
Abstract
Genetic technologies are necessary for understanding the molecular mechanisms of wide-ranging biological phenomena. However, genetic approaches are limited in the so-called model organisms. For example, genetics are not available in most marine invertebrates. Ciona intestinalis is a marine invertebrate chordate that provides excellent systems for studying allorecognition. We have established germline transformation with transposable elements, enhancer detection, and mutagenesis with transposons and customized nucleases. These genetic technologies are invaluable tools for uncovering the genetic functions underlying fertilization and allorecognition in this ascidian. In this chapter, achievements in the genetics of C. intestinalis are discussed.
You have full access to this open access chapter, Download conference paper PDF
Similar content being viewed by others

Keywords
1 Introduction
Ascidians, or sea squirts, are sessile invertebrates that live in the ocean (Fig. 38.1) (Satoh 1994). The larvae of ascidians are typical free-swimming tadpoles (Fig. 38.1a). As the larval tadpole body suggests, ascidians are closely related to vertebrates (Satoh 2003). Ascidians are members of the subphylum Urochordata, and belong to the phylum Chordata with cephalochordates and vertebrates. There are some basic body features that characterize chordates in addition to the tadpole body: a central nervous system consisting of a dorsally located neural tube, possession of a notochord at certain developmental stages, gill slits at the pharynx, and an endostyle/thyroid gland. Among chordates, urochordates are thought to be the most closely related to vertebrates (Delsuc et al. 2006).

The ascidian Ciona intestinalis. (a) A larva. No, notochord; SV, sensory vesicle (brain); Tu, tunic. (b) A juvenile. Larvae convert their tadpole body into sessile juveniles through metamorphosis. En, endostyle; PG, pharyngeal gill; St, stomach. (c) An adult. Body size is approximately 10 cm
Reflecting their phylogenetic position, ascidians have a gene set that specifies the chordate body with less redundancy than vertebrates (Dehal et al. 2002). Vertebrates are thought to have experienced genome duplication twice during evolution (Holland et al. 1994) and, as a consequence, they usually have several genes that are similar with respect to sequence, expression, and functions. These similar genes sometimes compensate functions for each other. Because of this redundancy of genes, disrupting a gene does not always cause strong phenotypes, and multiple paralogous genes must be analyzed to determine their functions. Genome duplication is not thought to have occurred in the ascidian lineage, and ascidians usually have one orthologous gene to their vertebrate counterparts (Sasakura et al. 2003a, b). Therefore, it is necessary to study one orthologous gene to determine such functions in ascidians. A knowledge of ascidian genes is useful in deducing functions of vertebrate orthologues that may be hidden by their redundancy. Ascidians provide an excellent experimental system because of their simple genome.
The cosmopolitan ascidian Ciona intestinalis holds a representative position among ascidians. The genome sequence of C. intestinalis was determined in 2002 (Dehal et al. 2002), and the genome has been well assembled and annotated (Satou et al. 2008). Accompanying the genome sequence, abundant expressed sequence tag (EST) and cDNA information and expression profiles of many developmentally relevant genes are available (Satou et al. 2002; Imai et al. 2006). Omic analyses of C. intestinalis have been conducted with the aid of the genome sequence, which provides gene and protein expression profiles of the germ cells (Hozumi et al. 2004; Yamada et al. 2009). Basic experimental systems analyzing genetic functions have been established in this ascidian. Forced expression of exogenous genes can be carried out by electroporation of plasmid DNAs into hundreds of embryos at once (Corbo et al. 1997). Additionally, gene functions have been disrupted by microinjection of antisense morpholino oligonucleotides (MOs) (Satou et al. 2001). The life cycle of C. intestinalis is about 2–3 months, and inland culturing systems have been developed (Joly et al. 2007). The short generation time facilitates genetic studies that are necessary for studying gene functions at later developmental stages, as discussed here (Nakatani et al. 1999; Sordino et al. 2000; Harada et al. 2008). With these characteristics, C. intestinalis is extensively studied as a splendid experimental model of ascidians.
