
Abstract
Woody pastures represent keystone habitats for biodiversity in agricultural landscapes, contributing to increased resource availability, landscape heterogeneity and structural variability. High taxonomic diversity is closely linked to vegetation structure in woody pastures, but examining functional characteristics of species assemblages can shed more light on the ecological mechanisms driving divergent responses to habitat characteristics and help guide good management practices. To this end, we use a multi-taxa approach to investigate how plant, bat and bird taxonomic and functional diversity are affected by pasture tree and shrub density, structural complexity and proximate forest cover in southern Sweden. In particular, we use a trait exclusion approach to estimate the sensitivity of diversity-environment relationships to specific traits. We found little congruence between corresponding diversity metrics across taxa. Bird species richness responded stronger to environmental variables than functional diversity metrics, whereas the functional response to the environment was stronger than the taxonomic response among plants and bats. While increasing tree densities increased the taxonomic diversity of all three taxa, a simultaneous functional response was only evident for plants. Contrasting measures of vegetation structure affected different aspects of functional diversity across taxa, driven by different traits. For plants and birds, traits linked to resource use contributed particularly to the functional response, whereas body mass had stronger influence on bat functional diversity metrics. Multi-taxa functional approaches are essential to understand the effects of woody pasture structural attributes on biodiversity, and ultimately inform management guidelines to preserve the biological values in woody pastures.
Similar content being viewed by others


Introduction
With growing human pressure on nature, the quality of agricultural landscapes has become increasingly important for protecting biodiversity (Karp et al. 2012). Land use intensification leads to landscape simplification (Landis 2017) with homogenized biological communities as a consequence (Gámez-Virués et al. 2015). Woody pastures, consisting of a variety of grazed semi-natural grassland habitats with some tree cover (Bergmeier et al. 2010), contribute significantly to landscape heterogeneity and biodiversity (Lentini et al. 2012; Wood et al. 2017). Woody pastures have typically been shaped by a long history of silvo-pastoral management, where the combination of grazing and presence of old trees enables co-existence of grassland species and species associated with old growth forests (Bergmeier et al. 2010; Dorresteijn et al. 2013; Garbarino & Bergmeier 2014). Multiple taxonomic groups therefore benefit from the heterogeneous structure of woody pastures, among which species richness and composition of plants, bats and birds have been the most studied (e.g. Fischer et al. 2010; Jakobsson & Lindborg 2015; Wood et al. 2017). However, research suggests diverging responses to pasture tree densities across taxa. For example, plant species richness often peak when canopy cover is 30–50% (e.g. Rolo et al. 2016), but in some contexts plant diversity increases linearly with increasing tree densities (Jakobsson and Lindborg 2015). The trees in woody pastures are also known to enhance bat species richness (Lentini et al. 2012), and the mere presence of a few trees may act as keystone habitats for bats (Tews et al. 2004; Fischer et al. 2010). Species richness of birds in pastures tend to saturate at low to intermediate tree densities (Tellería 2001), although shifts in bird species composition may be more strongly affected compared to species richness (Jakobsson and Lindborg 2017).
Woody pastures have been declining throughout Europe despite their high cultural and natural values (Bergmeier et al. 2010). During the last decade, the European Union (EU) has regulated the number of trees farmers are allowed to have on their pastures to receive subsidies for enhancing biodiversity (European Commission 2014). Therefore, tree density has inadvertently become a main characteristic affecting management of woody pastures in many countries such as Sweden (Beaufoy et al. 2011). Using tree density in woody pastures as an indicator of their quality may be an intuitive way to guide management practices that are easy to manage in practice. However, not only trees but also other vegetation characteristics act as local filters for biological communities (Zobel et al. 1998) and contribute to habitat heterogeneity, which in turn increases niche space and hence species richness (Rosenzweig 1995; Benton et al. 2003). Vegetation attributes in general also tend to have contrasting effects on different organism groups. For example, increasing understory vegetation may decrease bat species richness (Kalda et al. 2015b) and increase bird species richness (Laiolo et al. 2004), and grassland structural complexity in terms of vegetation height and patchiness may positively affect birds and arthropods but not plants (Lengyel et al. 2016). Multi-taxa approaches that consider a variety of structural habitat components are therefore needed to understand how biodiversity responds to vegetation attributes in woody pastures.
In addition to studying multiple taxonomic groups, examining functional rather than taxonomic diversity may help elucidate why species richness in different taxonomic groups show diverging responses to habitat and landscape characteristics in woody pastures. In particular, combining metrics of taxonomic and functional diversity provides a more holistic assessment of the multifaceted nature of biodiversity (Devictor et al. 2010). Whereas taxonomic diversity has been the traditional focus in conservation science, functional diversity of organisms is a better predictor of ecosystem functioning than taxonomic diversity (Gagic et al. 2015). By considering traits that determine how organisms respond to environmental change, a variety of functional diversity metrics can be used to improve the mechanistic understanding of changes in community composition along environmental gradients (Mouillot et al. 2013). Multi-taxa functional approaches have been used in a few recent studies (e.g. Barbaro and van Halder 2008; Pedley and Dolman 2014; Jonason et al. 2017), although none of these have focused on pasture structure and tree density specifically. Furthermore, the above studies have not evaluated whether relationships between functional diversity and the environment are driven by similar traits across multiple taxonomic groups. If similar traits drive patterns in functional diversity metrics consistently across taxonomic groups, it might indicate that functional diversity across taxa is shaped by similar ecological mechanisms. As an example, wing morphology and foraging behavior of bat assemblages relate to landscape characteristics and habitat structure in agroecosystems (Cisneros et al. 2015) and may cause shifts in the functional response of bat communities. Similar responses can be expected concerning birds, where mobility traits and foraging traits could result in a functional shift along gradients of habitat and landscape structure (Karp et al. 2012; Trisos et al. 2014), although mobility traits in birds also respond to patterns in migration strategy or home range sizes (Runge et al. 2014; Dufour et al. 2019).
In this study, we evaluated how the structure of woody pastures affects biodiversity, by analyzing taxonomic and functional diversity in three taxonomic groups: plants, bats and birds. Whereas plants and birds are commonly used biodiversity indicators (Wolters et al. 2006), bats are an understudied group of conservation concern (Jones et al. 2009b). Furthermore, these three taxa have different ecological requirements regarding their occurrence within woody pastures, including environmental characteristics at different spatial scales (e.g. Söderström et al. 2001; Kalda et al. 2015a). Bats and in particular birds are mobile and respond to both local habitat quality and landscape context, whereas plants may respond more strongly to local habitat factors. Bats may be more dependent on feeding related aspects compared to birds, which show a larger variation in home-range size and dietary requirements.
