
Abstract
Background
The Capparaceae family is commonly recognized as a caper, while Cleomaceae represents one of small flowering family within the order Brassicales. Earlier, Cleomaceae was included in the family Capparaceae; then, it was moved to a distinct family after DNA evidence. Variation in habits and a bewildering array of floral and fruit forms contributed to making Capparaceae a “trash-basket” family in which many unrelated plants were placed. Indeed, family Capparaceae and Cleomaceae are in clear need of more detailed systematic revision.
Results
Here, in the present study, the morphological characteristics and the ecological distribution as well as the genetic diversity analysis among the twelve species of both Capparaceae and Cleomaceae have been determined. The genetic analysis has been checked using 15 ISSR, 30 SRAP, and 18 ISTR to assess the systematic knots between the two families. In order to detect the molecular phylogeny, a comparative analysis of the three markers was performed based on the exposure of discriminating capacity, efficiency, and phylogenetic heatmap. Our results indicated that there is a morphological and ecological variation between the two families. Moreover, the molecular analysis confirmed that ISTR followed by SRAP markers has superior discriminating capacity for describing the genetic diversity and is able to simultaneously distinguish many polymorphic markers per reaction. Indeed, both the PCA and HCA data have drawn a successful annotation relationship in Capparaceae and Cleome species to evaluate whether the specific group sort individual or overlap groups.
Conclusion
The outcomes of the morphological and ecological characterization along with the genetic diversity indicated an insight solution thorny interspecies in Cleome and Gynandropsis genera as a distinct family (Cleomaceae) and the other genera (Capparis, Cadaba, Boscia, and Maerua) as Capparaceae. Finally, we recommended further studies to elucidate the systematic position of Dipterygium glaucum.
Similar content being viewed by others

Background
The flora of Egypt is among the richest within the Arab countries and comprises very vital genetic resources of medicinal, fodder, and fiber plants. Currently, plant diversity is under threat as never before. In agriculture, the broad selection of a few developed varieties has reduced the genetic base of the most essential food crops, and it has added to the withdrawal of hundreds of landraces [1]. The complicated relationship between the three closely related families, Capparaceae, Cleomaceae, and Brassicaceae, has been extensively studied since the appearance of the theory of alternation of generation by Hofmeister [2]. Hutchinson [3] in his phylogenetic work differentiated between Capparaceae and Brassicaceae according to their morphological features to two separate families. Shifting of some species between the two families is common, like Dipterygium glaucum has been treated as genera of Brassicaceae [4], then moved to Capparaceae as a subfamily: Dipterigpideae [5]. Tackholm [6] classifies Dipterygium as a species of Cruciferae, while Hedge et al. [7] classify it as a member of Capparaceae. This assumption has been reinforced by Boulos [8].
In the Egyptian flora, the set of Capparaceae plus Cleomaceae includes seven genera, twenty-two species, and four varieties with a wide range of ecological and geographical distribution [8]. They differ considerably in their life forms from trees (e.g., Boscia angustifolia) or shrubs (e.g., Capparis cartilaginea) to annual (e.g., Gynandropsis gynandra) or perennial herbs (e.g., Cleome amblyocarpa). The Egyptian taxa of Capparaceae refer to the xerophytic communities [9, 10], without Gynandropsis gynandra that is a common weed of the arable fields [11]. The taxonomic approach of the family in Egypt is interested only on seed morphology [12], leaf anatomy [13], and pollen morphology [14]. The systematic review of the natural species of Capparaceae (except Cleome) announced the unclear occurrence of Boscia angustifolia, while Capparis spinosa is described by three varieties, viz., deserti, spinosa, and inermis [15]. Spilt from Cleomaceae may be unreasonable since complications are met in selecting the genera. Accurate qualified information on fruit construction and gynoecium is obscure or non-present. Therefore, the taxonomic relationships between Capparaceae and Cleomaceae are still at discussion. Tackholm [6] distinguished among the two families concurring to gland formation, fruit type, and development of gynophores, whereas Zohary [16] included the exciting genus Cleome in the subfamily Cleomoideae of Capparaceae. The Capparaceae in Boulos [8], however, involved together Cleomaceae and Capparaceae. On the species level, Tackholm [6] identified eight species of Capparis, while Boulos [8] divided the genus as three species and four varieties. Though flower and fruit types have been shown very beneficial in