
Summary
-
1.
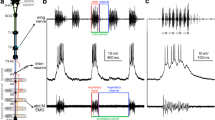
The calling, courtship and aggression songs ofT. oceanicus have distinctive temporal patterns but their spectral compositions are very similar: the carrier frequency occurs around 4.5 kHz and harmonics occur which extend above background noise level up to at least 40 kHz (Fig. 1).
-
2.
Units were recorded in the cervical connectives which responded to sound frequencies over the range from 1 kHz to at least 40 kHz. The sensitivities of some of the high frequency units were comparable to, or exceeded, the sensitivities of the units responding to the carrier frequencies of the species songs (Fig. 2).
-
3.
At the level of the neck connectives, units responding to the range from 10 kHz to 40 kHz could be divided, on the basis of their prothoracic morphology and physiology, into Through Neurones and Ascending Neurones. All the recorded neurones showed two-tone suppression effects but they varied in their degree of susceptibility to suppression. Frequencies in the range 4–5 kHz were most effective in producing suppression of the responses to high frequency tones (Figs. 5, 6).
-
4.
The two identified Ascending Neurone types had similar morphologies in the prothoracic ganglion (Fig. 3). ANA showed tonic responses to pure tones in the frequency range from 10 kHz to 40 kHz. Below 10 kHz high intensity inhibition of the response occurred and the latency increased from about 15 ms for an 18 kHz tone pulse, to about 55 ms for a 4.5 kHz tone (Fig. 4). ANB responded with 1–3 spikes to stimuli in the frequency range from 7 kHz to 40 kHz. The response latency to an 18 kHz tone pulse was about 15 ms. No responses were obtained to 50 ms stimuli in the range from 2 kHz to 7 kHz (Fig. 6). However, considerable rebound activity was produced in response to 4.5 kHz stimuli of 200 ms or more; the latency of the rebound to 200 ms stimuli was about 400 ms (Fig. 6C).
-
5.
ANA neurones responded to the syllables of the natural songs (Fig. 7), clearly coding the temporal patterns of the three songs. Cutting the contralateral leg nerve resulted in no detectable change in the neurone's response pattern. ANB neurones responded only during the intervals between chirps and during pauses within chirps. A long period of bursting activity occurred at the end of each song sequence (i.e. 3 chirps). This rebound activity may be the result of extreme inhibition caused by the CF of the songs (Fig. 7).
-
6.
Synthesised natural songs (Fig. 1 E) with the correct temporal parameters but filled with 4.5 kHz alone, 18 kHz alone or a combination of 18 kHz and 4.5 kHz, were produced to investigate the role of two-tone suppression phenomena in song coding. The responses of both ANA and ANB (Fig. 8) showed the interaction of excitatory and inhibitory inputs to these units during the coding of the species songs.
-
7.
Two-tone inhibition effects are advanced as the basic mechanism for the accurate coding of temporal characteristics in the songs ofT. oceanicus. Two different strategies for the recognition of species song types in crickets are advanced.
Similar content being viewed by others

Abbreviations
- ANA, ANB :
-
ascending neurones A, B
- CF :
-
carrier frequency
- CT :
-
control tone
- NS :
-
natural song
- SNS :
-
simulated natural song
- TN :
-
through neurone
- TT :
-
test tone
References
Bacon JM, Altman JS (1977) A silver intensification method for cobalt-filled neurones in wholemount preparations. Brain Res 138:359–363
Bentley D (1977) Control of cricket song patterns by descending interneurons. J Comp Physiol 116:19–38
Boyan GS (1981) Two-tone suppression of an identified auditory neurone in the brain of the cricketGryllus bimaculatus (DeGeer). J Comp Physiol 114:117–125
Elsner N, Popov AV (1978) Neuroethology of acoustic communication. Adv Insect Physiol 13:229–355
Hill KG (1974) Carrier frequency as a factor in phonotactic behaviour of female crickets (Teleogryllus commodus). J Comp Physiol 93:7–18
Hill KG (1975) Acoustic communication in the Australian field cricketsTeleogryllus commodus andT. oceanicus (Orthoptera: Gryllidae). PhD thesis, Melbourne University
Hill KG, Boyan GS (1977) Sensitivity to frequency and direction of sound in the auditory system of crickets (Gryllidae). J Comp Physiol 121:79–97