Our primary objective was to analyze the relative importance of tree density and other local structural attributes of woody pastures of potential relevance from a management perspective on taxonomic diversity and multiple functional attributes of species assemblages across three different taxa. To estimate functional diversity, we used functional traits from plants, bats and birds that reflect variation between species regarding morphology, dispersal/mobility, height/foraging level, nutrient requirement/feeding, reproduction and life history. We analyzed the diversity metrics in relation to local structural vegetation attributes of the woody pastures by using tree density, shrub density and structural complexity, in parallel with controlling for proximate forest cover (see “Methods” for details).
We addressed the following three questions: (1) Are species richness and functional diversity metrics correlated among plants, bats and birds in woody pastures? (2) Are plant, bat and bird species richness and functional diversity metrics equally affected by structural vegetation attributes of woody pastures? and (3) How do specific traits and functional trait groups contribute to various functional diversity metrics, and their relations to structural vegetation attributes? We test the following hypotheses: (i) Diversity metrics generally show low congruence among the three taxa (Manning et al. 2015; Birkhofer et al. 2018). (ii) In contrast to species richness (cf. Jakobsson and Lindborg 2015, 2017; Wood et al. 2017), the functional response of bats and birds is more strongly linked to other structural vegetation attributes in and around the woody pastures than to tree density (Jung et al. 2012; Barbaro et al. 2014; Cisneros et al. 2015), whereas the functional response of plants is mainly affected by tree density (Rolo et al. 2016). Finally, (iii) traits belonging to the same functional groups (e.g. morphology) respond similarly to structural vegetation attributes and affect functional diversity metrics similarly across the three taxa (Luck et al. 2012; Dehling et al. 2014).
Methods
Study area
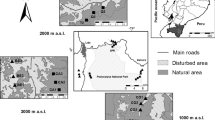
Our study area was the biosphere reserve Östra Vätterbranterna, located in the boreo-nemoral region of southern Sweden (Fig. 1). Mean temperature is 16.0 °C in July and − 1.9 °C in February and mean annual precipitation is 676 mm (SMHI 2020; average 1986–2015). In this area, the amount of woody pastures is relatively high compared to other parts of Sweden, creating a mosaic landscape structure with lowland arable fields, forest dominated hills and woody pastures in between (Jakobsson and Lindborg 2014). Woody pastures in this region are not fertilized and generally under low-intensity grazing pressure, mostly located on farms with cattle dominated rotational grazing systems, and tree densities in the study area range from just a few to more than 200 trees/ha (Jakobsson and Lindborg 2015). The landscape composition and the wide range of tree density makes this area well suitable for studying effects of variation in habitat characteristics across woody pastures under continuous low-intensity management. For this study, we selected 21 semi-natural woody pastures for biodiversity assessments, evenly distributed across the study area (Fig. 1; center points of the pastures > 1 km apart from each other) and with an average size of 4.1 ± 2.9 (SD) ha. All woody pastures fulfilled the five selection criteria predefined by Jakobsson and Lindborg (2015): (1) continuously grazed by livestock, (2) no recent and extensive clearing of trees, (3) fairly homogeneous tree density across the woody pasture, (4) situated on dry to semi-dry soil and (5) herb/grass dominated field layer. The pastures were individually fenced, of which a few belonged to the same properties, situated in the mosaic landscape. Although the studied woody pastures were rather small on average, and in particular birds and bats typically use multiple habitat types in mosaic landscapes such as our study area, the woody pastures have relatively characteristic species assemblages (Jakobsson and Lindborg 2017; Wood et al. 2017).

Modified from Jakobsson and Lindborg (2015)
The study area in southern Sweden, with the 21 woody pastures included in this study.
Data collection
For this comparative study, we compiled species community data of plants, birds and bats, together with environmental explanatory variables, from previously published studies by the authors (Jakobsson and Lindborg 2015, 2017; Wood et al. 2017). We were therefore limited to use data from different years for each taxon and chose to use the subset of 21 woody pastures for which data had been collected for all taxa. Local management and habitat conditions were similar during all study years, with negligible changes observed in vegetation attributes during field visits. Weather conditions were comparable across the three sampling years, with no evident deviation from the reference period (1986–2015) (Tables S1, S2). Hence, it is highly unlikely that habitat characteristics or weather conditions would not have been comparable across the three study years in our sites.
We recorded presence of vascular plant species (including shrubs and trees) in the summer 2013 using ten 1 m2 plots within a 0.8–1.4 ha subset of each pasture, giving a frequency of 0–10 for each species per pasture. We chose subsets to be representative areas of the pastures with relatively homogeneous tree cover (see Jakobsson and Lindborg (2015) for details on plant inventories). We surveyed birds using 50 m fixed radius point counts (5 min), using the pasture subset center as the observation point, with an additional slow walk within the 50 m radius circle (5 min). All birds seen or heard were recorded. We conducted the bird surveys within the first four hours from sunrise between 22 April and 15 June 2014 under favorable weather conditions (air temperature ≥ 5 °C, no heavy rainfall and wind speed < 5 m/s), repeated five times for each pasture (see Jakobsson and Lindborg (2017) for details on bird surveys). In the analysis, we pooled presence/absence species data from the five visits to calculate a relative frequency of 0–5 for each species per pasture. In 2015, we recorded bats during one night per pasture subset, using Song Meter SM3 bat detectors (Wildlife Acoustics Inc.) placed in the central point of the pasture subsets. We controlled for single night observations (cf. Fuentes-Montemayor et al. 2011; Kalda et al. 2015a, b; Barbaro et al. 2019) with strict weather condition criteria (air temperature ≥ 5 °C, no heavy rainfall and wind speed < 5 m/s) (Wood et al. 2017). We identified the calls manually using Sonobat v2.98 analysis software (https://sonobat.com) in parallel with the iBatsID identification system (Walters et al. 2012), and excluded the first 30 min after sunset and the last 30 min before sunrise to avoid the peaks of commuting to and from roosts (Wood et al. 2017). For this dataset, we used bat activity as a proxy for species abundance, estimated by the square root of the number of bat passes per night per species (cf. Barbaro et al. 2019). A bat pass was defined as two or more consecutive echolocation calls or one or more social calls (see Wood et al. (2017) for details on bat surveys). In some cases, bat calls could only be identified at the genus level, and we treated these observations as individual species in the analysis on bat species richness. When calculating functional diversity indices, we accounted for genus-level observations by using mean trait values of potential species within the genus occurring in the study area.