2 Transposon-Mediated Germline Transgenesis
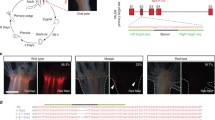
In C. intestinalis, methods of germline transformation with Tc1/mariner superfamily transposable elements have been well established (Sasakura et al. 2003c; Hozumi et al. 2013). Two transposons, Minos and Sleeping Beauty, have been used for germline transformation of C. intestinalis (Franz and Savakis 1991; Ivics et al. 1997). Transposon vectors can be modified to contain the DNA elements that a researcher wishes to introduce into the C. intestinalis genome without losing transposon activity. The transposon vectors are introduced with their transposase mRNA into C. intestinalis embryos through microinjection or electroporation (Matsuoka et al. 2005). In experimental Ciona, transposon vectors are inserted into the genome with the aid of transposase, and the transposon insertions are inherited stably by subsequent generations (Fig. 38.2). Approximately 30 % of transposon-introduced Ciona become founders (Sasakura et al. 2007). The transgenic lines are useful because they express exogenous genes in a non-mosaic fashion. The expression of exogenous genes can be controlled by selecting appropriate cis elements for the purposes of experiments. For example, the cis element of a gene expressing in a tissue-specific manner can be used to express the exogenous gene specifically in the tissue (Fig. 38.2).

Transgenic line of Ciona intestinalis. A juvenile of the transgenic line expressing green fluorescent protein (GFP) in the central nervous system (arrow in middle image). Left, a differential contrast image; middle, a fluorescence image; right, a merged image
Many kinds of exogenous genes can be utilized for molecular and cellular studies in C. intestinalis. Genes encoding fluorescent proteins are good markers for labeling cells that are alive. These fluorescent proteins can be fused to signal peptides to specifically label the organelle. Various researchers have developed multiple indicators that monitor cellular features such as calcium, voltage, apoptosis, and cell cycle (Miyawaki et al. 1997; Nakai et al. 2001; Takemoto et al. 2003; Tsutsui et al. 2008; Sakaue-Sawano et al. 2008). Transgenic lines are valuable resources for analyzing the cellular features of C. intestinalis. C. intestinalis transgenic lines are databased, and the information is available on the C. intestinalis Transgenic line RESources (CITRES) website (http://marinebio.nbrp.jp/ciona/). Transgenic lines are provided to researchers upon request (Sasakura et al. 2009).
3 Enhancer Detection
Enhancers are DNA elements that control the spatial and temporal expression patterns of genes. Enhancers are usually flexible in terms of their location and orientation with respect to the genes they regulate. When a transposon vector with a reporter gene is inserted near enhancer elements, the expression pattern of the reporter gene is altered by the enhancers (O’Kane and Gehring 1987). This phenomenon is called “enhancer detection” or “enhancer trap.”
In C. intestinalis, both Minos and Sleeping Beauty cause enhancer detection (Awazu et al. 2004; Hozumi et al. 2013). Large-scale enhancer detection has been conducted with Minos (Sasakura et al. 2008; Hozumi et al. 2010). Through enhancer detection, many marker transgenic lines that express fluorescent proteins in a tissue- or organ-specific manner have been created. Because the C. intestinalis genome is compact and on average one gene per 10-kilobase pair is located, enhancers are also densely present in the genome; this results in high frequency enhancer detection in this organism. With the aid of fluorescent markers of the enhancer detection lines, a detailed characterization of tissues and organs has been achieved in Ciona (Ohta et al. 2010). For example, the collection of enhancer detection lines that emit green fluorescent protein (GFP) in the digestive organs revealed a novel subdivision of the digestive tract (Yoshida and Sasakura 2012).
4 Insertional Mutagenesis
Transposons have the chance to insert themselves into the genomic regions and disrupt the functions of genes. For this reason, transposons can be utilized as a tool for mutagenesis. In C. intestinalis, Minos-mediated insertional mutants have been reported. In a mutant named swimming juvenile, the gene encoding cellulose synthase is mutated (Sasakura et al. 2005). Ascidians have in common the characteristic that they produce cellulose. Ascidian cellulose is found in the mantle layer, named the tunic (Fig. 38.1a), which surrounds the body to protect it from predators. In larvaceans, cellulose is found in the ‘house,’ which acts as a filter for collecting small particles in the seawater to feed (Sagane et al. 2010). The swimming juvenile mutants of C. intestinalis have a malformed and soft tunic compared to that of wild types, suggesting that cellulose gives tunics the physical strength to withstand attack from predators. In addition to this defect, swimming juvenile mutants exhibit abnormality in their process of metamorphosis. The wild-type larvae start metamorphosis after they adhere to substrates. The swimming juvenile larvae start some metamorphic events at the trunk region without adhesion to substrates, suggesting that cellulose or cellulose synthase is responsible for the pathway triggering certain metamorphic events (Nakayama-Ishimura et al. 2009).
