
Abstract
Fall Armyworm (FAW), Spodoptera frugiperda (Lepidoptera: Noctuidae), native to the Americas, is a polyphagous insect pest feeding on more than 350 plant species. We studied the developmental and demographic parameters of the maize (Zea mays) strain of FAW on rice (Oryza sativa), and compared the results with its prime host, maize. The developmental period from egg to adult among rice varieties did not differ significantly; however, it did differ significantly between rice and maize, as feeding on rice rather than maize extends development duration of FAW larvae by 15.15%. FAW larvae collected and reared on maize were found to be of significantly higher weight than those reared on rice at two sequential dates of their development; pupal weight however was observed as statistically similar between these two host crops. Regardless of the host, female adults always emerged before males; in maize, female FAW appeared 3.36 days earlier than males. Females derived from rice had longer pre-oviposition periods and shorter oviposition ones than those derived from maize. In rice and maize, the age-specific fecundity rate (mx) peaked at 40 days and 33 days, respectively. When the Fall Armyworm consumed maize instead of rice, there was an increase in the reproduction rate (R0), the intrinsic rate of natural increase (rm), and the finite rate of increase (λ). For instance, when FAW fed on rice, the rm value was 0.121, whereas it rose to 0.173 when FAW fed on maize. Feeding on rice instead of maize resulted in significantly longer mean length of generation (tG) and doubling time (tD) for the fall armyworm (FAW). This suggests that it took a longer time for the FAW population to double when it was fed rice under controlled greenhouse conditions. In summary, our research suggests that FAW can survive and complete its life cycle on rice plants and on multiple varieties of rice in Bangladesh. However, field verification is necessary before drawing strong conclusions as to the risk posed by FAW in rice. This requires additional studies of FAW and associated insect community dynamics under non-controlled conditions and in the context of multi-species interactions in Asian rice fields.
Similar content being viewed by others


Introduction
Fall Armyworm (FAW), Spodoptera frugiperda (J. E. Smith, 1797) (Lepidoptera: Noctuidae) is one of the most important global invasive pests. Native to the Americas, it was first documented in West Africa in early 2016 (Goergen et al. 2016) (CABI 2020; Early et al. 2018), posing a significant threat to food security and agricultural sustainability (Rwomushana et al. 2018). After first invading West Africa, FAW quickly invaded most African countries after which it spread eastward to India, Myanmar, Thailand, China, Bangladesh, and other countries (Early et al. 2018; CABI 2017; FAO 2018; FAO 2020; Ali et al. 2021; Nakweta 2018; Alam et al. 2018; Yang et al. 2019; Sun et al. 2021).
Its severity is due to its polyphagous nature, feeding on the leaves, stems, fruits, and cobs of more than 350 plant species including maize, rice, millet, wheat, sugarcane, many pasture grasses and weeds (Montewzano et al. 2018; Overton et al. 2021). There are two host strains of Fall Armyworm: the rice strain and the maize (Zea mays) strain. Rice (Oryza sativa) is the main food for the population of the countries invaded by FAW after 2016, including India, China, Myanmar, Thailand and Bangladesh, making it a potentially serious threat to food security in rice-growing countries. Exploring the response of FAW to rice could be valuable for the development of integrated pest management (IPM) strategies of this invasive pest. To help improve IPM of FAW, the incorporation of host plant resistance is desireable, although efforts to achieve native geneic resistance remain early-stage, due in part to limited studies screening available germplasm and cultivars (Farias et al. 2014; Abrahams et al. 2017; Kumela et al. 2018). Like other countries, Bangladesh currently lacks commercially available rice cultivars that possess resistance to FAW. Therefore, it is crucial to examine a variety of genetic resources and identify rice genotypes that could be incorporated into IPM programs and efforts to breed for FAW resistance. By evaluating diverse rice genotypes, including those already cultivated by farmers, superior materials could potentially pinpointed from a broader range of genetic resources to enhance specific desirable traits.
Alam et al. (2018) first reported the occurrence of FAW in maize fields in Bangladesh using pheromone trapping and confirmed the arrival of the pest through identification of its morphological features. Later, FAW larvae were collected from different maize fields and characterized using molecular approaches; this indicated that the immigrant population observed in Bangladesh are mixed in their strain types (both maize and rice are present, unpublished data). In Bangladesh, FAW generally attacks maize and is observed in all over the country. FAW has also been observed to attack rice plants (Sartiami et al. 2020); yet there is no robust information on damage by this pest to paddy in Bangladesh. Rice is the staple food of Bangladesh and is widely cultivated in three seasons throughout the year. More than 100 rice varieties are commercially cultivated in Bangladesh, and a larger number of landraces are present. Testing rice varieties for their potential resistance to FAW is however an untapped area of research important in the context of pre-emptive pest control and IPM programs.
A clear understanding of the basic biological, developmental and life-history parameters of FAW is prerequisite to developing effective and sustainable IPM tactics. This is because the survival, longevity, fecundity, body size and population growth of herbivorous insects are affected significantly by host plant types (Heinrichs et al. 2017; Luginbill 1982; Maia et al. 2000; Pantoja et al. 1986; Awmack and Leather 2002). Demographic studies could be helpful in determining the predominant age distribution and mortality rate in natural populations, with the intrinsic rate of increase (rm) as a key demographic parameter necesssary for predicting potential establishment and population growth (Mehrkhou et al. 2012; Southwood et al. 2000). Such data also help to evaluate host suitability and damage potential to cultivated varieties.
Although FAW has a wide number of plant hosts, only a limited group of host plants have been thoroughly examined to quantify their effect on FAW development, reproduction, and demographic parameters. All of FAW’s demographic parameters – including generation time (tG), intrinsic rate of increase (rm), finite rate of increase (λ) and net reproductive rate (R0) have – varied significantly by host plant species in a range of studies (Li-Hong et al. 2021; Chen et al. 2020; Hutasoti et al. 2020; Montezano et al. 2019). Li-Hong et al. (2020) found the rm value of FAW to be similar in maize and in tomato, but lower when FAW fed on pepper. This underscores the ways in which demographic parameters can change significantly depending on which host plant experiences herbivory (Fathipour et al. 2011, 2020; Mehrkhou et al. 2012). In this study,we test the hypothesis that when compared to maize, rice will differentially affect the demographic parameters of FAW and influence the pest's developmental period from egg to adult, thereby affecting overall survivability.