the description and definition of the genera and species, there are situations in which these tools are not potential for the study as in Capparaceae. The subject of the reproductive characteristics of this group is uncertain for various purposes, among others; the challenge of maintaining the flowers in some genera as in Capparis [17] the unusual variability in their dimension and form at the individual level species [18] and several long-lived tropical plant flowers are uncommon and different [19]. Therefore, there is a great necessity to distinguish and order the wild plants utilizing morphological and molecular tools. Hence, the investigation of molecular information is the main outcome in our opinion of plant phylogeny and systematic, from the low joints dividing of the main plant groups to varieties and populations [20]. Molecular identification represents an effective instrument for genome analysis and allows the linkage of heritage traits attached to genomic divergence. Presently, these genomic tools are valuable basics for knowledge and improving resource for understanding and developing various frequent sequence approaches such as microsatellites and retrotransposon loci. Within PCR-based approaches, inter-simple sequence repeats (ISSRs) became to be addressed development of minisatellite DNA for classification of varieties or species and population genetic structure, with high efficiency, stability, low cost, and simple operation [21, 22]. Previous research of ISSR studies on several wild plant species, involving endangered and rare genus, have elucidate the hypervariable type of microsatellite loci and their inherent utility in species identification and inhabitant’s diversity [23,24,25], while sequence-related amplified polymorphism (SRAP) technique, which favorably aims regions including promoters and exonic sequences [26], have several performances, containing the use for a rather minor quantity of DNA, and high levels of polymorphism amplification targeting an open reading frames (ORFs) in many plant species [27].
In regard to inverse sequence-tagged repeat (ISTR), it is a retrotransposon-based marker [28] which has been viewed in most form of the organism, is widespread in distribution, effective, and plentiful in eukaryotic genomes [29, 30]. Therefore, ISTR markers are co-dominant markers which can define the wild flora and phylogeny at a subgenus level [31]. The three molecular markers recorded beyond are co-dominant or dominant inheritance, and the collectively utilized could be farther beneficial to identify diverse portions of the genome [32]. Consequently, comparison is necessary to determine the marker sensitivity and appropriateness for the topic being studied [33].
The current investigation was aimed to provide a broad description of the morphological, ecological distribution attributes, and connected species along with three selected molecular markers of 12 representing species of Capparaceae and Cleomaceae. In detail, first, family Capparaceae is in clear necessity of more detailed morphological, ecological, and molecular study. Modulation inhabits and a confusing display of floral and fruit profiles provided to forming Capparaceae a “trash-basket” family in which several discrete plants were ordered. Second is to evaluate the discrimination capacity and the performance of the three marker system involving ISSR, SRAP, and ISTR and, finally, to infer the taxonomic questions and species borders treating the genetic diversity of Egyptian Capparaceae and Cleomaceae germplasm. To date, there has been no announcement concerning the performance and effectiveness of ISSR, SRAP, and ISTR markers in Egyptian Capparaceae and Cleomaceae species.
Methods
Plant materials
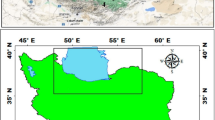
A total of twelve plant samples of Capparaceae and Cleomaceae were collected from Sinai Peninsula (three plants), Western desert (two plants), and Eastern desert (seven plants) (Fig. 1) (Table 1). Associate species were recorded, and identification was performed according to Boulos [8, 11]. Plant density and cover were assessed by the random plot and line intercept methods. A hierarchical dichotomous analysis (TWINSPAN) was used to classify plant species due to their density by PAST 3.14 [34].

Map showing the distributions and locations of 12 Capparaceae species within Western, Eastern desert, and Sinai Peninsula in Egypt
DNA extraction
A fresh leaf of Capparaceae and Cleomaceae samples was used to extract the total genomic DNA according to the manufacturer’s protocol using plant DNA purification mini kit (Thermo Scientific GeneJET kit, K0791, USA). In each species, three to five replicate DNAs were utilized. However, the concentration and quality of the DNA samples were verified in a Quawell Q5000 UV-Vis spectrophotometer (V2.1.4, USA); then, the DNA was diluted to 50 ng/μl for use in ISSR, SRAP, and ISTR assay. Both the stock and diluted portions were kept at − 20 °C.