Reflecting the compact genome of C. intestinalis, enhancer detection lines of this organism frequently have transposon insertions inside or close to genes (Hozumi et al. 2013), which suggests that enhancer detection lines are a good resource of insertional mutants. Indeed, an insertional mutant line of Ci-Hox1, which encodes a transcription factor widely conserved among metazoans, has been isolated (Sasakura et al. 2012). Detailed analyses of the Ci-Hox1 mutant have revealed that this gene is essential for forming the atrial siphon primordia, structures in the epidermis thought to be homologous to the otic placodes in vertebrates (Manni et al. 2004; Kourakis et al. 2010).
To the best of my knowledge, no mutant has yet been reported in C. intestinalis that shows defects during fertilization. Future studies will screen mutants focusing on fertilization to isolate mutants in this step, and characterization of the mutants will reveal novel molecular and genetic mechanisms responsible for allorecognition in C. intestinalis.
5 Engineered Nuclease-Mediated Genome Editing
In C. intestinalis, one major approach for disrupting the functions of genes-of-interest is to introduce antisense MOs into embryos (Satou et al. 2001). The MOs that are designed to have a sequence complementary to that of the target genes can be bound to the RNAs of the target genes to disrupt their splicing or translation. A technical limitation of this method is that MOs are lost during development and therefore their knockdown effects do not persist to the later stages. For this reason, disruption of genes in the genome through genetic modifications, namely knockout, is desirable to observe functions of genes in the germ cells because these cells are produced at later (adult) stages.
Attempts at such a reverse-genetic approach have been made in C. intestinalis with engineered nucleases (Bibikova et al. 2003; Kawai et al. 2012). The engineered nucleases are fusions of the nuclease domain of the restriction enzyme FokI and zinc-finger DNA-binding motifs (ZFNs). We can change the target sequences of ZFNs by customizing the zinc-finger motifs through substitutions of amino-acid sequences. When the ZFNs are bound to the target DNA sequences, the FokI domain introduces double-strand breaks to the target site. The double-strand breaks are repaired through two mechanisms: homologous recombination and nonhomologous end-joining. During the latter repair system, insertions and/or deletions are introduced to the target sequence, which results in the mutation of the target genes.
In C. intestinalis, the mutation frequency of ZFNs has been estimated with the ZFNs targeting enhanced GFP genes (EGFP) of the transgenic lines (Kawai et al. 2012). When 10 ng/μl EGFP-ZFN mRNA solution was introduced into embryos of an EGFP-transgenic line created by Minos, almost all the embryos became negative in their EGFP fluorescence. Genomic analyses of the EGFP-ZFN-introduced embryos indicated that the mutation frequency of EGFP is nearly 100 %. Mutations were observed in germ cells as well as in somatic cells, and the mutations can be inherited by subsequent generations. Therefore, mutagenesis with ZFNs is feasible in C. intestinalis.
Future studies will achieve knockout of endogenous genes with engineered nucleases to characterize their functions. One difficulty with ZFNs is their complicated way of construction. Recently, another DNA-binding motif derived from the transcription activator-like (TAL) effector of a plant pathogen has been used for engineered nucleases named TALENs. The DNA-binding motifs of TAL effectors can be customized more easily than those of zinc-fingers (Christian et al. 2010; Cermak et al. 2011). Therefore, knockout of genes with TALENs will be a better choice for the easy disruption of genes-of-interest.
6 Conclusions
As stated here, C. intestinalis provides a splendid experimental system for analyzing genetic functions in the chordate body. Genetic and transgenic technologies are helpful for studying fertilization and allorecognition mechanisms in this organism, and provide a superb opportunity to obtain considerable novel knowledge in future studies.