Prior studies have shown that developmental periods are prolonged when FAW larvae are reared on cotton and soybean rather than maize, with a negative impact on pupal weight (Chen et al. 2020; Silva et al. 2017). He et al. (2021a) studied the effect of soybean, oilseed rape and sunflower on the development and lifetable parameters of FAW and found them to be significantly impacted by each host. Unfortunately, only a limited number of reports are available which address the developmental and demographic parameters of FAW using rice as a host. Given the unprecidented insursion of FAW into predominantly rice growing areas in Asia since 2018, an understanding of this information is important for developing more effective management approaches. A very limited 0.4% survival rate for FAW larvae has been documented in earlier studies examining feeding on rice as a host, as FAW larvae were unable to pupate successfully (Devi et al. 2018; Harrison et al. 2019; He et al. 2021b). Yet given the limited number of studies and in light of the recent invasion of FAW into Asia, a more thorough investigation into the effects of rice as a host plant on the development, survival, reproduction and demographic parameters of FAW could be useful. In response, we studied the effect of rice on the development and demographic parameters of FAW compared with maize as a more suitable host. We also describe the development of a novel method for the study of lifecycles and varietal screening that can be replicated in other settings. This research thus provides insights on the host feeding preferences and biological ramifications of FAW in the context of rice-based farming systems that can help to inform more comprehensive IPM strategies in Asia and other regions.
Materials and methods
Stock culture of FAW and rearing in the greenhouse
FAW larvae identified as the Zea mays feeding strain were collected from an infested maize field in Alamdanga, Chuadanga district, Bangladesh (23°73'50N'', 88°91'31'E). The larvae were subsequently raised on maize plants within the greenhouse of the Entomology Division at the Bangladesh Rice Research Institute (BRRI) in Gazipur (23°59'27"N 90°24'28"E), Bangladesh. Larvae were monitored daily, and their food was replenished as needed. After emergence, the male and female fall armyworms were released into plastic jars. Brown paper from a paper bag was placed inside the jars to provide a suitable surface for oviposition. The open top of each jar was covered with a fine nylon mesh net, which allowed ventilation while also preventing moths from escaping. To sustain larval health, a food source consisting of a twenty percent sugar solution was provided. Cotton material was used to provide a substrate for the sugar solution. Surprisingly, the moths displayed a preference for laying eggs on the mesh net rather than on the brown paper, although eggs were found on both surfaces. Eggs were then collected from both the net and the paper surface for subsequent research. Through molecular and phenotypic studies, the collected sample was confirmed to be of the the maize strain of FAW.
We also placed eggs on the leaves of maize plants in the greenhouse. Once the eggs hatched, the larvae began feeding on the maize leaves and underwent developmental to become adult moths. These female moths subsequently laid eggs on the leaves of the maize plants. Through these processes, we successfully established a stock culture of FAW in our laboratory, that was maintained using maize plants as hosts within the greenhouse. As soon as an ample number of adult moths and larvae were available, we utilized them for the experimental studies described below.
Collection of rice varieties
Fifteen rice varieties were used for this study,Footnote 1 with the seeds collected from the Genetic Resources and Seed Division and Plant Breeding Division of BRRI, Gazipur, Bangladesh. To date, BRRI has developed 113 rice varieties which are estimated to cover more than 80% of the rice production areas of Bangladesh. Most of these varieties are however susceptible to other rice insect pests including plant hoppers, leafhoppers, stem borers, rice hispa, leaffolder and rice bug. Among the 113 varieties available on a commercial basis in Bangladesh, 15 varieties are randomly selected for testing against FAW.
The impact of different rice varieties on the development of the Fall Armyworm (FAW)
Two different methods, namely the potted plant method and the tray seedling method, were employed in experiments to assess the development of FAW when feeding on rice. To prepare the medium for the experiments, soil was collected from a rice field located at BRRI. The collected soil was pulverized to eliminate inert materials, plant roots, visible insect pests, and plant propagules, ensuring a clean and controlled environment for the study. After pulverization, the soil was sun-dried and then thoroughly mixed to ensure uniformity. For the experiment, individual plastic pots measuring 13.97 cm in diameter and 12.7 cm in height were filled with 2.5 kg of the dry soil. To prepare the soil for sowingan adequate amount of water was added to make it muddy one day prior to crop establishment. The soil in the pots was fertilized following a standard regime based on the guidelines provided by BRRI in 2017 (BRRI 2017). In each pot and tray, a total of 200 pre-germinated seeds of each of the 15 varieties were sown onto the wet mud (Fig. 1). Prior to sowing, the seeds underwent a specific preparation process: they were soaked in water for 24 h, drained, and then incubated. After another 48 h of soaking, the seeds were sown once they had sprouted and when the radicle had grown to a length of 2–3 mm. After sowing germinated seeds, water was splashed onto them and they were lightly pressed down by hand to improve soil to seed contact. To maintain the appropriate moisture levels, the seed pots were initially kept saturated with water for the first five days. As the seedlings began to grow, the water level was gradually increased to reach a depth of 5 cm. For each rice variety, 10 pots and three separate trays were utilized for seeding. The trays were filled with the same prepared soil that was used in the pots, ensuring consistency across the experimental setup.

Young seedlings emerged in pots and in galvanized stee; trays. Seeds were sown on 25 December, 2020
For FAW establishment, thirty-day-old rice seedlings that were grown in earthen pots were selected (Fig. 2). In each pot, ten first-instar neonate larvae were introduced using a fine camel hair brush. To confine the larvae within each pot, mylar polyester film cages (Fig. 3) were utilized. The emerging adults from the infested pots were recorded and subsequently used for fecundity experiments. In this experiment, five pots were used for each rice variety, with each pot serving as one replication. The experimental setup followed a completely randomized design, with pots rotated in location daily to avoid any microclimate variability within the greenhouse. Rice seedlings of the variety BRRI dhan49 and maize seedlings of the hybrid corn PAC-339 (from Lal Teer Seed Ltd, Bangladesh) were conversely cultivated in galvanized steel trays. The management practices employed were identical to those used in the pot experiment (as depicted in Fig. 4, left). For the experiment, seedlings aged between 25 and 30 days were selected as recommended as the age for transplanting by BRRI.

Above: Young rice seedlings in pots, ready for FAW larvae infestation. Below: Maize used as control plants for experiments

Potted rice seedlings, that were treated with 10 neonate first-instar FAW larvae and covered with a mylar cage. The open top of the mylar cage was covered with fine mesh netting which ensured proper ventilation and prevented the hatched moths from escaping.