ISSR analysis
The ISSR-PCR amplification was prepared according to the earlier method demonstrated by Sankar and Moore [35]. The amplification reaction of ISSR analysis was done in a total volume of 25 μl on a Sure Cycler 8800 Thermal Cycler from Agilent Technologies. The reaction combination of 25 μl involved 3.5 μl of Green PCR buffer, 1 μM of each primer, 0.5 μM of dNTPs (10 mM each) (Thermo Fisher Scientific), 1 unit of taq DNA polymerase (5 U/μL) (Thermo Fisher Scientific), and 40 ng DNA template. The PCR program was as follows, denaturation (one cycle) in 94 °C for 2 min, followed by 40 cycles as follows: 94 °C for 30 s, 44 °C for 45 s, 72 °C for 1 min and 30 s, and finally one cycle extension at 72 °C for 10 min, and 4 °C (infinitive). The amplified products were separated on 1.2% agarose gel by electrophoresis. A 100 bp DNA ladder (GeneRuler plus, Thermo Scientific, SM0321) was utilized as the molecular guideline to verify the competent ISSR markers. The gels were stained in ethidium bromide (0.5 μg/ml), and the amplicons were pictured below UV light using the Gel Doc XR system (Bio-rad, America).
SRAP analysis
The SRAP analysis was presented as illustrated by Li and Quiros [26]. SRAP primer combinations were tested using 30 various combinations which employed utilizing seven reverse and nine forward primers applied (Table 2). Every PCR reaction mix of 25 μl included 3.5 μl of green PCR buffer, 0.3 μM of each primer, 200 μM of dNTPs, 1 unit of taq DNA polymerase, 30 ng of genomic DNA, and deionized water up to 25 μl. PCR cycling program comprised 4 min of denaturing at 94 °C, five cycles of three steps: 1 min of denaturing at 94 °C, 1 min of annealing at 35 °C, and 1 min of elongation at 72 °C. In the next 35 cycles, the annealing temperature was increased to 50 °C, and for an extension, one cycle of 7 min at 72 °C. GeneRuler 50 bp Plus DNA ladder (Thermo Scientific, SM0371) was utilized as a molecular guideline to verify the accurate SRAP markers.
ISTR analysis
ISTR assessment was conducted following Aga and Bryngelsson [36]. ISTR primer combinations were primarily examined using a total of 70 primer combinations from seven reverse and ten forward primers. Within all primers screened, only 24 ISTR combinations were picked for advanced analysis (Table 2). Each PCR included a reaction mix of 3.5 μl of green PCR buffer, 200 μM of dNTPs, 0.3 μM of each primer, 50 ng of genomic DNA, 1 unit of taq DNA polymerase, and finally deionized water up to 25 μl. PCR amplification performed involved of 1 cycle at 95 °C, 3 min; 40 cycles of 94 °C, 30 s; 45 °C, 30 s; 72 °C, 2 min; 1 cycle at 72 °C, 10 min; and 4 °C for infinitive. However, amplification products were separated and visualized subsequent the same procedure described for ISSR.
Data analysis
All clearly detectable ISSR, SRAP, and ISTR products were counted as band absence (0) and presence (1) using the Bio-Rad Gel Doc™ XR+ imaging analysis system with Image Lab™ (USA), and adjusted manually as necessary and collected onto a data matrix. However, the evaluations of the marker efficiency, level of polymorphism, discriminating capacity, and informativeness of the three marker profiles were calculated according to the indices of Powell et al. [37].
To measure the effectiveness of the three marker systems, polymorphic information content (PIC) was analyzed using the following formula of Roldán-Ruiz et al. [38] PIC = 2fi(1 − fi), where fi is the frequency of the amplified allele and 1 – fi is the frequency of the null allele. While heterozygosity per locus was determined according to the formula: He = 1 − p2 − q2, where p2 = fi. Meanwhile, the average heterozygosity per marker was evaluated based on Hav = ∑(He/L), where L = total of detected bands. The multiplex ratio was calculated as MR = L/T, where T = the total number of primer combinations. The marker index (MI) was achieved by developing the average heterozygosity by the multiplex ratio: MI = Hav X MR.
To gain accurate perspectives on genetic diversity analyses among the Capparaceae and Cleomaceae germplasm, a graphic demonstration of principal coordinates analysis (PCA) and the heatmap cluster analysis (HCA) was provided an explanation to demonstrate the multidimensional genetic relationship and its split among species using ClustVis web tool for visualizing clustering of multivariate data [39].
Results
Morphological basis of Capparaceae taxonomy
In the present scenario, the main differences between Capparis and Cleome are growth habit and life span, as presented in Fig. 2. All individuals of Capparaceae are trees or shrubs, whereas species of Cleome are annual or perennial herbs. According to the type of fruits of the family, Cleomaceae were divided into Dipterygium (fruits one-seeded), Gynandropsis, and Cleome (fruits contains much seed), and then separated according to the number of stamens and presence or absence of androphore. On the other hand, family Capparaceae is classified into four species due to the presence of a stipule spine (Capparis) and the absence of a stipule (Maeura, Cadaba, and Boscia). Boscia species leave their group due to its fruit type (hard indehiscent). Recently, both Maeura and Cadaba are isolated according to the number of stamens and their contact with the androphore.