Hill KG, Loftus-Hills JJ, Gartside DF (1972) Premating isolation between the Australian field cricketsTeleogryllus commodus andT. oceanicus (Orthoptera: Gryllidae). Aust J Zool 20:153–163
Hill KG, Lewis DB, Hutchings ME, Coles RB (1980) Directional hearing in the Japanese quail (Coturnix coturnix japonica). I. Acoustic properties of the auditory system. J Exp Biol 86:135–151
Hutchings ME, Lewis DB (1981) Response properties of primary auditory fibres in the cricketTeleogryllus oceanicus (Le Guillou). J Comp Physiol 143:129–134
Kalmring K (1975) The afferent auditory pathway in the ventral cord ofLocusta migratoria (Acrididae). I–II. J Comp Physiol 104:103–159
Loftus-Hills JJ, Littlejohn MJ, Hill KG (1971) Auditory sensitivity of the cricketsTeleogryllus commodus andT. oceanicus. Nature New Biol 223:184–185
Loher W, Rence B (1978) The mating behaviour ofTeleogryllus commodus (Walker) and its central and peripheral control. Z Tierpsychol 46:225–259
Moiseff A (1980) Auditory interneurons and phonotactic behaviour in the Australian field cricketTeleogryllus oceanicus. PhD thesis, Cornell University
Oldfield BP (1980) Accuracy of orientation in female cricketsTeleogryllus oceanicus (Gryllidae): dependence on song spectrum. J Comp Physiol 141:93–99
Pollack AS, Hoy R (1981) Phonotaxis to individual rhythmic components of a complex cricket calling song. J Comp Physiol 144:367–373
Pollack AS, Plourde N (1982) Directionality of acoustic orientation in flying crickets. J Comp Physiol 146:207–215
Popov AV, Shuvalov VF (1977) Phonotactic behaviour of crickets. J Comp Physiol 119:111–126
Popov AV, Markovich AM (1982) Auditory interneurons in the prothoracic ganglion of the cricket,Gryllus bimaculatus. II. A high frequency ascending neuron (HF1AN). J Comp Physiol 146:351–359
Rheinlaender J (1975) Transmission of acoustic information at three neuronal levels in the auditory system ofDecticus verrucivorus (Tettigoniidae, Orthoptera). J Comp Physiol 97:1–53
Rheinlaender J, Kalmring K, Popov AV, Rehbein H (1976) Brain projections and information processing of biologically significant sounds by two large ventral cord neurons ofGryllus bimaculatus DeGeer (Orthoptera, Gryllidae). J Comp Physiol 110:251–269
Sachs MB, O'Connell NA (1983) Frequency analysis in the central auditory system of non-mammalian vertebrates. In: Lewis B (ed) Bioacoustics. A comparative approach. Academic Press, London, pp 283–310
Sachs MB, Young ED (1979) Encoding of steady-state vowels in the auditory nerve: Representation in terms of discharge rate. J Acoust Soc Am 66:470–479
Taylor MR (1978a) Trapezoidal waveform generator. New Electronics 11 (3):20
Taylor MR (1978b) Synchronised tone burst generator. New Electronics 11 (20):20
Thorson J, Weber T, Huber F (1982) Auditory behaviour in the cricket. II. Simplicity of calling song recognition inGryllus and anomalous phonotaxis at abnormal carrier frequencies. J Comp Physiol 146:361–378
Walker TJ (1957) Specificity in the response of female tree crickets (Orthoptera, Gryllidae, Oecanthidae) to calling songs of the males. Ann Entomol Soc 50:626–636
Weber T, Thorson J, Huber F (1981) Auditory behaviour of the cricket. I. Dynamics of compensated walking and discrimination paradigms on the Kramer treadmill. J Comp Physiol 141:215–232
Wohlers DW, Huber F (1978) Intracellular recording and staining of cricket auditory interneurons (Gryllus campestris L.,Gryllus bimaculatus DeGeer). J Comp Physiol 127:11–28
Wohlers DW, Huber F (1982) Processing of sound signals by six types of neurons in the prothoracic ganglion of the cricketGryllus campestris L. J Comp Physiol 146:161–173
Zaretsky MD (1972) Specificity of the calling sound and short term changes in the phonotactic response by female crickets,Scapsipedus marginatus (Gryllidae). J Comp Physiol 79:153–172
Author information
Authors and Affiliations
Rights and permissions
About this article
Cite this article
Hutchings, M., Lewis, B. The role of two-tone suppression in song coding by ventral cord neurones in the cricketTeleogryllus oceanicus (Le Guillou). J. Comp. Physiol. 154, 103–112 (1984). https://doi.org/10.1007/BF00605395
Accepted:
Issue Date:
DOI: https://doi.org/10.1007/BF00605395