Above (Left): Rice seedlings ready in a galvanized steel tray with 100 first-instar FAW larvae to be released in each tray. (Right) Infested tray covered with fine mesh net supported by an iron frame
To initiate experimental feeding, fifty one-day-old neonate larvae were released onto the rice seedlings grown in the trays. This was accomplished using a fine camel hair brush to ensure precise placement of the larvae on the seedlings. To protect the trays from feeding by other insects and to prevent FAW escape, a nylon mesh netting was utilized to cover the tray. Additionally, an iron cage was employed as support for the netting as shown in Fig. 4 (right).
Larval development was visually monitored throughout their growth stages, starting from the first instar until adulthood. This allowed for a comprehensive assessment of their life cycle on the rice host. Similarly, for the comparison with the rice host, 50 first-day neonate larvae were released onto maize seedlings using methods as described above for rice. The growth and development of the larvae on maize were observed and compared to their growth on the rice host. To calculate larval survival rates, data regarding the emergence of adult moths was collected and analyzed. This provided insights into the percentage of larvae that successfully reached adulthood.
In a separate experiment conducted using the tray seedling method, we examined the impact of the host on FAW larval growth. In this experiment, we released 100 first-day neonate larvae onto both maize and rice seedlings that were grown in a tray. The growth rates of the larvae were then compared on two specific dates. All of these experiments were conducted within a controlled greenhouse environment, maintained at a temperature of 27 °C ± 2 °C and a relative humidity of 65% ± 5% dur the duration of the experiment.
To gather data on larval and pupal development, we recorded the weights of 7-day-old and 13-day-old larvae, as well as the weights of the pupal stages. An analytical balance (KERN ABJ220-4NM, Germany) was used for weighing, with a maximum capacity of 200 g, minimum readability of 10 mg, and a precision of 0.1 mg. On each of the two specific dates, we measured the weights of 30 larvae and pupae from the rice host for each variety tested, as well as 30 larvae and pupae (2-day-old) from the maize host. This allowed for a comprehensive comparison of the larval and pupal weights between the two hosts.
Adult longevity, oviposition period and fecundity
The population and reproduction parameters of FAW on rice and maize were determined using the methods described by Birch (1948) and Carey (1993, 2001). These methodologies provide a framework for calculating various parameters related to population dynamics and reproduction. By applying these models and methodologies to the population data obtained from FAW on rice and maize, we can calculated and compared their respective populations and reproduction parameters. Each transparent plastic container (11 cm in diameter × 12 cm in height) was filled with a combination of female and male moths for this experiment (25–35 replications). The opening was covered with fine nylon mesh for ventilation and to keep the moths from escape. To provide a supply of carbohydrates for adult feeding, a smallcotton wick soaked in a solution of 20 percent sugar was placed inside the containers. Each of the eggs layed by female moths were counted daily until the ovipositing moth died. To determine the percentage of egg hatchability, 10 egg masses were collected from each replicate. Egg masses were then placed in a plastic box within a controlled greenhouse environment at a temperature of 27 °C ± 2 °C and a relative humidity of 65% ± 5%. Eggs were monitored daily until they hatched. The time taken for the eggs to hatch and the percentage of hatched eggs were recorded, the latter calculated as follows:
Effect of host plants on the demographic parameters of Fall Armyworm
In the process of calculating life-history parameters, it is important to take into account any mortality that occurs during the oviposition and post-oviposition period. Some adult females may have died shortly after release into the plastic jar, while others could have been accidentally killed during transfer and inspection of the experimental unit within two or three days following release. The number of females that died or were accidentally killed should be recorded separately and excluded from the calculations for these specific parameters. These individuals were therefore excluded from subsequent analysis. Life tables were generated for each host using age-specific mortality (lx) and fecundity (mx) rates on a daily basis. The intrinsic rate of natural increase (rm) was calculated from the life–fecundity table, according to the equation [∑e−rmxlxmx = 1] provided by Birch (1948), where x is the female FAW age in days, lx is the age-specific survival rate [(fraction of females surviving at age x) × (rate of egg hatchability) × (survival rate of immature stages)] and mx is the expected number of daughters produced per female alive at age x [(age-specific oviposition) × (proportion of females)] (Sabelis 1985; Gotoh and Gomi 2003). The net reproductive rate (R0) is given by [R0 = ∑lxmx], while the mean generation time (tG) in days is given by [tG = ln R0/rm]. The finite rate of increase (λ) is given by [λ = erm], and the doubling time (tD) in days is [tD = (ln 2)/rm].
After calculating rm (ri) from the original data (rall), the standard errors for the life-history parameters at different constant temperatures were estimated using the jackknife method. This involves systematically omitting one insect at a time from the dataset and recalculating rm (ri) based on the remaining insects (n − 1). Based on the approach described by Meyer et al. (1986), the jackknife pseudo-value (rj) was computed for each subset of the original data using the equation: rj = n rall − (n − 1)ri. This process was repeated for all possible omissions of one insect from the original dataset, generating a set of pseudo-values. By using these pseudo-values, confidence limits for the parameter values were computed, allowing for a more accurate estimation of the standard errors associated with the life-history parameters. The jackknife method provides a robust approach for assessing the variability and uncertainty in the estimated parameter values at different constant temperatures (Kavallieratos et al. 2020), and was therefore applied in our study.
Statistical analysis
The data related to the immature developmental period (egg-adult), larval growth rate, adult longevity, pre-oviposition period, oviposition period, post-oviposition period, age-specific fecundity, and age-specific daily egg production were subjected to statistical analysis using R and SPSS software. For the analysis of the immature developmental period, one-way analysis of variance (ANOVA) was performed, with rice varieties serving as the explanatory variables. Welch t-tests were used to compare FAW demographic parameters between maize and rice hosts. Prior to conducting the statistical analysis, the response variable data were transformed using a logarithmic or arcsine function. This transformation was done to stabilize the variance of the data and ensure that the back-transformed values remained non-negative. By applying appropriate transformations to the data, any issues related to unequal variances were addressed, allowing for valid statistical analysis and interpretation of the results.
Results
Effect of rice varieties and maize on the development of Fall Armyworm
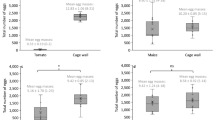
Figure 5 displays the duration of the developmental period from egg to adult when FAW larvae were fed different rice varieties in the pot experiment. The results of the statistical analysis indicated that rice varieties had no significant impact on the development period (F = 1.349, df = 69, P = 0.2108). Based on these findings, we decided to use only the seedling tray method for further studies, focusing on both rice and maize as host plants. The mean duration of the immature developmental period (from larva to adult) significantly differed between the rice host (31.75 ± 1.65 days) and the maize host (26.94 ± 0.72 days) (t = 11.997, df = 6, P < 0.01). This indicates that FAW development from larva to adult is delayed by approximately 15.15% days when feeding on rice compared to maize. Further analysis revealed that the development time from egg to adult did not significantly differ between males and females within the same host (Table 1). Additionally, the host plant (rice or maize) did not have a significant impact on the survival rate (t = 0.172, df = 6, P = 0.869).