Morphological basis of Capparaceae taxonomy
Distribution and habitat classification of Capparaceae and Cleomaceae species
According to vegetation density and cover, classification of the twelve targeted species and 29 associates are mainly divided according to species distribution, habitat, and locations to six vegetation groups. The first group comprises of Maerua crassifolia and Capparis deserti inhabiting the sandy formation of the southern wadis of the Eastern desert and Siwa oasis, respectively. Moreover, they are tending to form pure communities with high density and cover. The second group is formed of Cleome and Dipterygium, and they are collected from the main channel of Wadi Abrq, Shalatein area. The third group involves of Capparis spinosa var. canescens and C. aegyptia, and they are similar in habitat; both of them are hanging between the rock fissures at south Sinai. While the fourth group (Cadaba farinosa and Capparis decidua) is distributed on the sandy formation of red sea wadis, the fifth group is represented with Capparis spinosa var. inermis; this species is restricted to the maritime cliffs and rocky ridges at the western Mediterranean section. However, the last group is occupied with the biggest Capparis species (Capparis cartilaginea) (Fig. 3).

The tree represents the classification of the twelve selected Capparaceae and Cleomaceae species of the study, in blue, together with the 29 accompanying species in red
Comparison of polymorphic levels and informativeness obtained with ISSR, SRAP, and ISTR markers
In the present investigation, the levels of polymorphism of ISSR, SRAP, and ISTR markers and the index associating their informativeness are described in Table 3 and Fig. 4. All markers used pointed out to be helpful implements for the discovery of polymorphism and evaluating genetic diversity in Capparaceae and Cleome genotypes, but the level of sensitivity varied on the method applied. We primarily tested 20 of ISSR primers and 49 and 70 combinations of SRAP and ISTR primers among the twelve Capparaceae and Cleome species, respectively. Among all, only 15 ISSR, 30 SRAP, and 18 ISTR primers exhibited significant levels of polymorphism as shown in Table 3 and Fig. 4a–c. The total number of bands recorded for SRAP was almost high with 503 bands, followed by 337 and 252 bands for ISTR and ISSR markers, respectively. However, the total numbers of polymorphic bands (p) were ranged from 479 for SRAP, 333 for ISTR, to 239 for ISSR markers. On behalf of the total number of effective alleles (Ne), it was correlated significantly with the total number of bands (L) and the total numbers of polymorphic bands (p). Additionally, the average number of polymorphic bands/assay unit (np/U) was relatively high for ISTR being 18.5 with an intermediate value of 15.96 and 15.93 for SRAP and ISSR, respectively. Meanwhile, the PIC value for ISSR, SRAP, and ISTR marker system was almost parallel and relatively high being, 0.97, 0.98, and 0.99, respectively. Here, the present result showed that the ISTR marker was the most powerful marker in several detected parameters and PIC values. A comparative summary of the discriminating capacity of ISSR, SRAP, and ISTR markers are summarized in Table 3 and Fig. 5. On average, the three factors, assay efficiency index (Ai), effective multiples ratio (E), and marker index (MI), presented higher in ISTR marker, highlighting the notable characteristics of this marker compared to SRAP and ISSR (2.4×, 1.2× and 1.2× respectively).This certainly is due to the highest value of the assay efficiency index for the ISTR marker, inferring that ISTR has a higher discriminating capacity for counting the genetic diversity and can concurrently discover many polymorphic markers per reaction. Although the variances in some of the diversity statistics, these outcomes reveal that ISTR following by SRAP markers can be applied to assess the level of polymorphism in Capparaceae and Cleome species.

