% relative humidity. For the final 26 days, plants were grown under the following conditions: 0800–2200 at 300 μmol intensity, 22 °C:18 °C with ambient relative humidity.
Plants were harvested at 60 d, just prior to inflorescence, to mimic field maturity. Shoots were dried at 60 °C for 2 d and prepared for elemental analysis as described above. In this study, B values derived for clover were not viable, and a literature reference value was used (Schipanski and Drinkwater 2011).
Statistical Analysis
Data were analyzed using R statistical programming. All site-years were individually analyzed due to high variability. The experimental design was a complete randomized block design with a single combined fixed factor: cover crop + inoculation treatment. Concatenating cover crop and inoculation allowed comparison of legume treatments with non-legume treatments while contrasts were used to compare species or ecotype results. Block was treated as a random factor. Data was log-transformed to meet assumptions of homoscedasticity where Levene’s test was significant at α = 0.05. Outliers in the %Ndfa dataset were defined as outside of 1.5 times the interquartile range and removed from analysis (Tukey 1977). For Grand Rapids 2015–2016 %Ndfa data, a nonparametric robust analysis of variance (ANOVA) from the R package “Rfit” was used to account for heteroscedasticity and unbalanced data and a robust means comparison from the R package “rcompanion” was used. Preliminary analysis was accomplished using type II sums of squares ANOVA because no interactions were expected between treatment and block, and unbalanced data occurred when outliers were removed. Tukey’s honest significant difference (HSD) was used to separate means when p < 0.05. Weed data was not included in the final analysis.
Results
Cover crop biomass and N accumulation
Cover crop biomass and N accumulation differed by site-year and were affected by climatic factors that impacted growth and maturity (Table 4). At termination in all site-years, both vetch ecotypes had reached 0–10% flowering, while clover did not flower. Rye reached or surpassed 50% heading at both locations in 2015–2016, but only achieved stem elongation and early boot stage at Grand Rapids and early heading in Lamberton in 2016–2017. Cover crops were terminated at earlier than the 50% flowering optimal stage for maximum N accumulation due to increased risk to sweet corn yield with later planting (Williams 2008). In this experiment, sweet corn was planted in early-mid June, during the second half of the typical staggered planting in the region to meet processing contract requirements; the last day for planting to meet full crop insurance requirements is June 20 (Processing Sweet Corn: Iowa, Minnesota, Wisconsin 2017). Inoculation did not affect cover crop biomass or N of either species at either site (α = 0.05).
The highest biomass and N accumulation occurred at Lamberton in 2016–2017, with the rye-vetch biculture contributing significantly more biomass than V1, V2, and CLO at 9.3 Mg ha−1, but all treatments contributing statistically equivalent total N. The MIX and RYE treatments consistently yielded the most biomass across site-years, with V2 yielding equivalent biomass to MIX and RYE in Grand Rapids in 2015–2016. Both vetch monocultures produced the most biomass N among treatments in all site-years, as did MIX and RYE treatments with the exception of Grand Rapids in 2015–2016 due to the low proportion of legume by dry weight (16%) in the biculture compared to 31% vetch in Lamberton. While vetch was low in MIX treatments in 2016–2017 (2%), the rye biomass contained enough N to match the poor monoculture stands. There were no differences in biomass and N accumulation between vetch accessions. With the exception of 2016–2017 at Lamberton, CLO consistently contributed among the lowest amounts of N by cover crop treatment.
Effect of inoculation on nodulation and rhizobia characterization
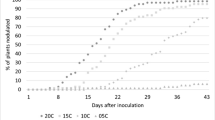
Clover had higher nodule numbers than did vetch plants, averaging 271 and 360 nodules per plant in Lamberton and Grand Rapids, respectively (Table 5). For hairy vetch, nodule number per plant was not affected by ecotype (V1 or V2) or biculture planting at either site. While clover showed the greater nodule number when compared to vetch, total nodule mass was lower than vetch, though this trend was not significant at either site (α = 0.05). Average vetch nodule mass in Grand Rapids (0.0975 g) was 1.5 times that of Lamberton (0.0647 g). Clover had similar nodule mass at both sites, though nodules were more numerous in Grand Rapids.