Developmental time (egg to adult) of FAW fed on different rice varieties (earthen pot experiment). NS indicates non-significant difference at five percent level of significance. Error bar represents standard error
The data in Figure 6 demonstrate that larvae fed on maize exhibited significantly higher larval weight compared to those fed on rice at both 7 days (df = 31, P < 0.001) and 13 days (df = 42, P < 0.001) of growth. However, when considering FAW pupae, no statistically significant difference in weight was observed between the two hosts (F = 2.0385, df = 18, df = 21, P = 0.1193). This suggests that FAW achieved similar weight during the pupal stage regardless of the host. From these findings, we can infer that the host plant mainly influences the growth rate per day rather than the final body weight of the FAW, which is critical for their adulthood. Notably, female adults emerged earlier than the male adults, regardless of the host (Table 1). This distinction was particularly pronounced in maize, where female adults emerged approximately 3.36 days earlier than male adults. These results highlight the impact of the host plant on larval weight and the timing of adult emergence in FAW, suggesting that maize facilitates higher larval weight and exhibits a more pronounced difference in emergence timing between male and female adults.

Effect of host plant on the body weight of FAW larvae. Weight was measured at 7-day old larval, and 13-day old larval and pupal stage (n = 30). Error bars indicate standard error of the mean. **indicates significant differences at P < 0.01. NS = non-significant at the 5% level of significance
Reproduction
Host plant specicies had a significant impact on the pre-oviposition, oviposition, and post-oviposition periods, as well as the fecundity per FAW female, as shown in Table 2. Females that developed on maize exhibited a shorter pre-oviposition period and a longer oviposition period compared to females that developed on rice. Additionally, females developing on maize had a significantly longer post-oviposition period and higher fecundity compared to those developed on rice. However, the daily egg production (eggs per female per day) did not differ significantly between maize and rice (Table 2, F = 1.4501, df = 27, P = 0.3461). Although there are differences in the pre-oviposition, oviposition, and post-oviposition periods, this observation suggests that daily egg production remained similar between the two hosts.
Demographic parameters
The proportion of female offspring and the survival rate of immature stages were analyzed to determine demographic parameters. The age-specific survival rate (lx) showed an earlier decline in maize compared to rice (Fig. 7). Significantly higher proportions of female individuals were observed when the larvae were fed on rice compared to maize (Table 2, F = 0.3309, df = 26, P = 0.0046). Female adults started to die on days 39 and 33, on rice and maize, respectively (Fig. 7). Oviposition began within two to nine days after adult emergence, with females feeding on rice exhibiting a shorter oviposition period compared to those feeding on maize (Table 2). The age-specific fecundity rate (mx) peaked at 40 days in rice and 33 days in maize (Fig. 7).

Age-specific survival (lx) and fecundity (mx) rates of female Fall Armyworm on rice and maize hosts
The intrinsic rate of natural increase (rm), reproductive rate (R0), and finite rate of increase (λ) were significantly higher when FAW larvae were fed on maize compared to rice (Table 3; t = 175.97, df = 52, P < 0.001 for rm, t = 156.71, df = 35, P < 0.001 for R0, and t = 7.7061, df = 52, P < 0.001 for λ). The average length of a generation time (tG) was significantly lower when females fed on maize compared to rice (t = − 2.4712, df = 45, P = 0.0173). Similarly, a significantly lower doubling time (tD) was observed when females were fed on maize (Table 3, t = − 9.0012, df = 39, P < 0.001). These results indicate that FAW takes a longer time to double its population when fed on rice compared to maize.
Discussion and conclusions
The development of the maize strain of FAW and its biological parameters have been studied on many host plants including maize, sorghum, wheat, soybean, peanut and cotton; however, rice as a host plant has not yet been a major focus of comprehensive research. Silva et al. (2021) tested the effect of different rice genotypes on the biology of FAW and found that different varieties had a significant impact of FAW development. This study however was confined to the assessment of biological parameters of FAW on just four rice varieties. Conversely, our study includes the assessment of demographic indices while also testing a new methodology in the form of tray-based screening. This method appears to hold promise for both the study of FAW herbivory and as a complement for varietal screening programs. More conventional methods have also included the study of herbivory and insect development using petridishes or tubes. These however entail high transactions costs as food and water must be resupplied on a more regular basis than in the current tray-based study.
We also used rice as a host compared to maize as a control for our study of FAW demographics, as rice is the staple food for more than half of the global population and is the primary crop grown in the areas of Asia invaded by FAW since 2018. Our results showed that both sexes of FAW required longer developmental times when fed on rice compared to maize (Table 1), possibly due to variation in the nutritional quality and palatability of rice. Quality of food is among the most important factors for insect growth and development (Barros et al. 2010), with insect performance greatly affected by the type and quality of host plant organs (Koussoroplis and Wacker 2016; McCormick et al. 2019). The immature stage developmental period (26.94 days) of maize-fed FAW observed in our study was similar to that reported in discovered by previous studies, for example, the developmental time of FAW from egg to adult being 25 days (Chen et al. 2020) and 26.4 days (Sharanabasappa et al. 2018).