Observed the ISSR (a), SRAP (b), and ISTR (c) profiles of 12 Capparaceae species

Observed the comparison information obtained and the discriminating capacity of a ISSR, b SRAP, and c ISTR profile among 12 Capparaceae species
Diversity analyses and phylogenetic heatmap
Here, we present a graphic demonstration of both PCA and HCA, utilizing variable information matrix as input, wherever numerous features of ISSR, SRAP, and ISTR marker data are assessed in several observations. The overall PCA plot data for the three marker profile as shown in Fig. 6 created four relatively clustered groups, with a total of 19.73% of the molecular variance (PC1—10.32%, PC2—9.41%). Cluster I compressed Capparis spinosa inermis, Capparis spinosa var. canescens, Capparis spinosa deserti, Capparis cartilaginea, and Capparis aegyptia with a closer relationship than other groups. Moreover, cluster II assembled Cleome droserifolia, Cleome chrysantha, and Cleome amblyocarpa in a particular group, while Maerua crassifolia, Capparis decidua, and Dipterygium glaucum places jointly as cluster III. However, Cadaba farinosa was separated individually as out-group species.

Schematic representation the principal coordinates analysis of 12 Capparaceae species based on ISSR, SRAP and ISTR markers. PC1and PC2 refer to the first and second principal component, respectively
To further determine the genetic diversity, HCA exhibits the abundance of the relationships between the twelve species of Capparaceae and Cleome. The distribution of hot points indicates significant variations between the major groups of the Capparaceae and Cleome species and able to cluster in a sub-clade. As a result, the HCA was constructed based on the three sets of ISSR, SRAP, and ISTR markers (Fig. 7). The results were similar to each other with a tiny difference in the placement of some species, where the ISTR-HCA tree was the most consistent with the morphological taxonomy data of Capparaceae. Overall, three confirmed clades were identified, which have the ability to clearly distinguish among the twelve species. In detail, the first clade assembled together Capparis decidua, Cadaba farinosa, and Maerua crassifolia in a particular monophyletic clade. However, the three Cleome species and Dipterygium glaucum were placed jointly in the second clade with a high proportion of close relationships. In the framework, the third clade formed two monophyletic sub-clades, not based on their type but on their sampling origin, where the three species of Capparis spinosa are place jointly with high portions within the first sub-clade, whereas the second sub-clade occupies Capparis aegyptia and Capparis cartilaginea with a close genetic relationship. Collectively, we found both PCA and HCA data have drawn a successful annotation relationship in Capparaceae and Cleome species to evaluate whether the specific group sort individual or overlap groups.

Heatmap cluster analysis (HCA) signatures among 12 Capparaceae species using ISTR profiles. Subclades are highlighted by a colorful background; the scale bar showed on the top illustrates the relative genetic variability from 0.5 to − 0.5
Discussion
The systematic approach and phylogenetic relationships of the Egyptian Capparis and Cleome species remain obscure, and unsolved dilemmas regarding their taxonomy and biology require further verifying and review [40]. Species identification in Capparis and Cleome is problematic because of the challenge of preserving the flowers [17] or maybe impossible when only vegetative parts are present, which is commonly the case during collection. Ettingshausen [41] made the primary comprehensive attempt to systematize the description of the vegetative leaf architecture together with his classification of venation patterns. Leaf architectural characters have demonstrated valuable taxonomic and systematic data both in fossil and living plants [42,43,44]. Leaf architecture and venation pattern were examined in numerous families of dicotyledons, among others, Composite [45], Solanaceae [46], Bignoniaceae [47], Hamamelidaceae sensu lato [48], Leguminosae [49], Amaranthaceae [50], Ulmaceae [51], Fagaceae [52], and in some monocots [53].
Various traditional taxonomic positions of Capparis and Cleome were derived from the study of both quantitative and qualitative macro-morphological characters which might become a broad margin of mistake. Therefore, additional knowledge about the genotype of plants is much required to resolve taxonomic problems in these genera [54]. Hence, evaluations of molecular statistics has had a serious effect on our perception of plant evolution and relations on all taxonomic levels, from the deep nodes dividing the key plant groups to species and populations [20]. Consequently, studying the levels of genetic diversity in natural populations is an essential precursor for the survey of plant species and will offer perceptions about the evolution of the species [55]. During the last two decades, there are little individual molecular studies involving Capparis and Cleome like the RAPD marker [54, 56] and therefore, the AFLP marker [57].