To estimate structural attributes within the woody pasture subsets, we identified all trees and shrubs and measured their diameter at breast height (DBH) (Jakobsson and Lindborg 2015). Based on those data, we calculated three explanatory variables to describe each pastures’ structure: tree density (trees/ha (> 10 cm DBH), shrub density (shrubs/ha), structural complexity (standard deviation of DBH) (Table 1). We used standard deviation of DBH as a measure of structural complexity because it correlates with most other complexity metrics (Neumann and Starlinger 2001) and is easily calculated using simple field measurements of tree sizes. Due to the small size of the habitat patches, some pasture subsets used for sampling were situated next to the habitat edge. To account for potential effects of adjacent habitats on the studied biological communities we calculated proximate forest cover (cf. Duflot et al. 2018) using interpretation of aerial photos within a 250 m buffer zone of the pastures (see Jakobsson and Lindborg (2015)). We used proximate forest cover instead of distance to the edge of the woody pastures because the contrast between woody pastures and the surrounding landscapes were typically not sharp in our study system. We also confirmed that including distance to the nearest forest patch had no qualitative effects on our final models (Table S3).
We aimed to match traits between the three taxa according to six main categories: morphology, dispersal/mobility, height/foraging level, nutrient requirement/feeding, reproduction and life history (Table 2). We chose these traits because they have all been shown to be relevant predictors of local biodiversity, and they represent well-known response traits to land-use change in the studied taxonomic groups (Hill et al. 1999; Garnier et al. 2007; Luck et al. 2012). We obtained plant traits from the life-history traits of the Northwest European Flora database (LEDA; Kleyer et al. 2008) and Hill et al. (1999) (see Table S4 for details), bird and bat traits from the Amniote database (Myhrvold et al. 2015) and Elton 1.0 database (Wilman et al. 2014). We extracted additional bird traits from Cramp et al. (1985–1994) and additional bat traits from the panTHERIA database (Jones et al., 2009a, b), Vaughan (1997), Safi and Kerth (2004), Bartonička et al. (2008), Ciechanowski and Zapart (2012) and https://www.nathusius.org.uk (Table 2). To avoid the effect of missing traits on functional diversity metrics (Pakeman 2014), we interpolated missing trait values using the mice package (van Buuren et al. 2015) in R. The values of a species’ other traits and their relationship to other species’ traits (excluding traits within the same trait group, see 2.3) were used to predict the missing trait values. In total, we interpolated 431 (7%) of the plant traits and one (< 1%) of the bat traits, whereas there were no missing traits for birds.
Using the same, or ecologically corresponding, traits to investigate effects of environmental factors on community functions of different taxa has been criticized due to diffuse trait-function relationships that differ between taxa (Pedley and Dolman 2014) and therefore taxon-specific approaches could be appropriate. However, Dehling et al. (2014) successfully investigated trait relationships between birds and plants in direct interaction networks of these taxa in the tropics. Similar analogies between bird and bat traits have been found in agricultural systems where these organism groups co-occur (Luck et al. 2012), although trait-function relationships may diverge substantially in terms of, for example, their foraging niches (Maas et al. 2016). We followed the rationale in Dehling et al. (2014) to compare corresponding trait groups across the three studied, co-occurring taxa in order to estimate which trait groups had the strongest effect on functional diversity metrics.
To avoid convergence of estimates of functional diversity (Lefcheck et al. 2015), we removed the allometric correlation between morphology (body mass) and mobility (forearm/wing length) for bats and birds by regressing forearm/wing length against body mass (body mass of birds was square root transformed for better linear fit). We thereafter used the residuals from this regression as our (relative) forearm/wing length variable (see Table S5a–c).
Analysis
We calculated species richness per taxon as the total number of species observed within each pasture during the study period. Using the selected traits, we calculated four multidimensional functional diversity metrics, weighted by trait groups (see Table 2): functional richness (FRic), functional evenness (FEve), functional divergence (FDiv) and Rao's Quadratic entropy (Rao’s Q) (function dbFD in R; Villéger et al. 2008) to use as response variables together with species richness in subsequent analysis. FRic is the incidence-based volume of the multidimensional functional space; FEve measures how evenly distributed traits are (abundance-weighted) within the multidimensional functional space; FDiv measures how divergent the community is (abundance-weighted) within its functional space, i.e. the proportion of species with extreme trait values; Rao’s Q is the abundance-weighted pairwise distance between species in the functional space. These four metrics were used in this study because they all have distinct functional meanings. FRic estimates the range of functions, without considering their distribution or dominance. FEve takes into account abundance of species and estimates if the functional space is effectively used (how evenly distributed traits are within the functional space), i.e. higher FEve values indicate more effective use (less parts of the space being underused), in which case additional species are also more likely to be functionally redundant (Luck et al. 2013). In contrast, FDiv uses abundance of species to estimate how the dominant species overlap in their use of the functional space, i.e. the higher the FDiv the less overlap (niche differentiation). Rao’s Q, instead, combines the other three metrics to estimate the mean functional similarity between two species within a community (Villéger et al. 2008).
First, we tested univariate correlations between all corresponding diversity metrics. Secondly, we used multiple linear models with stepwise model selection (function step in R; both directions) to analyze how functional diversity metrics and species richness of each taxon was affected by the four explanatory variables. Due to skewed distributions, we square root transformed shrub and tree density prior to analysis (Quinn and Keough 2002). To test for non-linear relationships, we compared univariate polynomial and linear responses (Chi-square tests; anova function in R) for tree density, shrub density and proximate forest cover prior to model construction. If the polynomial term improved the model (p < 0.10), we kept the term during the model selection process, and tested its significance (p < 0.05) in the final linear model. We checked residual plots of the full and final models to ensure assumptions of normality. Thirdly, we investigated how much different traits contribute to the functional diversity metrics and to the relationship between functional diversity metrics and environmental variables. We did this by recalculating each functional diversity metric after excluding the traits belonging to one of the groups and compared the model fit (adjusted R2) values of the original reduced linear models with models (the same reduced models) using the recalculated diversity metrics as response variables. To complement the last analysis, we conducted a fourth corner analysis (Dray and Legendre 2008) to test the non-parametric univariate associations between traits and environmental variables. All analyses were conducted in R (R Core Team 2017) using packages ade4 (Dray and Dufour 2007), FD (Laliberté et al. 2015), mice (van Buuren et al. 2015) and vegan (Oksanen et al. 2017).