Our results also suggested that FAW larvae reared on maize performed better than those reared on rice. This is perhaps unsurprising, as maize is the prime host of FAW, with numerous studies documenting developmental performance of FAW on maize compared to alternative hosts (Li-Hong et al. 2021; Wang et al. 2020; Montezano et al. 2019; Muria and Virla 2004). Buntin (1986) and Nagoshi et al. (2007), for example, reported that FAW larvae fed on maize performed better than those fed on cotton or soybean. Similarly, FAW larvae fed on these alternative crops had longer larval duration and reduced pupal weight than larvae fed on maize (Chen et al. 2020). Our results also showed that a rice diet had an impact on larval growth rate (Fig. 2). Significantly higher larval growth rate was observed at 7 and 13 days when fed on maize than when fed on rice. Immature survival rates were higher (50.0%) and larvae developed more quickly (4.81 days earlier) when reared on maize than rice (43.0%). Host quality and nutrients obtained from the host plant might cause these observed differences , which are commonly found in other insects when they are reared on different host plants (Silva et al. 2017; Day et al. 2017; Groote et al. 2016). Chen et al. (2020) reported that FAW larvae gained different body weights according to the host plant they fed on, while Silva et al. (2017) reported that when FAW were reared on cotton and soybean rather than maize had extended larval duration and reduced body weight. They also found reduced pupal body weight when fed on plants other than maize. In contrast however, our result shows statistically similar pupal weight when fed on rice and maize, a result dissimilar to other reports. Silva et al. (2017) also reported that compared to maize, FAW larvae fed on soybean and cotton expereinced reduced pupal weight. To some extent, the pupal stage's status reflects the prior larvae’s ability to adapt to feed on a particular host and in their environment (Soto et al. 2018). FAW reared on tomato had a considerably greater pupal weight than FAW fed on maize or pepper in prior research, indicating that tomato-fed FAW larvae may have experienced improved nutrition on that host (Li-Hong et al. 2021).
We also found that host type plant affects the fecundity and longevity of FAW. Female FAW fed on rice plants in our study laid a significantly lower number of eggs than those fed on maize. They and also had a shorter lifespan, indicating that type of host plant has an important influenvce on the fecundity and adult lifespan of FAW, as also reported by Barros in 2010 (Barros et al. 2010). Females reared on cotton had a slightly shorter lifespan than females reared on maize and soybean, (Barros et al. 2010). In the present study, however, the fecundity of FAW reared on maize was similar to that reported by Sharanabasappa et al. (2018). Moreover, the number of eggs per female produced by cotton-fed FAW has been reported to be significantly lower than the number of eggs per female when fed on maize (Chen et al. 2020). Again, these results indicate that host plants can have considerable influence on the FAW reproduction rate, although field studies in the presence of multiple species and potential predatory and parasitic organisms are needed to quantify the degree to which host plant type may affect reproduction and development under on-farm conditions.
Within a given set of climate and food conditions, the intrinsic rate of population growth (rm) is a key variable in the the population dynamics of any insect pest (Southwood and Henderson 2000). It is possible to achieve rapid population growth through a high reproduction rate, a short developmental time, or a combination of the two; however, the relative contributions of these parameters to population growth are not equal (Krips et al. 1998). In our study, female FAW larvae reared on rice had a significant lower rm value than those reared on maize. This was due to the shorter developmental time of the female fed on maize, which resulted in a higher rm value than that in rice. Previous research found that shorter development time was the most important contributor to higher rm found on maize (Lei et al. 2012). In addition, the higher rm observed in maize is likely to be attributed to a higher R0. Our results also suggested that females reared on maize have a higher net reproduction rate (R0) (Table 3). Similar variability was observed in other studies, such as that by Li-Hong et al. (2021) who reported the rm value of FAW to be similar when fed on maize and tomato but lower when fed on pepper. The rm values for rice and maize in our study were 0.121 and 0.173 respectively, falling within the range of other studies. Li-Hong et al. (2021) also reported that the rm value of FAW is 0.164 on maize, 0.147 on tomato and 0.081 on pepper. Montezano et al. (2019) found the rm of FAW fed on an artificial diet to be 0.22. He et al. (2021a) documented the rm values of FAW as 0.0844, 0.1041 and 0.1134 when reared on soybean, oilseed rape and sunflower as hosts, respectively. Wang et al. (2020) also reported the rm value of FAW to be 0.2056, 0.1827, 0.1418, 0.1296, and 0.1342 when reared on maize, wheat, soybean, tomato and cotton, respectively. Our study also indicates that the host can have a a significant impact on R0, λ, tG and tD. FAW fed on rice showed lower R0 and λ values but higher generation (tG) and doubling time (tD) than when fed maize. A similar effect was observed in other studies. For example, that maize had higher R0, and λ values but lower tG and tD values than those of all the other host plants tested (Li-Hong et al. 2020; Wang et al. 2020; Hutasoit et al. 2020).
The age at which the first egg is laid is critical for calculating rm (Hong et al. 2019). If fecundity remains constant, early reproduction will be associated with a high rm (Huang et al. 2012). Consequently, our study shows that females reared on maize started to lay eggs earlier than those fed on rice. As greater infestation levels have been reported on maize, higher rm values could be indicative of increased incidence and severity of the pest. Life table parameters can vary based on factors such as temperature, chemical pesticides, host species, and interactions with other species within the community (Zuo et al. 2015; Sousa et al. 2016; Özgökçe et al. 2018; Xie et al. 2019). The intrinsic rate of increase (rm) is a measure of an animal's physiological qualities relative to its reproductive capacity. As a metric, it iscommonly used to compare population fitness under different ecological conditions (Tibshirani and Efron 1993; Saeed et al. 2010). Therefore, higher adaptability in an insect population corresponds to a higher r value, indicating their potential ability to thrive in more diverse environments, at least in consideration of different host plants. In this study, S. frugiperda demonstrated enhanced adaptability to different host species, as evidenced by its shortest adult preoviposition period and total preoviposition period on maize, resulting in higher r values. Furthermore, variation in reproductive values were observed among individuals of the same age but different stages. We found that FAW is able to develop and reproduce on some of the common rice cultivars grown commercially by farmers in Bangladesh, although our research was conducted under controlled greenhouse conditions. Field verification under on-farm settings and in the presence of multi-species interactions is conversely needed before drawing strong conclusions on the risks posed to rice by FAW in Asia. In our study, maize was the more suitable host, based our assessment of demographic parameters. The present study demonstrates that for FAW, factors related to its fitness traits such as reproductive behavior, adult longevity, fecundity, survival, lifecycle and generation time are at least partly influenced by host plant species, and that this could potentially affect population parameters. Such information is necessary to better understand pest development and to select appropriate IPM straties. Future research should nonetheless focus on field and modeling studies of the long-term interaction and development of this pest and the rice plant, such as plant compatibility and/or resistance to the target insect pest, as well as multi-species interactions under non-controlled field conditions.
The findings of our study have practical implications for predicting population survival under specific conditions, which are crucial for more accurately forecasting the growth trend of pest populations and formulating effective IPM strategies. In pest management and decision-making, computer simulations that project pest population growth using life table data are considered essential tools (Tuan et al. 2014). Currently, the management approach for S. frugiperda in Bangladesh primarily involves monitoring male adults using pheromone traps, validating the incidence and serverity of FAW at a field level, and using this information to select from a range of IPM options. Our results suggest that management approaches applied in rice are likely to be different from maize since the increasing rate of population is significantly higher in maize than that of rice. More data from various locations aid in estimating the host range, potential economic losses, and developing effective FAW pest management measures for Bangladesh and other rice producing Asian countries.