Nodule efficiency, or the ratio of energy exchange between the legume and its colonized rhizobia, is known to be a powerful determinant of biological nitrogen fixation in legume species. We used nodule mass as a proxy for nodule efficiency. Nodule efficiency in this study was not correlated with cover crop productivity (correlation = 0.05, p = 0.6941). Nodule mass and nodule number were either collinear with cover crop treatment (in which case they were removed from models) or not associated with shoot biomass, total N, or %Ndfa (α = 0.05). Additionally, nitrate is well understood to inhibit nodule formation and BNF activity at high levels (Streeter 1985). Soil extractable N at termination (Liebman 2018) did not impact nodule mass (p = 0.7192) or number (p = 0.1165) at either site.
The 16S rRNA phylogenetic trees from partial DNA sequences illustrated a close or even identical relationship between isolated field Rhizobium strains with Rhizobium leguminosarum species found at the Lamberton site, regardless to the legume host species or inoculant application (data not shown).
Nitrogen derived from the atmosphere
B values were determined for V1 and V2 treatments. CLO treatments failed to reach field maturity, so a reference value, − 1.65‰, from Schipanski and Drinkwater (2011) was used for N2 fixation calculations (Carlsson et al. 2006; Unkovich et al. 2008). No difference was found between the B values of V1 and V2 (p = 0.3206), so a single value was determined from the combined data. The B value for V1 and V2 was determined to be − 0.22 ± 0.01‰, which is higher than − 0.68 and − 0.53‰ as reported by Parr et al. (2011) in North Carolina, but similar to − 0.35‰ reported by Büchi et al. (2015) in Switzerland. B values may differ by location because different strains of rhizobia may fractionate 14N/15N differently (Carlsson et al. 2006; Unkovich et al. 2008).
The fractionation of 14N/15N in the soil at the field site must be significantly different from air for the natural abundance method to accurately measure %Ndfa (Shearer and Kohl 1986). In Grand Rapids in 2015–2016, very little isotopic fractionation occurred, resulting in several outliers and values outside the expected range (Tables 6, 7).
In all site-years, the MIX treatment exhibited among the highest %Ndfa (Table 6). Specifically, in three out of four site-years, the V2 in the MIX treatment had higher %Ndfa than the V2 monoculture, though it was not different from the V1 monoculture at either site in 2015–2016. Clover %Ndfa was equal to vetch monocultures in all site-years except in Grand Rapids in 2015–2016, where it was lower than V1.
Sweet corn yield
Sweet corn yields for both marketable and total yields were not different between inoculated and non-inoculated plots of the same cover crop treatment (data not shown). The variation in corn yield due to total biomass and total N was accounted for by cover crop treatment differences and were therefore removed from all models to avoid collinearity. Interestingly, total and marketable corn yields were not different between cover cropped plots and the bare ground control at all site-years with the exception of Lamberton in 2016–2017, where the fallow control plots yielded more than the clover plots (see Table 8). All treatments containing hairy vetch were among the top corn producers in all site-years, and rye consistently was among the lowest-producing treatments, yielding as little as 4.1 Mg ha−1 in husked fresh weight.
Corn yields were highest in Lamberton in 2015–2016 and lowest in Grand Rapids in 2016–2017. Corn in Grand Rapids failed to reach full maturity at both site-years, likely attributed to lower growing degree days (GDD; Fig. 2), and yield at this site was also impacted by poor germination. Lamberton husked weight corn yields in 2015–2016 reached 13.9 Mg ha−1 in the V2 treatment, nearing the state average unhusked yield of conventional sweet corn for processing of 16.5 Mg ha−1 in 2015 (Lofthus and Byrne 2016).
Discussion
Cover crop legumes in Minnesota produced sufficient biomass and fixed-N to contribute N in a majority of the site-years. Cover crop treatments varied in their productivity according to GDD, which was site-year dependent (see e.g. vetch, Table 9), and were not affected by inoculation treatment. This is consistent with other studies in the region; cover crop treatments produced more or similar biomass with the exception of Grand Rapids in 2016–2017, where GDD were lowest and a fall freeze–thaw likely contributed to vetch winter kill (e.g. Leavitt et al. 2011; Stute and Posner 1995).