Results
Diversity relationship
In total, we found 146 plant species, 10 (confirmed) bat species and 46 bird species within the 21 woody pastures (Table S6). Within taxa, FRic correlated positively with species richness for all taxa, whereas Rao’s Q correlated positively with species richness for plants and birds only. Between taxa, FRic of plants and bats were the only corresponding diversity metrics that were significantly correlated (Table 3).
Effects of environmental variables on plant, bat and bird diversity
The predictors explained more variation in bird species richness (43%) compared to plants and bats (≤ 18%) and to bird functional diversity metrics (≤ 22%). In contrast, the predictors explained more variation in plant Rao’s Q (37%) and bat FRic and FEve (30–31%) compared to species richness of plants and bats (Table 4).
Plant species richness, FRic and Rao’s Q showed similar positive responses to increasing tree density, whereas only Rao’s Q increased with increasing structural complexity (Fig. 2). Bat species richness and FDiv responded positively to increasing tree densities. Increasing bat FEve was driven by higher proportion of forest cover in the proximate landscape, whereas bat FRic increased up to about 50% forest cover and then dropped (Fig. 2). Bird species richness increased up to around 100 trees/ha and 70 shrubs/ha, after which it dropped in relation to higher tree and shrub densities (Fig. 2), whereas there was no significant functional response of birds to any measured environmental variable (Table 4).

Significant (see Table 4) relationships between taxonomic and functional diversity of plants, bats and birds in relation to the environment. Lines represent linear/polynomial fits to the data ± 95% CI (shaded area). Silhouettes from Freepik (www.freepik.com) and Vecteezy (www.vecteezy.com)
Effects of trait group exclusion on functional diversity metrics
For plants, excluding nutrient requirement traits from functional diversity metrics strongly reduced responses in plant FRic, whereas excluding plant height had similarly strong effects on responses in Rao’s Q (Table 4). Excluding morphology had the strongest effect on bat FRic and FDiv responses to proximate forest cover and tree density, respectively. In contrast, variability in plant and bat FEve was sensitive to the exclusion of multiple traits, namely foraging level/height, life history and dispersal/mobility. Bat Rao’s Q was not sensitive to the exclusion of any single trait (Table 4). Regarding birds, trait-specific contributions to the rather weak functional responses were absent except for the influence of foraging level on bird FRic and its (non-significant) response to shrubs and forest cover (Table 4).
Finally, trait-specific analyses revealed strong effects of environmental variables (tree density and proximate tree cover) on plant nutrient requirements and height, i.e. the same traits that had the strongest effect on functional diversity metrics (Table S7). Concerning bats, trait-specific responses to environmental drivers were generally weak, except for a significant relationship between proximate forest cover and feeding traits (Table S7). In contrast, tree and shrub density significantly affected bird feeding traits, foraging levels and reproduction (Table S7), although only reproduction considerably affected the functional response of birds.
Discussion
This study demonstrates a general lack of congruence between different diversity measures of plants, bats and birds in woody pastures in mosaic landscapes in southern Sweden. We also found contrasting effects of woody pasture characteristics on different taxa and diversity components, with increasing tree densities mainly benefiting taxonomic diversity whereas other vegetation attributes to a larger extent explained variation in the functional response. Concerning bats and plants, the functional response to woody pasture characteristics was stronger compared to the taxonomic response, whereas this was the opposite for birds, suggesting that a conservation strategy based of tree density in wood pastures fails to benefit both taxonomic and functional diversity in multiple organism groups. Finally, whereas traits directly linked to resource use (height/foraging level and nutrient requirement/feeding) had the strongest impact on the functional response in plants and birds, morphology (body mass) was most important in determining bat functional responses, most likely because bats showed less variation in resource use compared to plants and birds. In the following, we discuss the conservation implications of i) the contrasting taxonomic and functional responses across the three taxa, and ii) how our trait-based approach sheds light on the ecological mechanisms that underpin taxon-specific diversity patterns in woody pastures.
Correlations between diversity metrics
There was only one significant relationship between corresponding diversity metrics across taxa (FRic of plants and bats), confirming our first hypothesis (Maas et al. 2016; Birkhofer et al. 2018). Our results highlight that functional diversity in general does not increase in tandem with increasing taxonomic diversity in woody pastures. Low congruence between different indices of functional diversity is in line with earlier research (Jonason et al. 2017), but may also in part be a consequence of our approach to match trait data across taxa that differ in resolution between taxa, which may result in an underestimation of the covariance in functional diversity metrics between plants, bats and birds. As an example, we used Ellenberg indicator values as a proxy for resource use in plants, which only partly reflects the total nutrient requirements of plant species. In contrast to plants, we used the complete feeding gradient for bats and birds. Furthermore, bat species in our study area are 100% insectivorous and thus we used different arthropod taxa to quantify bat feeding traits, in contrast to the wider variation in bird feeding strategies. Whilst we note that alternative approaches to measure diet specialization among bats and birds are available (cf. Barbaro et al. 2019), we believe that our approach provides a good representation of diet preferences for analyzing functional diversity among bats and birds in our study system. However, because of these differences in trait resolution, we acknowledge that the relationships between a larger functional space in plant communities and higher trophic levels in our study should be interpreted with caution (Perner et al. 2005).
Diversity responses to the environment
In line with our second hypothesis, tree density had less pronounced effects on the functional response of bats and birds compared to species richness, whereas the positive effect of tree density on plant species richness extended to the functional response (Rao’s Q). This shows that tree density, more than other vegetation features, determine multi-taxa species richness, but that multiple vegetation features affect the multi-taxa functional response in woody pastures.
The plant communities showed similar functional responses as for species richness, with no decoupling of taxonomic and functional diversity metrics along the tree density gradient (cf. Rolo et al. 2016). Increasing tree density was associated with an increasing total functional space (FRic) in plants in woody pastures. We also show that increasing tree densities resulted in increasing plant functional diversity due to the addition of functionally different species (Rao’s Q) along the tree density gradient. Our results are therefore in line with the expectation that woody pastures create conditions that favor a variety of species that are associated to both open grassland and forests (Garbarino and Bergmeier 2014; Jakobsson and Lindborg 2015), which contributes to the unique conservation values of these habitats (Bergmeier et al. 2010).