Availability of data and materials
All data related to publication available within the manuscript are available by contacting the corresponding author.
Notes
BR3, BRRI dhan28, BRRI dhan29, BRRI dhan33, BRRI dhan48, BRRI dhan49, BRRI dhan50, BRRI dhna58, BRRI dhan67, BRRI dhan75, BRRI dhan87, BRRI dhan89, BRRI dhan92, Sadamota and Kalijira.
Abbreviations
- ANOVA:
-
Analyses of variance
- BRRI:
-
Bangladesh Rice Research Institute
- CABI:
-
Centre for Agriculture and Bioscience International
- FAO:
-
Food and Agriculture Organization
- IPM:
-
Integrated pest management
- Mx:
-
The age-specific fecundity rate (mx)
- R 0 :
-
The reproduction rate
- r m :
-
The intrinsic rate of natural increase and the
- SPSS:
-
Statistical Packages for Social Science
- t D :
-
Doubling time
- FAW:
-
Fall armyworm
- t G :
-
The mean length of generation and the
- USD:
-
United State Dollar
- λ:
-
Finite rate of increase
References
Abrahams P, Bateman M, Beale T, Clottey V, Cock M, Colemenarez Y, et al. Fall armyworm: impacts and implications for Africa. Evidence note (2). Outlooks Pest Manag. 2017;28:196–201.
Alam SN, Sarker D, Pradhan MZH, Rashid MH, Sarkar MA, Begum K, et al. First report of occurrence of fall armyworm Spodoptera frugiperda in Bangladesh. Bangladesh J Entom. 2018;28:97–101.
Ali MY, Naseem T, Arshad M, Ashraf I, Rizwan M, Tahir M, et al. Host-plant variations affect the biotic potential, survival, and population projection of Myzus persicae (Hemiptera: Aphididae). InSects. 2021;12(5):375.
Awmack CS, Leather SR. Host plant quality and fecundity in herbivorous insects. Annu Rev Entomol. 2002;47:817–44.
Barros EM, Torres JB, Ruberson JR, Oliveira MD. Development of Spodoptera frugiperda on different hosts and damage to reproductive structures in cotton. Entom Exp Appl. 2010;137(3):237–45.
Birch LC. The intrinsic rate of natural increase of an insect population. J Anim Ecol. 1948;17:15–26.
BRRI. Adhunik Dhaner Chash. Bangladesh Rice Research Institute, Gazipur, Bangladesh. Dhaka, Bangladesh; 2017.
Buntin GDA. review of plant response to fall armyworm, Spodoptera frugiperda (JE Smith), injury in selected field and forage crops. Fla Entomol. 1986;69:549–59.
CABI. Invasive species compendium datasheets—Spodoptera frugiperda (fall armyworm). http://cabidigitallibrary.org (2020). Accessed 24 May 2019.
CABI. Fall armyworm: Impacts and implications for Africa. Retrieved from (2017) https://www.cabi.org/isc/datasheet/29810
Carey JR. Applied demography for biologists: with special emphasis on insects. Oxford: Oxford University Press; 1993.
Carey JR. Insect biodemography. Annu Rev Entomol. 2001;46:79–110.
Chen X, Anjiang T, Palli SR. Identification and functional analysis of promoters of heat-shock genes from the fall armyworm, Spodoptera frugiperda. Sci Rep. 2020;10(1):1–9.
Day R, Abrahams P, Bateman M, Beale T, Clottey V, Cock M. Fall armyworm: impacts and implications for Africa. Outlooks Pest Manag. 2017;28:196–201.
De Groote H, Oloo F, Tongruksawattana S, Das B. Community-survey based assessment of the geographic distribution and impact of maize lethal necrosis (MLN) disease in Kenya. Crop Prot. 2016;82:30–5.
Devi S. Fall armyworm threatens food security in southern Africa. The Lancet. 2018;391(10122):727.
Early R, González-Moreno P, Murphy ST, Day R. Forecasting the global extent of invasion of the cereal pest Spodoptera frugiperda, the fall armyworm. NeoBiota. 2018;40:25–50.
Farias JR, Andow DA, Horikoshi RJ, Sorgatto RJ, Fresia P, dos Santos AC, Omoto C. Field-evolved resistance to Cry1F maize by Spodoptera frugiperda (Lepidoptera: Noctuidae) in Brazil Crop Protect. 2014, 64:150–158
FAO (Food and Agriculture Organization). First detection of fall army worm on the border of Thailand [EB/OL]. [2018-12-20]; 2018. https://www.ippc.int/en/countries/thailand/pestreports/2018/12/first-detection-of-fall-army-worm-on-the-border-of-thailand/.
FAO. Forecasting threats to the food chain affecting food security in countries and regions. Food Chain Crisis Early Warning Bulletin No. 34, January–March 2020. Rome (2020).
Fathipour Y, Hossain AL, Naseri B. Soybeancultivars affecting performance of Helicoverpa armigera (Lepidoptera: Noctuidae). In: Ng, T. B. (Ed.), Soybean -Biochemistry, Chemistry and Physiology.InTech, Rijeka, Croatia, 2011.599–630. http://www.intechopen.com/articles/show/title/soybean-cultivars-affecting - performance-of- helicoverpa-armigera-lepidoptera-noctuidae.
Fathipour Y, Baghery F, Bagheri A, Naseri B. Development, reproduction and life table parameters of Helicoverpa armigera (Lepidoptera: Noctuidae) on five main host plants. J Crop Prot. 2020;9:551–61.
Goergen G, Kumar PL, Sankung SB, Togola A, Tamò M, Luthe DS. First report of outbreaks of the fall armyworm Spodoptera frugiperda (J E Smith) (Lepidoptera, Noctuidae), a new alien invasive pest in West and Central Africa. PLoS ONE. 2016;11:e0165632.
Gotoh T, Gomi K. Life-history traits of the Kanzawa spider mite Tetranychus kanzawai (Acari: Tetranychidae). Appl Entomol Zool. 2003;38:7–14.
Hong F, Han HL, Pu P, Wei D, Wang J, Liu Y. Effects of five host plant species on the life history and population growth parameters of Myzus persicae (Hemiptera: Aphididae). Journal of Insect Science, 2019, 19(6):803–823
Harrison RD, Thierfelder C, Baudron F, Chinwada P, Midega C, Schaffner U, van den Burg J. Agro-ecological options for fall armyworm (Spodoptera frugiperda JE Smith) management: providing low-cost, smallholder friendly solutions to an invasive pest. J Environ Manag. 2019;243(318):330.