References
Awazu S, Sasaki A, Matsuoka T, Satoh N, Sasakura Y (2004) An enhancer trap in the ascidian Ciona intestinalis identifies enhancers of its Musashi orthologous gene. Dev Biol 275:459–472
Bibikova M, Beumer K, Trautman JK, Carroll D (2003) Enhancing gene targeting with designed zinc finger nucleases. Science 300(5620):764
Cermak T, Doyle EL, Christian M, Wang L, Zhang Y, Schmidt C, Baller JA, Somia NV, Bogdanove AJ, Voytas DF (2011) Efficient design and assembly of custom TALEN and other TAL effector-based constructs for DNA targeting. Nucleic Acids Res 39(12):e82
Christian M, Cermak T, Doyle EL, Schmidt C, Zhang F, Hummel A, Bogdanove A, Voytas DF (2010) Targeting DNA double-strand breaks with TAL effector nucleases. Genetics 186:757–761
Corbo JC, Levine M, Zeller RW (1997) Characterization of a notochord-specific enhancer from the Brachyury promoter region of the ascidian, Ciona intestinalis. Development (Camb) 124:589–602
Dehal P, Satou Y, Campbell RK, Chapman J, Degnan B, De Tomaso A, Davidson B, Di Gregorio A, Gelpke M, Goodstein DM, Harafuji N, Hastings KE, Ho I, Hotta K, Huang W, Kawashima T, Lemaire P, Martinez D, Meinertzhagen IA, Necula S, Nonaka M, Putnam N, Rash S, Saiga H, Satake M, Terry A, Yamada L, Wang HG, Awazu S, Azumi K, Boore J, Branno M, Chin-Bow S, DeSantis R, Doyle S, Francino P, Keys DN, Haga S, Hayashi H, Hino K, Imai KS, Inaba K, Kano S, Kobayashi K, Kobayashi M, Lee BI, Makabe KW, Manohar C, Matassi G, Medina M, Mochizuki Y, Mount S, Morishita T, Miura S, Nakayama A, Nishizaka S, Nomoto H, Ohta F, Oishi K, Rigoutsos I, Sano M, Sasaki A, Sasakura Y, Shoguchi E, Shin-I T, Spagnuolo A, Stainier D, Suzuki MM, Tassy O, Takatori N, Tokuoka M, Yagi K, Yoshizaki F, Wada S, Zhang C, Hyatt PD, Larimer F, Detter C, Doggett N, Glavina T, Hawkins T, Richardson P, Lucas S, Kohara Y, Levine M, Satoh N, Rokhsar DS (2002) The draft genome of Ciona intestinalis: insight into chordate and vertebrate origins. Science 298(5601):2157–2167
Delsuc F, Brinkmann H, Chourrout D, Philippe H (2006) Tunicates and not cephalochordates are the closest living relatives of vertebrates. Nature (Lond) 439(7079):965–968
Franz G, Savakis CC (1991) Minos, a new transposable element from Drosophila hydei, is a member of the Tc1-like family of transposons. Nucleic Acids Res 19:6646
Harada Y, Takagaki Y, Sunagawa M, Saito T, Yamada L, Taniguchi H, Shoguchi E, Sawada H (2008) Mechanism of self-sterility in a hermaphroditic chordate. Science 320(5875):548–550
Holland PW, Garcia-Fernàndez J, WIlliams NA, Sidow A (1994) Gene duplications and the origins of vertebrate development. Dev Suppl:125–133
Hozumi A, Satouh Y, Ishibe D, Kaizu M, Konno A, Ushimaru Y, Toda T, Inaba K (2004) Local database and the search program for proteomic analysis of sperm proteins in the ascidian Ciona intestinalis. Biochem Biophys Res Commun 319:1241–1246
Hozumi A, Kawai N, Yoshida R, Ogura Y, Ohta N, Satake H, Satoh N, Sasakura Y (2010) Efficient transposition of a single Minos transposon copy in the genome of the ascidian Ciona intestinalis with a transgenic line expressing transposase in eggs. Dev Dyn 239:1076–1088
Hozumi A, Mita K, Miskey C, Mates L, Izsvak Z, Ivics Z, Satake H, Sasakura Y (2013) Germline transgenesis of the chordate Ciona intestinalis with hyperactive variants of sleeping beauty transposable element. Dev Dyn 242:30–43
Imai KS, Levine M, Satoh N, Satou Y (2006) Regulatory blueprint for a chordate embryo. Science 312:1183–1187
Ivics Z, Hackett PB, Plasterk RH, Izsvak Z (1997) Molecular reconstruction of Sleeping Beauty, a Tc1-like transposon from fish, and its transposition in human cells. Cell 91:501–510