Similar to Lentini et al. (2012), bat species richness increased with increasing tree density, though with rather low explanatory power, which may to some extent be explained by the fact that bat sampling was restricted to one night per site in our study. Notably, the functional response of bats to habitat characteristics was much stronger than the taxonomic response, probably because of expected guild-specific responses to vegetation features (Müller et al. 2012; Kalda et al. 2015a; Barbaro et al. 2019). Increasing FDiv with increasing tree density in woody pastures suggest that bat communities had lower overlap in functional space in conditions with high tree densities. In addition, a higher forest cover in the proximate landscape was associated with higher FRic in bat communities, suggesting that landscape context is important for the range of niches that bat communities span in woody pastures. Thus, our results confirm the importance of vegetation structures at multiple spatial scales on bat diversity in mosaic landscapes (Kalda et al. 2015a).
Bird species richness responded strongly to pasture habitat characteristics, peaking at intermediate tree and shrub densities, consistent with earlier research (Tellería 2001; Laiolo et al. 2004). In contrast, no explanatory variable had a significant effect on any of the bird functional diversity metrics. Birds commonly use multiple habitats scattered across agricultural landscapes, and landscape effects on bird communities are well known (e.g. Karp et al. 2012), and therefore it is not surprising that small-scale habitat structures in woody pastures did not significantly affect bird functional diversity metrics in our study. Although we acknowledge that our analyses may have suffered from low statistical power, studies based on large datasets have shown that landscape patterns even within relatively large extents (25 hectares) may have weak effects on bird functional diversity (Jonason et al. 2017). However, individual traits showed significant responses to the vegetation attributes, which indicate a stronger direct environment-trait link, but might also be an artifact from a larger trait variability gradient among birds compared to the other two taxa.
Because of short-term data collection over different sampling years for each taxon, we had limited precision in interpreting inter-taxa relationships and effects of the environmental variables. However, because weather and local management and habitat conditions were similar across all study years (see “Methods”), we believe that extant variation attributable to different sampling years had a minor influence on the relative relationship between multi-taxa functional diversity metrics and effects of explanatory variables. Hence, we believe that our analysis of the functional responses accurately reflects the main underlying ecological patterns in the woody pastures in our study. We would expect similar responses in comparable mosaic landscapes including habitats with woody cover (Tews et al. 2004), but in other systems the outcome will depend on the contrast between the focal habitats and properties of the surrounding landscapes.
Our main objective was to implement a holistic approach including taxonomic and functional characteristics of species assemblages to understand the biodiversity effects of pasture management, which are policy-driven practices that typically act at local scales without capturing the ecological context at larger scales (European Commission 2014; Ekroos et al. 2017). To this end, our study highlights how simplified management goals linked to what is regarded best management practice based on one taxonomic group (in many cases the plant community) (e.g. Minden et al. 2016; Tälle et al. 2016), or one habitat characteristic, may fail to enhance the broad majority of biological values (see also Söderström et al. 2001). We also note that including taxonomic groups from complementary trophic levels, with contrasting mobility and degree of habitat structure specialization (e.g. arthropods), would be needed to show the full potential of the effects of different management practices in woody pastures on biodiversity.
Drivers of functional diversity responses
In contrast to our third hypothesis, effects of corresponding trait groups on functional diversity metrics or specific traits’ relationship to the environment showed little congruence across taxa, suggesting that the ecological mechanisms determining community structure may differ substantially between the studied taxonomical groups. The trait exclusion approach showed that functionally distinct plant species regarding light and nutrient requirements contributed strongly to increasing functional richness (FRic) in plant communities as tree density increases. Trees create a heterogeneous environment where resource competition during plant community establishment enables a rich functional space (Belsky 1994; Martens et al. 2000; see also Jakobsson and Lindborg 2015). In addition, the same traits contributed to a parallel increase in plant Rao’s Q with increasing structural complexity, which emphasizes how vegetation complexity has a positive impact on plant functional diversity also at this small spatial scale (cf. Ma and Herzon 2014). However, only in the case of plants did the same traits that contributed most strongly to functional responses also respond individually to environmental factors. In contrast, bats’ functional responses were mainly linked to morphology (body mass), with additional effects of mobility (forearm length) and longevity. Variation in body mass contributed to increases in bat FRic and FDiv with increasing forest cover and tree density, respectively, indicating morphologically driven niche differentiation as a response to tree cover at local and proximate scales. We note that there is a relatively clear division of smaller (Myotis, Pipistrellus and Plecotus sp.) and larger (Eptesicus, Nyctalus and Verspertilio sp.) species in the study area, which might explain this link to morphology. The functional space in bat communities becomes more evenly distributed with increasing proximate forest cover because of (i) increases in interface specialists foraging within the semi-open pastures due to additional vegetation features in the surroundings (cf. Wood et al. 2017), (ii) increases in bat species with relatively short forearms, adapted to denser vegetation, without simultaneous losses of bats with long forearms, and (iii) increases in bats with higher longevity, because most long-lived species in our study area prefer forests. Our results confirm that heterogeneous vegetation structures are important for bats in semi-open habitats (Adams et al. 2009) by moderating the functional space of bat communities through species’ differences in general habitat preferences and maneuverability (Jung et al. 2012; Müller et al. 2012). In line with Ikin et al. (2012), variation in food preferences and foraging strategies among birds responded significantly to vegetation characteristics, especially increasing tree density, driving decreases in the number of ground foragers and seed eaters whilst favoring mid-high foraging birds (Table S7). However, among birds, functional diversity metrics remained unaffected, highlighting how individual traits of birds may be influenced by local habitat characteristics in woody pastures without resulting in an overall functional response.