He LM, Wang TL, Chen YC, Ge SS, Wyckhuys KAG, Wu KM. Larval diet affects development and reproduction of East Asian strain of the fall armyworm, Spodoptera frugiperda. J Integr Agric. 2021a;20:736–44.
He LM, Wu QL, Gao XW, Wu KM. Population life tables for the invasive fall armyworm, Spodoptera frugiperda fed on major oil crops planted in China. J Integr Agric. 2021b;20:745–75.
Heinrichs EA, Nwilene FE, Stout MJ, Hadi BAR, Freitas T. Rice insect pests and their management. USA: Burleigh Dodds Science Publishing; 2017. p. 360p.
Huang YB, Chi H. Age-stage, two-sex life tables of Bactrocera cucurbitae (Coquillett) (Diptera: Tephritidae) with a discussion on the problem of applying female age-specific life tables to insect populations. Insect Sci. 2012;19(2):263–73.
Hutasoit R, Kalqutny SH, Widiarta D. Spatial distribution pattern, bionomic, and demographic parameters of a new invasive species of armyworm Spodoptera frugiperda (Lepidoptera; Noctuidae) in maize of South Sumatra, Indonesia. Biodiversitas J Biol Divers. 2020;21:3576–82. https://doi.org/10.13057/biodiv/d210821.
Kavallieratos NG, Andri G, Golić MP, Nika EP, Skourti A, Kljajić P, Papanikolaou NE. Biological features and population growth of two Southeastern European Tribolium confusum Jacquelin du Val (Coleoptera: Tenebrionidae) strains. InSects. 2020;11(4):218.
Koussoroplis AM, Wacker A. Covariance modulates the effect of joint temperature and food variance on ectotherm life-history traits. Ecol Lett. 2016;19(2):143–52.
Krips OE, Witul A, Willems PEL, Dicke M. Intrinsic rate of population increase of the spider mite Tetranychus urticae on the ornamental crop gerbera: intraspecific variation in host plant and herbivore. Entomol Exp Appl. 1998;89(2):159–68.
Kumela J, Simiyu B, Sisay P, Likhayo E, Mendesil L, Gohole T. Farmers' knowledge, perceptions, and management practices of the new invasive pest, fall armyworm (Spodoptera frugiperda) in Ethiopia and Kenya Int. J. Pest Manag. 2018, 65(1):1–9.
Li-Hong Wu, Zhou C, Long GY, Yang XB, Wei ZY, Liao YJ, Yang H, Hu CX. Population life tables for the invasive fall armyworm, Spodoptera frugiperda fed on major oil crops planted in China. J Integr Agric. 2020;18:755–63.
Lei X, Li D, Li Z, Zalom FG, Gao L, Shen Z. Effect of host plants on developmental time and life table parameters of Carposina sasakii (Lepidoptera: Carposinidae) under laboratory conditions. Environ Entomol. 2012;41(2):349–54.
Li-Hong Wu, Zhou C, Long GY, Yang XB, Wei ZY, Liao YJ, Yang H, Hu CX. Fitness of fall armyworm, Spodoptera frugiperda to three solanaceous vegetables. J Integr Agric. 2021;20:755–63.
Luginbill P. The fall armyworm. US Dept Agric Tech Bull. 1928;34:1–91.
Maia AHN, Luiz AJB, Campanhola C. Statistical inference on associated fertility life table parameters using jackknife technique: computational aspects. J Econ Entomol. 2000;93:511–8.
McCormick AC, Arrigo L, Eggenberger H, Mescher MC, De Moraes CM. Divergent behavioural responses of gypsy moth (Lymantria dispar) caterpillars from three different subspecies to potential host trees. Sci Rep. 2019;9(1):1–12.
Mehrkhou F, Talebi AA, Moharramipour S, Naveh VH. Demographic parameters of Spodoptera exigua (Lepidoptera: Noctuidae) on different soybean cultivars. Environ Entomol. 2012;41(2):326–32.
Meyer JS, Ingersoll CC, McDonald LL, Boyce MS. Estimating uncertainty in population growth rates: jackknife vs bootstrap techniques. Ecology. 1986;67:1156–66.
Montezano DG, Specht A, Sosa-Gómez DR, Roque-Specht VF, Sousa-Silva JC, Paula-Moraes SV, Peterson JA, Hunt T. Host plants of Spodoptera frugiperda (Lepidoptera: Noctuidae) in the Americas. Afr Entomol. 2018;26:286–300.
Montezano DG, Specht A, Sosa-Gómez DR, Roque-Specht VF, Malaquias JV, Paula-Moraes SV, Peterson JA, Hunt TE. Biotic potential and reproductive parameters of Spodoptera frugiperda (J. E. Smith, 1797) (Lepidoptera: Noctuidae). J Agric Sci. 2019;11:240. https://doi.org/10.5539/jas.v11n13p240.
Murúa G, Virla E. Population parameters of Spodoptera frugiperda (Smith) (Lep.: Noctuidae) fed on maize and two predominant grasses in Tucuman (Argentina). Acta Zool Mex. 2004;20:199–210.
Nagoshi RN, Pierre S, Robert LM. Comparison of haplotype frequencies differentiate fall armyworm (Lepidoptera: Noctuidae) corn-strain populations from Florida and Brazil. J Econ Entomol. 2007;100(3):954–61.
Nakweta G. Global actions needed to combat fall armyworm. [2018-10-05]; 2018. https://www.scidev.net/subsaharan-africa/farming/news/global-actions-combat-fall-armyworm.html.
Naseri B, Fathipour Y, Moharramipou S, Hosseininaveh V. Comparativereproductive performance of Helicoverpaarmigera (Hübner) (Lepidoptera: Noctuidae)reared on thirteen soybean varieties. Journalof Agricultural Science and Technology, 2011. 13:17–26
Overton K, Maino JL, Day R, Umina PA, Bett B, Carnovale D, Ekesi S, Meagher R, Reynolds OL. Global crop impacts, yield losses and action thresholds for fall armyworm (Spodoptera frugiperda): a review. Crop Prot. 2021;145:105641.
Özgökçe MS, Chi H, Atlıhan R, Kara H. Demography and population projection of Myzus persicae (Sulz.) (Hemiptera: Aphididae) on five pepper (Capsicum annuum L.) cultivars. Phytoparasitica. 2018;46:153–67. https://doi.org/10.1007/s12600-018-0651-0.