Equivalent N between the legume monocultures and the RYE and MIX treatments is attributed to the higher N concentration of the legume monoculture treatments despite lower biomass when compared to RYE and MIX (Table 4). In 2016–2017 in Grand Rapids, the MIX biomass and N is entirely attributed to rye, as vetch demonstrated significant winter kill in both monoculture and biculture; lower maturity of rye may have contributed to equal amounts of N accumulation to the vetch monocultures in this site-year. The high biomass and N in Lamberton 2016–2017 compared to all other site-years is likely attributed to a warmer than average fall and higher minimum winter temperatures, which strengthened establishment and winter survival.
Cereal rye is a common winter annual cover crop in field corn systems in the Upper Midwest, with 18.4% of farmer respondents to a survey reporting using cereal rye as a cover crop and choosing it as the preferred cover crop after corn (Singer 2008). Although less common, legume cover crops such as clover and vetch are of increasing interest to field corn producers as a between-row intercrop (Rusch et al. 2019). Our study showed that in southern Minnesota (zone 4b), hairy vetch planted prior to corn as an overwintered cover crop can achieve canopy closure and accumulate biomass similar to Northeastern states such as New York (Teasdale et al. 2004), as well as nitrogen that is comparable to rye, but with a lower C:N ratio. However, this cropping system may limit staggered planting typical in this region to the latter half of normal planting dates. In more northern climates, variable springs and falls may be more likely to cause winter kill of crops that perform well in more southern regions of the Upper Midwest.
Following fallow in low organic matter soils, sweet corn requires up to 190 kg ha−1 N fertilizer in Minnesota; high organic matter soils may require 157 kg ha−1(Rosen and Eliason 2005). In Lamberton, the cover crops in our study mostly accumulated enough N to provide for a sweet corn crop in Minnesota, although proportional N release from decomposing cover crops in a season can vary widely (Poffenbargeret al. 2015a, b). If this is indeed the case, these cover crops may offset additional N inputs by approximately half. Winter cover crops may, in a short-season environment such as Minnesota, benefit from a fall application of fertilizer to provide a boost of fall growth to improve overwintering (Seguin et al. 2001). Additionally, legumes may effectively scavenge N from fall-applied manure or poultry litter, resulting in increased N delivery at spring cover crop termination (Seman-Varner et al. 2017). However, evolutionary theory suggests N fertilizer may cause legumes to select for more parasitic rhizobial strains, effectively fixing less N (Kiers et al. 2002). Additionally, N availability in the form of nitrate may inhibit nodulation (Streeter 1985), and tradeoffs between establishment and N fixation should be considered. In our study, soil extractable N at termination was not associated with nodulation or N fixation. It is possible that soil nitrate analysis at legume planting, and not at termination as in this study, would prove a better predictor of nodulation and BNF, as well as an analysis of nitrate specifically. Long-term economic and environmental tradeoffs of fertilizing cover crops should be evaluated to increase biomass and N production in northern growing regions.
Inoculation treatments may have failed to affect nodulation due to colonization by native strains via rhizobial competition. While the Grand Rapids organic site has not been cropped with legumes, Dutch white clover (Trifolium repens) has been cropped in adjacent fields, and common vetch (Vicia sativa) was identified in the adjacent woodlot. The Lamberton site has been organically cropped for two decades and was expected to harbor competitive rhizobia populations. High density endogenous populations may overcome high inoculation rates, and the endogenous population density may be the best predictor of inoculant efficacy (Thies et al. 1992).
Nodule mass appeared consistent with other literature and appears to be species-specific. In a study quantifying hairy vetch nodule mass in North Carolina soils, Mothapo et al. (2013b) reported lower nodule mass than we observed in Grand Rapids, but equal to that observed at Lamberton. In this study, clover nodules trended towards lower mass than vetch nodules, which is expected, as morphologically clover nodules may be smaller and more numerous than vetch nodules (Walsh 1995). Combined, nodule number and mass data suggests higher dependence on BNF in Grand Rapids, which was a field that was uncultivated for 10 years prior to this experiment and had lower organic matter concentrations. This phenomenon was not observed in 2016–2017 perhaps due to the site being in its second year of annual crop production, or the cumulative addition of crop residue to the soil prior to growing cover crops.