Conclusions
Our study showed that there is little congruence between corresponding taxonomic and functional diversity metrics of plants, bats and birds in woody pastures. Further, we found that increasing tree densities in woody pastures benefitted species richness of all three taxa, whereas functional diversity metrics responded to a broader set of vegetation characteristics, including increasing shrub density, structural complexity, proximate forest cover and tree density. The results highlight the need to account for a broader suite of vegetation attributes in management prescriptions, rather than simply limiting high tree densities to preserve high functional diversity in woody pastures, especially regarding communities of mobile organisms like bats and birds. Furthermore, by using a trait group exclusion approach we were able to quantify the underlying ecological effects that contributed most to the observed functional responses. Notably, we found that the functional responses of plants and birds were driven by traits related to resource use, whereas body mass was the main driver of bat functional responses. Our study also shows that trait-specific responses are not necessarily directly linked to functional diversity responses (as for bats and birds), which may to some extent reflect the fact that birds and bats utilize multiple habitat types in mosaic landscapes. We therefore encourage further studies including similar approaches to ours, but in different habitat and landscape contexts. The absent links between diversity of these trophic levels and the complex responses to habitat structure show that multi-taxa functional approaches investigating effects of pasture attributes are needed to inform woody pasture management. By increasing the mechanistic understanding of multi-taxa functional diversity responses there are even better chances to preserve the high biological values in woody pastures in mosaic agricultural landscapes.
Data availability
Data used for this study are stored at Figshare (https://doi.org/10.6084/m9.figshare.9895727).
References
Adams MD, Law BS, French KO (2009) Vegetation structure influences the vertical stratification of open- and edge-space aerial-foraging bats in harvested forests. For Ecol Manage 258:2090–2100. https://doi.org/10.1111/j.1467-8330.1974.tb00606.x
Barbaro L, van Halder I (2008) Linking bird, carabid beetle and butterfly life-history traits to habitat fragmentation in mosaic landscapes. Ecography (Cop) 32:321–333. https://doi.org/10.1111/j.1600-0587.2008.05546.x
Barbaro L, Giffard B, Charbonnier Y et al (2014) Bird functional diversity enhances insectivory at forest edges: a transcontinental experiment. Divers Distrib 20:149–159. https://doi.org/10.1111/ddi.12132
Barbaro L, Allan E, Ampoorter E et al (2019) Biotic predictors complement models of bat and bird responses to climate and tree diverstiy in European forests. Proc R Soc B 286:20182193. https://doi.org/10.1098/rspb.2018.2193
Bartonička T, Řehák Z, Andreas M (2008) Diet composition and foraging activity of Pipistrellus pygmaeus in a floodplain forest. Biologia 63:1–7. https://doi.org/10.2478/s11756-008-0034-y
Beaufoy G, Jones G, Kazakova Y, McGurn P (2011) Permanent pastures and meadows under the CAP. The Situation in 6 Countries, European Forumon Nature Conservation and Pastoralism.
Belsky AJ (1994) Influences of trees on savanna productivity: tests of shade, nutrients, and tree-grass competition. Ecology 75:922–932. https://doi.org/10.2307/1939416
Benton TG, Vickery JA, Wilson JD (2003) Farmland biodiversity: is habitat heterogeneity the key? Trends Ecol Evol 18:182–188
Bergmeier E, Petermann J, Schröder E (2010) Geobotanical survey of wood-pasture habitats in Europe: diversity, threats and conservation. Biodivers Conserv 19:2995–3014. https://doi.org/10.1007/s10531-010-9872-3
Birkhofer K, Andersson GKS, Bengtsson J et al (2018) Relationships between multiple biodiversity components and ecosystem services along a landscape complexity gradient. Biol Conserv 218:247–253. https://doi.org/10.1016/j.biocon.2017.12.027
Ciechanowski M, Zapart A (2012) The diet of the pond bat and its seasonal variation in a forested lakeland of northern Poland. Acta Chiropter 14:73–79
Cisneros LM, Fagan ME, Willig MR (2015) Effects of human-modified landscapes on taxonomic, functional and phylogenetic dimensions of bat biodiversity. Divers Distrib 21:523–533. https://doi.org/10.1111/ddi.12277
Cramp S, Simmons KEL, Perrins CM (1985–1994) Handbook of the birds of Europe, the Middle East, and North Africa: the birds of the western Palearctic. Volume IV-IX. Oxford University Press, Oxford
Dehling DM, Töpfer T, Schaefer HM et al (2014) Functional relationships beyond species richness patterns: trait matching in plant-bird mutualisms across scales. Glob Ecol Biogeogr 23:1085–1093. https://doi.org/10.1111/geb.12193
Devictor V, Mouillot D, Meynard C, Jiguet F, Thuiller W, Mouquet N (2010) Spatial mismatch and congruence between taxonomic, phylogenetic and functional diversity: the need for integrative conservation strategies in a changing world. Ecol Lett 13:1030–1040. https://doi.org/10.1111/j.1461-0248.2010.01493.x
Dorresteijn I, Hartel T, von Wehrden H, Fischer J (2013) The conservation value of traditional rural landscapes: the case of woodpeckers in Transylvania, Romania. PLoS ONE 8:e65236. https://doi.org/10.1371/journal.pone.0065236
Dray S, Dufour A-B (2007) The ade4 package: Implementing the duality diagram for ecologists. J Stat Softw. https://doi.org/10.18637/jss.v022.i04
Dray S, Legendre P (2008) Testing the species traits–environment relationships: the fourth-corner problem revisited. Ecology 89:3400–3412. https://doi.org/10.1890/08-0349.1
Duflot R, Daniel H, Aviron S et al (2018) Adjacent woodlands rather than habitat connectivity infuence grassland plant, carabid and bird assemblages in farmland landscapes. Biodivers Conserv 27:1925–1942. https://doi.org/10.1007/s10531-018-1517-y
Dufour P, Descamps S, Chantepie S et al (2019) Reconstructing the geographic and climatic origins of long-distance bird migrations. J Biogeogr 47:155–166. https://doi.org/10.1111/jbi.13700
Ekroos J, Leventon J, Fischer J et al (2017) Embedding evidence on conservation interventions within a context of multi-level governance. Conserv Lett 10:139–145
European Commission (2014) Commission delegated regulation (EU) of 11.3.2014, supplementing Regulation (EU) No 1306/2013
Fischer J, Stott J, Law BS (2010) The disproportionate value of scattered trees. Biol Conserv 143:1564–1567. https://doi.org/10.1016/j.biocon.2010.03.030
Fuentes-Montemayor E, Goulson D, Park KJ (2011) Pipistrelle bats and their prey do not benefit from four widely appliedagri-environment management prescriptions. Biol Conserv 144:2233–2246
Gagic V, Bartomeus I, Jonsson T et al (2015) Functional identity and diversity of animals predict ecosystem functioning better than species-based indices. Proc R Soc B. https://doi.org/10.1098/rspb.2014.2620
Gámez-Virués S, Perovic DJ, Gossner MM et al (2015) Landscape simplification filters species traits and drives biotic homogenization. Nat Commun. https://doi.org/10.1038/ncomms9568
Garbarino M, Bergmeier E (2014) Plant and vegetation diversity in European wood-pastures. In: Hartel T, Plieninger T (eds) European wood-pastures in transition: a social-ecological approach, 1st edn. Routledge, Abingdon, pp 113–131
Garnier E, Lavorel S, Ansquer P et al (2007) Assessing the effects of land-use change on plant traits, communities and ecosystem functioning in grasslands: a standardized methodology and lessons from an application to 11 European Sites. Ann Bot 99:967–985
Hill MO, Mountford JO, Roy DB, Bunce RGH (1999) ECOFACT 2a Technical Annex: Ellenberg’s indicator values for British plants.