Pantoja A, Smith CM, Robinson JF. Effects of fall armyworm (Lepidoptera: Noctuidae) on rice yields. J Econ Entomol. 1986;79:1324–9.
Rwomushana I, Bateman M, Beale T, Beseh P, Cameron K, Chiluba M, Clottey V, Davis T, Day R, Early R, Godwin J, Gonzalez-Moreno P, Kansiime M, Kenis M, Makale F, Mugambi I, Murphy S, Nunda W, Phiri N, Tambo J. Fall Armyworm: impacts and implications for Africa Evidence Note Update, October 2018. Report to DFID. CABI; 2018.
Sabelis MW. Reproductive strategies. In: Helle W, Sabelis MW, editors. Spider mites: their biology natural enemies and control. Amsterdam: Elsevier; 1985. p. 265–78.
Saeed R, Sayyed AH, Shad SA, Zaka SM. Effect of different host plants on the fitness of diamond-back moth, Plutella xylostella (Lepidoptera: Plutellidae) Crop. Crop Protect. 2010;29:178–82. https://doi.org/10.1016/j.cropro.2009.09.012.
Sartiami D, Dadang IS, Harahap YM, Kusumah R. Anwar. First record of fall armyworm (Spodoptera frugiperda) in Indonesia and its occurrence in three provinces. In IOP Conference Series: Earth Environmental Science, 2020. vol. 468, p. 012021.
Sharanabasappa K, Maruthi MS, Pavithra HB. Biology of invasive fall army worm Spodoptera frugiperda (J.E. Smith) (Lepidoptera: Noctuidae) on maize. Indian J Entomol. 2018;80:540–3.
Silva CLT, Correa F, Almeida ACS, Araújo MS, Barrigossi JA, Jesus FG. Resistance of rice genotypes to fall armyworm Spodoptera frugiperda (Lepidoptera: Noctuidae). Rev Bras Entomol. 2021;65(3):e20210020. https://doi.org/10.1590/1806-9665-RBENT-2021-0020.
Silva D, Bueno A, Andrade K, Stecca C, Neves P, Oliveira M. Biology and nutrition of Spodoptera frugiperda (Lepidoptera: Noctuidae) fed on different food sources. Sci Agric. 2017;74:18–31.
Soto EM, Padró J, Milla Carmona P, Tuero DT, Carreira VP, Soto IM. Pupal emergence pattern in cactophilic Drosophila and the effect of host plants. Insect Sci. 2018;25(6):1108–18.
Sousa F, Mendes S, Santos Amaya O, Araujo O, Oliveira E, Pereira E. Life-history traits of Spodoptera frugiperdapopulations exposed to low-dose Bt maize. PLoS ONE. 2016;11:e0156608. https://doi.org/10.1371/journal.pone.0156608.
Southwood TRE, Henderson PA. Ecological methods. 3rd ed. New York: Wiley-Blackwell; 2000.
Sun XX, Hu CX, Jia HR, Wu QL, Shen XJ, Zhao SY, Jiang YY, Wu KM. Case study on the first immigration of fall armyworm, Spodoptera frugiperda invading into China. J Integr Agric. 2021;20:664–72.
Tibshirani RJ, Efron B. An introduction to the bootstrap, vol. 57. London: Chapman & Hall; 1993. p. 45–82.
Tuan SJ, Lee CC, Chi H. Population and damage projection of Spodoptera litura (F.) on peanuts (Arachis hypogaea L.) under different conditions using the age-stage, two-sex life table. Pest Manag Sci. 2014;70:805–13. https://doi.org/10.1002/ps.3618.
Wang W, Zhang P, Liu Y, Jing TX, Zhang X. The population growth of Spodoptera frugiperda on six cash crop species and implications for its occurrence and damage potential in China. InSects. 2020. https://doi.org/10.3390/insects11090639.
Xie DJ, Zhang L, Cheng YX, Jiang XF. Age-stage two-sex life table for laboratory populations of fall armyworm, Spodoptera frugiperda at different temperatures. Plant Prot. 2019;45:20–7.
Yang XL, Liu YC, Luo MZ, Li Y, Wang WH, Wan F, Jiang H. Spodoptera frugiperda (J E Smith) was first discovered in Jiangcheng County of Yunnan Province in southwestern China. Yunnan Agric. 2019;01:72 (in Chinese).
Zuo TQ, Zhang B, Zhang ST, Zheng CY, Wan FH. Combined effects of high temperature and acetamiprid on life table parameters of the F1 offspring of the treated Frankliniella occidentalis (Thysanoptera: Thripidae). Acta Entomol Sin. 2015;58:767–75.
Acknowledgements
The authors thank the staff and scientists of the Entomology Division for their assistance in rearing of Fall Armyworm in BRRI, Gazipur, in the greenhouse work and the Bangladesh Rice Research Institute (BRRI) for providing other necessary support.
Funding
This research was funded as part of the USAID/Bangladesh and Michigan State University (MSU) supported Fighting Back against Fall Armyworm (FAW) in Bangladesh project and by the USAID/Bangladesh and Virginia Tech (VT) Unviersity supported Integrated Pest Management Activity (IPMA). Further support was provided under Work Package 2 'Transforming agroecosystems and rural economies to boost income, and support diversified food production within environmental boundaries' of the CGIAR Regional Integrated Initiative Transforming Agrifood Systems in South Asia, or TAFSSA (https://www.cgiar.org/initiative/20-transforming-agrifood-systems-in-south-asia-tafssa/). The views and opinions expressed in this publication are those of the author(s) and are not necessarily representative of or endorsed by USAID, MSU, VT or CGIAR and shall not be used for advertising purposes.
Author information
Authors and Affiliations
Contributions
Conceptualization—MPA, SSH, TJK, SNA and MTHH; experiments and statistical analysis, MPA, SSH, MMH, MNB, MMMK, TR, MMH, JD; writing—original draft preparation, MPA, and MTHH; writing—review and editing, MPA, SNA, TJK, and MTHH; materials and equipment, MPA, SSH, MN, MMH, TR and JD; supervision, MPA, SSH, MMH; MNB, and MMMK funding acquisition TJK. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Ali, M.P., Haque, S.S., Hossain, M.M. et al. Development and demographic parameters of Fall Armyworm (Spodoptera frugiperda J.E. Smith) when feeding on rice (Oryza sativa). CABI Agric Biosci 4, 29 (2023). https://doi.org/10.1186/s43170-023-00162-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s43170-023-00162-6