The MIX treatment exhibited among highest Ndfa, consistent with previous research hypothesizing that legumes in biculture have higher rhizobial efficiency due to N scarcity in the rhizosphere (Brainard et al. 2012; Poffenbarger et al. 2015a, b). Despite higher %Ndfa in the biculture, vetch monocultures fixed more or equivalent total N as the biculture. This evidence suggests that growers may improve %Ndfa by growing grass-legume bicultures, but should consider tradeoffs of production such as lower total legume biomass and potential cash crop germination challenges due to high rye biomass residue at termination. Other site-years exhibited %Ndfa values above 100% or below 0%, possibly due to the varying plant maturity across site-years was inconsistent with the single maturity attained in the growth chamber to determine a singular B value. These values were included in statistical analysis unless outliers. We also calculated %Ndfa using an average of within-plot weeds and within-block rye as a proxy for δ15Nref in the absence of a non-fixing isoline of red clover or vetch (Parr et al. 2011). The weed species and maturity varied significantly among plots, potentially skewing data outside of the expected range. These data are more appropriately interpreted for relative rather than absolute information.
Sweet corn yield was lowest in the rye plots, which may be attributed to three simultaneous factors: (1) the high C:N of rye may increase soil N immobilization during microbial decomposition (Liebman 2018); (2) the high biomass production of rye may be impeding seed-soil contact for efficient sweet corn germination; and/or (3) high biomass production in rye may provide habitat for seedcorn maggot (Delia platura Meigen). Seedcorn maggot predation was observed in Grand Rapids in both years of the study and may have impacted germination, despite delaying sweet corn planting by 11 d and 9 d in 2015–2016 and 2016–2017 respectively as recommended by Hammond and Cooper (1993). Seedcorn maggot predation was observed in similar cropping strategies in Maine (Griffin et al. 2000).
Including a winter annual legume into the cropping system did not decrease sweet corn total yield or marketable yield when compared to a non-cover cropped plot in our study when all treatments were partially fertilized. This is consistent with data from previous studies (Griffin et al. 2000; Miguez and Bollero 2006). While some studies show that legume cover crops alone cannot fully provide enough N for a sweet corn cash crop (Lawson et al. 2012; West et al. 2016), improved legume productivity can improve the overall N contribution. For example, in Maryland, hairy vetch alone was enough to provide full N for field corn when over 4 Mg ha−1 biomass produced, but not at lower biomass accumulation (Spargo et al. 2016). In our study, all vetch treatments reached or nearly reached 4 Mg ha−1, with the exception of Grand Rapids in 2016–2017. Additionally, the low N rate applied to all treatments may have interacted with cover crop additions. Miguez and Bollero (2005) reviewed corn response ratios to fertilizer added to legume, grass-legume biculture, and grass cover crops finding that bicultures have a positive response ratio with the addition of N fertilizer while legumes do not and grasses only have a slightly positive response ratio. Further research should elucidate fertilizer interactions with cover crop decomposition for winter annual cover crops in Minnesota.
Conclusion
Legumes are an important source of N fertility in organic cropping systems because they fix atmospheric N, effectively bringing new N into the soil system. Due to short growing seasons and narrow management windows in the Upper Midwest, winter annual legume cover crops cannot often reach their full growth and N fixation potential. Elaborating on mechanisms of N fixation in cold climates may yield new insights for improving C/N cycling in organic cropping systems. Future studies should focus on overcoming the barrier of rhizobial competition from endogenous strains and/or the development of an inoculum specific to cold climates. Further, the relationship between soil N availability and rhizobial efficiency should be assessed to determine critical turning points in rhizobial activity and evaluate the tradeoffs between N fertilization for enhanced legume cover crop biomass and the resulting BNF. These studies are needed to determine best management practices for increasing N fixation in winter annual legumes in cold climates.