Ikin K, Knight E, Lindenmayer DB et al (2012) Linking bird species traits to vegetation characteristics in a future urban development zone: implications for urban planning. Urban Ecosyst 15:961–977. https://doi.org/10.1007/s11252-012-0247-2
Jakobsson S, Lindborg R (2015) Governing nature by numbers: EU subsidy regulations do not capture the unique values of woody pastures. Biol Conserv 191:1–9. https://doi.org/10.1016/j.biocon.2015.06.007
Jakobsson S, Lindborg R (2014) Wood pasture profile: East Vättern Scarp Landscape, Sweden. In: Hartel T, Plieninger T (eds) European wood-pastures in transition: a social-ecological approach, 1st edn. Routledge, Abingdon, pp 162–163
Jakobsson S, Lindborg R (2017) The importance of trees for woody pasture bird diversity and effects of the European Union’s tree density policy. J Appl Ecol 54:1638–1647. https://doi.org/10.1111/1365-2664.12871
Jonason D, Ekroos J, Öckinger E et al (2017) Weak functional response to agricultural landscape homogenisation among plants, butterflies and birds. Ecography (Cop) 40:1221–1230. https://doi.org/10.1111/ecog.02268
Jones KE, Bielby J, Cardillo M et al (2009a) PanTHERIA: a species-level database of life history, ecology, and geography of extant and recently extinct mammals. Ecology 90:2648–2648. https://doi.org/10.1890/08-1494.1
Jones G, Jacobs D, Kunz TH, Racey P (2009b) Carpe noctem: The importance of bats as bioindicators. Endanger Species Res 8:93–115. https://doi.org/10.3354/esr00182
Jung K, Kaiser S, Bo S et al (2012) Moving in three dimensions: effects of structural complexity on occurrence and activity of insectivorous bats in managed forest stands. J Appl Ecol 49:523–531. https://doi.org/10.1111/j.1365-2664.2012.02116.x
Kalda O, Kalda R, Liira J (2015a) Multi-scale ecology of insectivorous bats in agricultural landscapes. Agric Ecosyst Environ 199:105–113. https://doi.org/10.1016/j.agee.2014.08.028
Kalda R, Kalda O, Lõhmus K, Liira J (2015b) Multi-scale ecology of woodland bat the role of species pool, landscape complexity and stand structure. Biodivers Conserv 24:337–353. https://doi.org/10.1007/s10531-014-0811-6
Karp DS, Rominger AJ, Ranganathan J et al (2012) Intensive agriculture erodes β-diversity at large scales. Ecol Lett 15:963–970. https://doi.org/10.1111/j.1461-0248.2012.01815.x
Kleyer M, Bekker RM, Knevel IC et al (2008) The LEDA Traitbase: a database of life-history traits of the Northwest European flora. J Ecol 96:1266–1274. https://doi.org/10.1111/j.1365-2745.2008.01430.x
Laiolo P, Dondero F, Ciliento E, Rolando A (2004) Consequences of pastoral abandonment for the structure and diversity of the Alpine Avifauna. J Appl Ecol 41:294–304
Laliberté E, Legendre P, Bill Shipley (2015) Package “FD”: Measuring functional diversity (FD) from multiple traits, and other tools for functional ecology. R package v. 1.0-12. 1–28
Landis DA (2017) Designing agricultural landscapes for biodiversity-based ecosystem services. Basic Appl Ecol 18:1–12. https://doi.org/10.1016/j.baae.2016.07.005
Lefcheck JS, Bastazini VAG, Griffin JN (2015) Choosing and using multiple traits in functional diversity research. Environ Conserv 42:104–107. https://doi.org/10.1017/S0376892914000307
Lengyel S, Déri E, Magura T (2016) Species richness responses to structural or compositional habitat diversity between and within grassland patches: a multi-taxon approach. PLoS ONE. https://doi.org/10.1371/journal.pone.0149662
Lentini PE, Gibbons P, Fischer J et al (2012) Bats in a farming landscape benefit from linear remnants and unimproved pastures. PLoS ONE. https://doi.org/10.1371/journal.pone.0048201
Luck GW, Carter A, Smallbone L (2013) Changes in bird functional diversity across multiple land uses: interpretations of functional redundancy depend on functional group identity. PLoS ONE. https://doi.org/10.1371/journal.pone.0063671
Luck GW, Lavorel S, McIntyre S, Lumb K (2012) Improving the application of vertebrate trait-based frameworks to the study of ecosystem services. J Anim Ecol 81:1065–1076. https://doi.org/10.1111/j.1365-2656.2012.01974.x
Ma M, Herzon I (2014) Plant functional diversity in agricultural margins and fallow fields varies with landscape complexity level: Conservation implications. J Nat Conserv. https://doi.org/10.1016/j.jnc.2014.08.006
Maas B, Karp DS, Bumrungsri S et al (2016) Bird and bat predation services in tropical forests and agroforestry landscapes. Biol Rev 91:1081–1101. https://doi.org/10.1111/brv.12211
Manning P, Gossner MM, Bossdorf O et al (2015) Grassland management intensification weakens the associations among the diversities of multiple plant and animal taxa. Ecology 96:1492–1501. https://doi.org/10.1890/14-1307.1
Martens SN, Breshears DD, Meyer CW (2000) Spatial distributions of understory light along the grassland/forest continuum: effects of cover, height, and spatial pattern of tree canopies. Ecol Model 126:79–93. https://doi.org/10.1016/S0304-3800(99)00188-X
Minden V, Scherber C, Cebrián Piqueras MA et al (2016) Consistent drivers of plant biodiversity across managed ecosystems. Philos Trans R Soc B 371:20150284. https://doi.org/10.1098/rstb.2015.0284
Mouillot D, Grahan NAJ, Villéger S et al (2013) A functional approach reveals community responses to disturbances. Trends Ecol Evol 28:167–177. https://doi.org/10.1016/j.tree.2012.10.004
Müller J, Mehr M, Bässler C et al (2012) Aggregative response in bats: prey abundance versus habitat. Oecologia 169:673–684. https://doi.org/10.1007/s00442-011-2247-y