Joly JS, Kano S, Matsuoka T, Auger H, Hirayama K, Satoh N, Awazu S, Legendre L, Sasakura Y (2007) Culture of Ciona intestinalis in closed systems. Dev Dyn 236:1832–1840
Kawai N, Ochiai H, Sakuma T, Yamada L, Sawada H, Yamamoto T, Sasakura Y (2012) Efficient targeted mutagenesis of the chordate Ciona intestinalis genome with zinc-finger nucleases. Dev Growth Differ 54:535–545
Kourakis MJ, Newman-Smith E, Smith WC (2010) Key steps in the morphogenesis of a cranial placode in an invertebrate chordate, the tunicate Ciona savignyi. Dev Biol 340:134–144
Manni L, Lane N, Joly J, Gasparini F, Tiozzo S, Caicci F, Zaniolo G, Burighel P (2004) Neurogenic and non-neurogenic placodes in ascidians. J Exp Zool B Mol Dev Evol 302:483–504
Matsuoka T, Awazu S, Shoguchi E, Satoh N, Sasakura Y (2005) Germline transgenesis of the ascidian Ciona intestinalis by electroporation. Genesis 41:61–72
Miyawaki A, Llopis J, Heim R, McCaffery JM, Adams JA, Ikura M, Tsien RY (1997) Fluorescent indicators for Ca2+ based on green fluorescent proteins and calmodulin. Nature (Lond) 388:882–887
Nakai J, Ohkura M, Imoto K (2001) A high signal-to-noise Ca(2+) probe composed of a single green fluorescent protein. Nat Biotechnol 19:137–141
Nakatani Y, Moody R, Smith WC (1999) Mutations affecting tail and notochord development in the ascidian Ciona savignyi. Development 126:3293–3301
Nakayama-Ishimura A, Chambon JP, Horie T, Satoh N, Sasakura Y (2009) Delineating metamorphic pathways in the ascidian Ciona intestinalis. Dev Biol 326:357–367
Ohta N, Horie T, Satoh N, Sasakura Y (2010) Transposon-mediated enhancer detection reveals the location, morphology and development of the cupular organs, which are putative hydrodynamic sensors, in the ascidian Ciona intestinalis. Zool Sci 27:842–850
O’Kane C, Gehring WJ (1987) Detection in situ of genomic regulatory elements in Drosophila. Proc Natl Acad Sci USA 84:9123–9127
Sagane Y, Zech K, Bouquet JM, Schmid M, Bal U, Thompson EM (2010) Functional specialization of cellulose synthase genes of prokaryotic origin in chordate larvaceans. Development (Camb) 137:1483–1492
Sakaue-Sawano A, Kurokawa H, Morimura T, Hanyu A, Hama H, Osawa H, Kashiwagi S, Fukami K, Miyata T, Miyoshi H, Imamura T, Ogawa M, Masai H, Miyawaki A (2008) Visualizing spatiotemporal dynamics of multicellular cell-cycle progression. Cell 132:487–498
Sasakura Y, Yamada L, Takatori N, Satou Y, Satoh N (2003a) A genomewide survey of developmentally relevant genes in Ciona intestinalis. VII. Molecules involved in the regulation of cell polarity and actin dynamics. Dev Genes Evol 213:273–283
Sasakura Y, Shoguchi E, Takatori N, Wada S, Meinertzhagen IA, Satou Y, Satoh N (2003b) A genomewide survey of developmentally relevant genes in Ciona intestinalis. X. Genes for cell junctions and extracellular matrix. Dev Genes Evol 213:303–313
Sasakura Y, Awazu S, Chiba S, Satoh N (2003c) Germ-line transgenesis of the Tc1/mariner superfamily transposon Minos in Ciona intestinalis. Proc Natl Acad Sci USA 100(13):7726–7730
Sasakura Y, Nakashima K, Awazu S, Matsuoka T, Nakayama A, Azuma J, Satoh N (2005) Transposon-mediated insertional mutagenesis revealed the functions of animal cellulose synthase in the ascidian Ciona intestinalis. Proc Natl Acad Sci USA 102:15134–15139
Sasakura Y, Oogai Y, Matsuoka T, Satoh N, Awazu S (2007) Transposon-mediated transgenesis in a marine invertebrate chordate, Ciona intestinalis. Genome Biol 8(suppl 1):S3
Sasakura Y, Konno A, Mizuno K, Satoh N, Inaba K (2008) Enhancer detection in the ascidian Ciona intestinalis with transposase-expressing lines of Minos. Dev Dyn 237:39–50