Abbreviations
- BNF:
-
Biological nitrogen fixation
- GDD:
-
Growing degree days
- N:
-
Nitrogen
- ND:
-
Not determined
- Ndfa:
-
Nitrogen derived from the atmosphere
- OMRI:
-
Organic Materials Review Institute
References
Basche AD, Archontoulis SV, Kaspar TC, Jaynes DB, Parkin TB, Miguez FE (2016) Simulating long-term impacts of cover crops and climate change on crop production and environmental outcomes in the Midwestern United States. Agric Ecosyst Environ 218:95–106. https://doi.org/10.1016/j.agee.2015.11.011
Baumhardt RL, Stewart BA, Sainju UM (2015) North American soil degradation: Processes, practices, and mitigating strategies. Sustainability (Switzerland) 7(3):2936–2960. https://doi.org/10.3390/su7032936
Bergtold JS, Duffy PA, Hite D, Raper RL (2012) Demographic and management factors affecting the adoption and perceived yield benefit of winter cover crops in the Southeast. J Agric Appl Econ 44(1):99–116
Berry P, Sylvester-Bradley R, Philipps L, Hatch D, Cuttle S, Rayns F, Gosling P (2002) Is the productivity of organic farms restricted by the supply of available nitrogen? Soil Use Manag 18:248–255. https://doi.org/10.1079/SUM2002129
Blagodatskaya E, Blagodatsky S, Anderson TH, Kuzyakov Y (2014) Microbial growth and carbon use efficiency in the rhizosphere and root-free soil. PLoS ONE. https://doi.org/10.1371/journal.pone.0093282
Bordeleau LM, Prévost D (1994) Nodulation and nitrogen fixation in extreme environments. Plant Soil 161:115–125
Brainard D, Henshaw B, Snapp S (2012) Hairy vetch varieties and bi-cultures influence cover crop services in strip-tilled sweet corn. Agron J 104(3):629. https://doi.org/10.2134/agronj2011.0360
Brockwell J, Bottomley PJ, Thies JE (1995) Manipulation of rhizobia microflora for improving legume productivity and soil fertility: a critical assessment. Plant Soil 174:143–180
Broughton WJ, Dilworth MJ (1971) Control of leghaemoglobin synthesis in snake beans. Biochem J 125(4):1075–1080. https://doi.org/10.1042/bj1251075
Büchi L, Gebhard C-A, Liebisch F, Sinaj S, Ramseier H, Charles R (2015) Accumulation of biologically fixed nitrogen by legumes cultivated as cover crops in Switzerland. Plant Soil 393(1–2):163–175. https://doi.org/10.1007/s11104-015-2476-7
Carlsson G, Palmborg C, Huss-Danell K (2006) Discrimination against 15N in three N2-fixing Trifolium species as influenced by Rhizobium strain and plant age. Acta Agric Scand Sect B Soil Plant Sci 56(1):31–38. https://doi.org/10.1080/0906471051003131
Cook JC, Gallagher RS, Kaye JP, Lynch J, Bradley B (2010) Optimizing vetch nitrogen production and corn nitrogen accumulation under no-till management. Agron J 102:1491–1499. https://doi.org/10.2134/agronj2010.0165
Fageria NK, Baligar VC, Bailey BA (2005) Role of cover crops in improving soil and row crop productivity. Commun Soil Sci Plant Anal 36:2733–2757. https://doi.org/10.1080/00103620500303939
Finney DM, White CM, Kaye JP (2016) Biomass production and carbon/nitrogen ratio influence ecosystem services from cover crop mixtures. Agron J 108(1):39–52. https://doi.org/10.2134/agronj15.0182
Griffin T, Liebman M, Jemison J (2000) Cover crops for sweet corn production in a short-season environment. Agron J 92(1):144–151. https://doi.org/10.2134/agronj2000.921144x
Hammond RB, Cooper RL (1993) Interaction of planting times following the incorporation of a living, green cover crop and control measures on seedcorn maggot populations in soybean. Crop Prot 12(7):539–543. https://doi.org/10.1016/0261-2194(93)90096-2
Kiers ET, West SA, Denison RF (2002) Mediating mutualisms: Farm management practices and evolutionary changes in symbiont co-operation. J Appl Ecol 39(5):745–754. https://doi.org/10.1046/j.1365-2664.2002.00755.x