Myhrvold NP, Baldridge E, Chan B et al (2015) An amniote life-history database to perform comparative analyses with birds, mammals, and reptiles. Ecology 96:3109. https://doi.org/10.5061/dryad.t6m96
Neumann M, Starlinger F (2001) The significance of different indices for stand structure and diversity in forests. For Ecol Manag 145:91–106
Oksanen J, Blanchet FG, Friendly M, Kindt R, Legendre P, McGlinn D, Minchin PR, O’Hara RB, Simpson GL, Solymos P, Stevens HH, Szoecs E, Wagner H (2017) vegan: community ecology package, v2.4-4
Pakeman RJ (2014) Functional trait metrics are sensitive to the completeness of the species’ trait data? Methods Ecol Evol 5:9–15. https://doi.org/10.1111/2041-210X.12136
Pedley SM, Dolman PM (2014) Multi-taxa trait and functional responses to physical disturbance. J Anim Ecol 83:1542–1552. https://doi.org/10.1111/1365-2656.12249
Perner J, Wytrykush C, Kahmen A et al (2005) Effects of plant diversity, plant productivity and habitat parameters on arthropod abundance in montane European grasslands. Ecography (Cop) 28:429–442. https://doi.org/10.1111/j.0906-7590.2005.04119.x
Quinn GP, Keough MJ (2002) Experimental design and data analysis for biologists, 1st edn. Cambridge University Press, New York
R Core Team (2017) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Rolo V, Rivest D, Lorente M et al (2016) Taxonomic and functional diversity in Mediterranean pastures: insights on the biodiversity-productivity trade-off. J Appl Ecol 53:1575–1584. https://doi.org/10.1111/1365-2664.12685
Rosenzweig ML (1995) Species diversity in space and time. Cambridge University Press, Cambridge
Runge CA, Martin TG, Possingham HP et al (2014) Conserving mobile species. Front Ecol Environ 12:395–402. https://doi.org/10.1890/130237
Safi K, Kerth G (2004) A comparative analysis of specialization and extinction risk in temperate-zone bats. Conserv Biol 18:1293–1303
SMHI (2020) Klimatdata: The Swedish Meteorological and Hydrological Institute. Station: Ramsjöholm (ID 74490). Retrieved 19 May 2020.
Söderström B, Svensson B, Vessby K, Glimskär A (2001) Plants, insects and birds in semi-natural pastures in relation to local habitat and landscape factors. Biodivers Conserv 10:1839–1863. https://doi.org/10.1023/A:1013153427422
Tälle M, Deák B, Poschlod P et al (2016) Grazing vs. mowing: a meta-analysis of biodiversity benefits for grassland management. Agric Ecosyst Environ 222:200–212. https://doi.org/10.1016/j.agee.2016.02.008
Tellería JL (2001) Passerine bird communities of Iberian dehesas: a review. Anim Biodivers Conserv 24:67–78
Tews J, Brose U, Grimm V et al (2004) Animal species diversity driven by habitat heterogeneity/diversity: the importance of keystone structures. J Biogeogr 31:79–92
Trisos CH, Petchey OL, Tobias JA (2014) Unraveling the interplay of community assembly processes acting on multiple niche axes across spatial scales. Am Nat 184:593–608. https://doi.org/10.1086/678233
van Buuren S, Groothuis-Oudshoorn K, Robitzsch A, et al (2015) Package “mice”: multivariate imputation by chained equations. R package v. 2.30
Vaughan N (1997) The diet of British bats (Chiroptera). Mamm Rev 27:77–94. https://doi.org/10.1111/j.1365-2907.1997.tb00373.x
Villéger S, Mason NWH, Mouillot D (2008) New multidimensional functional diversity indices for a multifaceted framework in functional ecology. Ecology 89:2290–2301. https://doi.org/10.1890/07-1206.1
Walters CL, Freeman R, Collen A et al (2012) A continental-scale tool for acoustic identification of European bats. J Appl Ecol 49:1064–1074. https://doi.org/10.1111/j.1365-2664.2012.02182.x
Wilman H, Belmaker J, Jennifer S et al (2014) EltonTraits 1.0: Species-level foraging attributes of the world’ s birds and mammals. Ecology 95:2027. https://doi.org/10.1890/13-1917.1
Wolters V, Bengtsson J, Zaitsev AS (2006) Relationship among the species richness of different taxa. Ecology 87:1886–1895. https://doi.org/10.1890/0012-9658(2006)87[1886:RATSRO]2.0.CO;2
Wood H, Lindborg R, Jakobsson S (2017) European Union tree density limits do not reflect bat diversity in wood-pastures. Biol Conserv 210:60–71. https://doi.org/10.1016/j.biocon.2017.04.001
Zobel M, van der Maarel E, Dupré C (1998) Species pool: the concept, its determination and significance for community restoration. Appl Veg Sci 1:55–66. https://doi.org/10.2307/1479085
Acknowledgements
The authors thank B. Regelin for field assistance, E. Andersson for comments on bird census method, I. Ahlen for help with identification of bats and two anonymous reviewers for valuable comments on the manuscript. The authors have no conflict of interest to declare.
Funding
Open Access funding provided by Norwegian institute for nature research. This project was financially supported by the Swedish Research Council for Environment, Agricultural Sciences and Spatial Planning (FORMAS; 2011-977), Ahlmann’s foundation and Albert and Maria Bergström’s foundation.
Author information
Authors and Affiliations
Contributions
SJ, HW, JE and RL conceived the ideas and designed methodology; SJ and HW collected the data, SJ analyzed the data and led the writing of the manuscript. All authors contributed critically to the drafts and gave final approval of publication.
Corresponding author
Additional information
Communicated by Neil Brummitt.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Jakobsson, S., Wood, H., Ekroos, J. et al. Contrasting multi-taxa functional diversity patterns along vegetation structure gradients of woody pastures. Biodivers Conserv 29, 3551–3572 (2020). https://doi.org/10.1007/s10531-020-02037-y
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10531-020-02037-y