Sasakura Y, Inaba K, Satoh N, Kondo M, Akasaka K (2009) Ciona intestinalis and Oxycomanthus japonicus, representatives of marine invertebrates. Exp Anim 58:459–469
Sasakura Y, Kanda M, Ikeda T, Horie T, Kawai N, Ogura Y, Yoshida R, Hozumi A, Satoh N, Fujiwara S (2012) Retinoic acid-driven Hox1 is required in the epidermis for forming the otic/atrial placodes during ascidian metamorphosis. Development (Camb) 139:2156–2160
Satoh N (1994) Developmental biology of ascidians. Cambridge University Press, New York
Satoh N (2003) The ascidian tadpole larva: comparative molecular development and genomics. Nat Rev Genet 4:285–295
Satou Y, Imai KS, Satoh N (2001) Action of morpholinos in Ciona embryos. Genesis 30:103–106
Satou Y, Yamada L, Mochizuki Y, Takatori N, Kawashima T, Sasaki A, Hamaguchi M, Awazu S, Yagi K, Sasakura Y, Nakayama A, Ishikawa H, Inaba K, Satoh N (2002) A cDNA resource from the basal chordate Ciona intestinalis. Genesis 33:153–154
Satou Y, Mineta K, Ogasawara M, Sasakura Y, Shoguchi E, Ueno K, Yamada L, Matsumoto J, Wasserscheid J, Dewar K, Wiley GB, Macmil SL, Roe BA, Zeller RW, Hastings KE, Lemaire P, Lindquist E, Endo T, Hotta K, Inaba K (2008) Improved genome assembly and evidence-based global gene model set for the chordate Ciona intestinalis: new insight into intron and operon populations. Genome Biol 9:R152
Sordino P, Heisenberg CP, Cirino P, Toscano A, Giuliano P, Marino R, Pinto MR, De Santis R (2000) A mutational approach to the study of development of the protochordate Ciona intestinalis (Tunicata, Chordata). Sarsia 85:173–176
Takemoto K, Nagai T, Miyawaki A, Miura M (2003) Spatio-temporal activation of caspase revealed by indicator that is insensitive to environmental effects. J Cell Biol 160:235–243
Tsutsui H, Karasawa S, Okamura Y, Miyawaki A (2008) Improving membrane voltage measurements using FRET with new fluorescent proteins. Nat Methods 5:683–685
Yamada L, Saito T, Taniguchi H, Sawada H, Harada Y (2009) Comprehensive egg-coat proteome of an ascidian Ciona intestinalis reveals gamete recognition molecules involved in self-sterility. J Biol Chem 284:9402–9410
Yoshida R, Sasakura Y (2012) Establishment of enhancer detection lines expressing GFP in the gut of the ascidian Ciona intestinalis. Zool Sci 29:11–20
Acknowledgments
The author thanks the members of the Shimoda Marine Research Center at the University of Tsukuba for their kind cooperation during our study. I am also grateful to the National Bioresource Project, MEXT, Dr. Nobuo Yamaguchi, Dr. Kunifumi Tagawa, and Dr. Shigeki Fujiwara and his colleagues for providing me with Ciona adults.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
This chapter is published under an open access license. Please check the 'Copyright Information' section either on this page or in the PDF for details of this license and what re-use is permitted. If your intended use exceeds what is permitted by the license or if you are unable to locate the licence and re-use information, please contact the Rights and Permissions team.
Copyright information
© 2014 The Author(s)
About this paper
Cite this paper
Sasakura, Y. (2014). Germline Transformation in the Ascidian Ciona intestinalis . In: Sawada, H., Inoue, N., Iwano, M. (eds) Sexual Reproduction in Animals and Plants. Springer, Tokyo. https://doi.org/10.1007/978-4-431-54589-7_38
Download citation
DOI: https://doi.org/10.1007/978-4-431-54589-7_38
Published:
Publisher Name: Springer, Tokyo
Print ISBN: 978-4-431-54588-0
Online ISBN: 978-4-431-54589-7
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)