Kosslak RM, Bohlool BB, Dowdle S, Sadowsky MJ (1983) Competition of Rhizobium japonicum strains in early stages of soybean nodulation. Appl Environ Microbiol 46(4):870–873
Lawson A, Fortuna AM, Cogger C, Bary A, Stubbs T (2012) Nitrogen contribution of rye–hairy vetch cover crop mixtures to organically grown sweet corn. Renew Agric Food Syst 28(1):59–69. https://doi.org/10.1017/S1742170512000014
Leavitt MJ, Sheaffer CC, Wyse DL, Allan DL (2011) Rolled winter rye and hairy vetch cover crops lower weed density but reduce vegetable yields in no-tillage organic production. HortScience 46(3):387–395
Liebman A (2018) Legumes and soil organic matter transformations in Upper Midwest agroecosystems. University of Minnesota, Minneapolis
Lofthus D, Byrne T (2016) 2016 Agricultural statistics annual bulleton, Minnesota. St. Paul. https://www.nass.usda.gov/Publications/Ag_Statistics/
Miguez FE, Bollero GA (2005) Review of corn yield response under winter cover cropping systems using meta-analytic methods. Crop Sci 45(6):2318–2329. https://doi.org/10.2135/cropsci2005.0014
Miguez FE, Bollero GA (2006) Winter cover crops in Illinois: evaluation of ecophysiological characteristics of corn. Crop Sci 46(4):1536–1545. https://doi.org/10.2135/cropsci2005.09.0306
Mothapo N, Grossman J, Maul J, Shi W, Isleib T (2013a) Genetic diversity of resident soil rhizobia isolated from nodules of distinct hairy vetch (Vicia villosa Roth) genotypes. Appl Environ Microbiol 64:201–213
Mothapo NV, Grossman JM, Sooksa-nguan T, Maul J, Bräuer SL, Shi W (2013b) Cropping history affects nodulation and symbiotic efficiency of distinct hairy vetch (Vicia villosa Roth) genotypes with resident soil rhizobia. Biol Fertil Soils 49(7):871–879. https://doi.org/10.1007/s00374-013-0781-y
O’Rourke P, Hutchison WD (2001) Crop profile for sweet corn in Minnesota. Natl IPM Database. Retrieved from https://ipmdata.ipmcenters.org/documents/cropprofiles/MNsweetcorn.pdf
Pandey A, Li F, Askegaard M, Olesen JE (2017) Biological nitrogen fixation in three long-term organic and conventional arable crop rotation experiments in Denmark. Eur J Agron 90:87–95. https://doi.org/10.1016/j.eja.2017.07.009
Parr M, Grossman JM, Reberg-Horton SC, Brinton C, Crozier C (2011) Nitrogen delivery from legume cover crops in no-till organic corn production. Agron J 103(6):1578–1590. https://doi.org/10.2134/agronj2011.0007
Perkus EA (2018) Legume cover crops in high tunnels: field evaluation for soil health and controlled environment freezing tolerance. University of Minnesota, Minneapolis
Poffenbarger HJ, Mirsky SB, Weil RR, Kramer M, Spargo JT, Cavigelli MA (2015a) Legume proportion, poultry litter, and tillage effects on cover crop decomposition. Agron J 107(6):2083–2096. https://doi.org/10.2134/agronj15.0065
Poffenbarger HJ, Mirsky SB, Weil RR, Maul JE, Kramer M, Spargo JT, Cavigelli MA (2015b) Biomass and nitrogen content of hairy vetch–cereal rye cover crop mixtures as influenced by species proportions. Agron J 107(6):2069–2082. https://doi.org/10.2134/agronj14.0462
Processing Sweet Corn: Iowa, Minnesota, Wisconsin Processing Sweet Corn: Iowa, Minnesota, Wisconsin (2017) Saint Paul, MN
Ranells NN, Wagger MG (1997) Grass-legume bicultures as winter annual cover crops. Agron J 89:659–665. https://doi.org/10.2134/agronj1997.00021962008900040019x
Ratnadass A, Fernandes P, Avelino J, Habib R (2012) Plant species diversity for sustainable management of crop pests and diseases in agroecosystems: a review. Agron Sustain Dev 32(1):273–303. https://doi.org/10.1007/s13593-011-0022-4
Rodriguez RJ, Henson J, Van Volkenburgh E, Hoy M, Wright L, Beckwith F et al (2008) Stress tolerance in plants via habitat-adapted symbiosis. ISME J 2(4):404–416. https://doi.org/10.1038/ismej.2007.106
Rosen CJ, Eliason R (2005) Nutrient management in minnesota fruit & vegetable crops for commercial. University of Minnesota Extension, St Paul
Rusch H, Coulter J, Grossman J, Johnson G, Porter P, Garcia y Garcia A (2019) Interseeding cover crops in corn. https://blog-crop-news.extension.umn.edu/2019/06/interseeding-cover-crops-in-corn.html
Sainju UM, Whitehead WF, Singh BP (2005) Biculture legume-cereal cover crops for enhanced biomass yield and carbon and nitrogen. Agron J 97(5):1403–1412. https://doi.org/10.2134/agronj2004.0274
Schipanski ME, Drinkwater LE (2011) Nitrogen fixation of red clover interseeded with winter cereals across a management-induced fertility gradient. Nutr Cycl Agroecosyst 90(1):105–119. https://doi.org/10.1007/s10705-010-9415-z
Schipanski ME, Drinkwater LE (2012) Nitrogen fixation in annual and perennial legume-grass mixtures across a fertility gradient. Plant Soil 357(1):147–159. https://doi.org/10.1007/s11104-012-1137-3
Seguin P, Sheaffer CC, Ehlke NJ, Russelle MP, Graham PH (2001) Nitrogen fertilization and rhizobial inoculation effects on kura clover growth. Agron J 93(6):1262–1268
Seman-Varner R, Varco J, O’Rourke M (2017) Nitrogen benefits of winter cover crop and fall-applied poultry litter to corn. Agron J 109(6):2881–2888. https://doi.org/10.2134/agronj2016.11.0670
Shearer G, Kohl DH (1986) N2-fixation in field settings: estimations based on natural 15N abundance. Aust J Plant Physiol 13:699–756. https://doi.org/10.1016/0003-2697(92)90302-N
Singer JW (2008) Corn belt assessment of cover crop management and preferences. Agron J 100(6):1670–1672. https://doi.org/10.2134/agronj2008.0151
Soil Survey Staff, Natural Resources Conservation Service, United States Department of Agriculture. Web Soil Survey. Available online at: https://websoilsurvey.sc.egov.usda.gov/. Accessed 2017
Somasegaran P, Hoben HJ (1994) Handbook for rhizobia: methods in legume-rhizobium technology. Springer, New York
Spargo J, Cavigelli M, Mirsky S, Meisinger J, Ackroyd V (2016) Organic supplemental nitrogen sources for field corn production after a hairy vetch cover crop. Agron J 108(5):1992–2002. https://doi.org/10.2134/agronj2015.0485
Streeter JG (1985) Nitrate inhibition of legume nodule growth and activity. II. Short term studies with high nitrate supply. Plant Physiol 77(2):325–328. https://doi.org/10.1104/pp.77.2.325
Streeter JG (1994) Failure of inoculant rhizobia to overcome the dominance of indigenous strains for nodule formation. Can J Microbiol 40:513–522
Stute JK, Posner JL (1995) Synchrony between legume nitrogen release and corn demand in the upper Midwest. Agron J 87:1063–1069
Syswerda SP, Basso B, Hamilton SK, Tausig JB, Robertson GP (2012) Long-term nitrate loss along an agricultural intensity gradient in the Upper Midwest USA. Agric Ecosyst Environ 149:10–19. https://doi.org/10.1016/j.agee.2011.12.007
Teasdale JR, Devine TE, Mosjidis JA, Bellinder RR, Beste CE (2004) Growth and development of hairy vetch cultivars in the northeastern United States as influenced by planting and harvesting date. Agron J 96(5):1266–1271. https://doi.org/10.2134/agronj2004.1266
Thies JE, Bohlool BB, Singleton PW (1992) Environmental effects on competition for nodule occupancy between introduced and indigenous rhizobia and among introduced strains. Can J Microbiol 38:493–500
Tiemann LK, Grandy AS, Atkinson EE, Marin-Spiotta E, McDaniel MD (2015) Crop rotational diversity enhances belowground communities and functions in an agroecosystem. Ecol Lett 18(8):761–771. https://doi.org/10.1111/ele.12453
Tonitto C, David MB, Drinkwater LE (2006) Replacing bare fallows with cover crops in fertilizer-intensive cropping systems: A meta-analysis of crop yield and N dynamics. Agric Ecosyst Environ 112(1):58–72. https://doi.org/10.1016/j.agee.2005.07.003
Triplett EW, Sadowsky MJ (1992) Genetic of competition for nodulation of legumes. Annu Rev Microbiol 46:399–428
Tukey JW (1977) Exploratory data analysis, vol 2. https://pdfs.semanticscholar.org/2f40/fd06bc9fd00a27437e14ed171e96e4fd9326.pdf
Unkovich M, Herridge D, Peoples M, Cadisch G, Boddey B, Giller K et al (2008) Measuring plant-associated nitrogen fixation in agricultural systems. Canberra, Australia
Unkovich MJ, Baldock J, Peoples MB (2010) Prospects and problems of simple linear models for estimating symbiotic N2 fixation by crop and pasture legumes. Plant Soil 329(1):75–89. https://doi.org/10.1007/s11104-009-0136-5
USDA Agricultural Marketing Service (n.d.) Sweet corn for processing grades and standards. https://www.ams.usda.gov/grades-standards/sweet-corn-processing-grades-and-standards. Accessed 4 May 2015
Walsh KB (1995) Physiology of the legume nodule and its response to stress. Soil Biol Biochem 27(4–5):637–655. https://doi.org/10.1016/0038-0717(95)98644-4
West JR, Ruark MD, Bussan AJ, Colquhoun JB, Silva EM (2016) Nitrogen and weed management for organic sweet corn production on loamy sand. Agron J 108(2):758–769. https://doi.org/10.2134/agronj2015.0393
White CM, DuPont ST, Hautau M, Hartman D, Finney DM, Bradley B et al (2016) Managing the trade off between nitrogen supply and retention with cover crop mixtures. Agric Ecosyst Environ 237:121–133. https://doi.org/10.1016/j.agee.2016.12.016
Williams MM (2008) Sweet corn growth and yield responses to planting dates of the North Central United States. HortScience 43(6):1775–1779
Wongphatcharachai M, Staley C, Wang P, Moncada KM, Sheaffer CC, Sadowsky MJ (2015) Predominant populations of indigenous soybean-nodulating Bradyrhizobium japonicum strains obtained from organic farming systems in Minnesota. J Appl Microbiol 118:1152–1164. https://doi.org/10.1111/jam.12771
Wortman SE, Dawson JO (2015) Nitrogenase activity and nodule biomass of cowpea (Vigna unguiculata L Walp) decrease in cover crop mixtures. Commun Soil Sci Plant Anal 46(11):1443–1457. https://doi.org/10.1080/00103624.2015.1043457
Acknowledgements
The authors would like to thank Liz Perkus, Vivian Wauters, Dr. Peyton Ginakes, Dr. Fucui Li, Anne Pfeiffer, Charlotte Thurston, Dan Raskin, Dr. Michelle Dobbratz, Bruna de Bacco Lopes, Rachel Brann, Justin Panka, Emily Swanson, Caitlin Barnhart, Tori Hoeppner, Vick Hoffman, Kaleiilima Holt, and Yordanose Solomone for their field, lab, and data assistance. Additional thanks goes to Matt Galloway, Garett Heineck, and Dr. Andy Petran for their statistical contributions and Doug Brinkman for his growth chamber assistance. Finally, we thank Steve Quiring of the Southwest Research and Outreach Center in Lamberton as well as Keith Mann and Dani Sackett at the North Central Research and Outreach Center in Grand Rapids, in addition to their staff. We gratefully acknowledge the support of the North Central Sustainable Agriculture Research and Education program of USDA project number LNC14-364 and the University of Minnesota’s Forever Green Initiative for funding and the reviewers that graciously volunteered their time to provide this manuscript with meaningful feedback.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Perrone, S., Grossman, J., Liebman, A. et al. Nitrogen fixation and productivity of winter annual legume cover crops in Upper Midwest organic cropping systems. Nutr Cycl Agroecosyst 117, 61–76 (2020). https://doi.org/10.1007/s10705-020-10055-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10705-020-10055-z