In the same context, the selection of the foremost appropriate marker system for a certain survey is not evident and principally depends on the aim of the research because the genetic structure of the species was varied [33]. The use of ISSR, SRAP, and ISTR marker is strongly recommended by several studies addressing the effectiveness of such markers for investigating Capparaceae diversity. One of the challenges of the current research is the use of ecological and molecular markers to explore the genetic relationships among the Egyptian Capparis and Cleome species grown within the Egyptian desert. In the present investigation, the relatively high values of the effective number of alleles for ISSR, SRAP, and ISTR markers were used to provide indication of their discrimination capacity when study a huge number of plants. This trend is required for the germplasm bank’s certification when multiple species require to be correctly distinguished and classified [58]. In this revised, the effective number of alleles tracking the method: SRAP > ISTR > ISSR. This result suggests that the ISTR and SRAP is more useful evidence for Capparis and Cleome species classification and certification. It is well-known that the marker index (MI) may probably be a suitable value for marker effectiveness [58]. By this criterion, arithmetically 1.18 fold greater MI was estimated for ISTR against SRAP and ISSR, highlighting the unique character of the ISTR assay. This is definitely owing to the superior value of effective multiples ratio (EMR) and assay efficiency index (Ai) [59]. Many studies confirmed that the retrotransposons marker, e.g., ISTR had a superior discrimination capacity and have the flexibility to detect several polymorphic loci per individual reaction [60]. Recently, Du et al. [61] suggested that retrotransposons (RT) occupied 28.1 Mb of the genome sequence, accounting for 9.74% of the entire genome. These results indicated that ISTR had an abundant presence of Ty-1 Copia retrotransposons, which permit obtaining useful polymorphism among the tested genotypes of Capparaceae and Cleome germplasm. Indeed, our finding showed that RT-ISTR markers had numerous unique private loci that would allow diversity within the sub-species of Capparaceae and Cleome germplasm, which is in concurrence with earlier reports of this marker [31, 62].
In the Mediterranean area, five species are confirmed to be growing, e.g., Capparis aegyptia Lam. Boiss., Capparis spinosa L., Capparis orientalis Veil., Capparis sicula Veil., and Capparis ovata Desf [63, 64]. Mainly, in various floristic works, Capparis aegyptia is recognized from Northern Africa and also the Mideast [65]. In Flora Hellenica [66], it has been raised, Capparis aegyptia Lam., to a subspecies of Capparis spinosa, while Inocencio et al. [63] returned its status of individual species, while Özbek and Kara, [67] fully proposed that the two subspecies Capparis spinosa L. and Capparis ovata Desf. might be distinguished roughly. However, Inocencio et al. [63] confirm that Capparis aegyptia Lam. and Capparis ovata are very distinct from the opposite taxa but extremely tight to each other.
Recently, Al-Safadi et al. [64] recognized three Capparis species growing in Syria, C. sicula Duh, C. aegyptia Lam, and C. spinosa L., and the results support this theory that C. aegyptia Lam. might be an independent species and not a varietal level of C. spinosa. Initially, Rivera et al. [68] discussed the origin of Capparis cartilaginea and indicated that this is often a widely accepted name. Following the phylogenetic analysis of Saifi et al. [69], the dendrogram was assembled together with Capparis cartilaginea, Capparis aegyptia Lam, and Capparis spinosa in a sub-group. In view of the previous revisions, our phylogenetic analysis clear evidence supporting the undisputed viewpoint that Capparis aegyptia and Capparis cartilaginea are very closely associated with Capparis spinosa in a sister clade and seem to be distinguished. These results might be applied in systematics and evolutionary biology studies within the Egyptian species of Capparis and Cleome to clarify the complex interactions among species, as demonstrated in previous studies [64, 70]. Indeed, we are able to tentatively imply forward this theory as Capparis decidua, Cadaba farinosa, Maerua crassifolia, and Dipterygium glaucum formed a particular monophyletic clade with Cleome species. Based on the above considerations, our results verified the conclusions of systematic and taxonomic analyses that were performed on the collected samples, which is in consensus with previous articles on Capparis species [54, 57]. Interestingly, we observed a thorny interspecies in Capparis decidua, Cadaba farinosa, Maerua crassifolia, and Dipterygium glaucum specimens and also the concerned during this study. Currently, the advent of molecular markers overcame the majority of the challenges related to utilizing morphological markers during which main phenotype-varying genes were applied as genetic markers [71].
Conclusion
In the present investigation, we emphasize that the outcomes of the morphological and ecological characterization as well as the genetic analysis based ISSR, SRAP, and SRAP can capture the taxonomy and systematics of the various subgroups recovered with a good performance in clarifying genetic diversity within and among populations in the Egyptian Capparis and Cleome species. Our findings show Cleome and Gynandropsis genera as an explicit family; therefore, an in-depth study like next-generation sequencing (NGS) technologies is now emerging as precision tools to assess the molecular systematics and evolution in Dipterygium glaucum (if belongs to Capparaceae or Cleomaceae or another family). Collectively our results not only help within the classification of species but may distinguish species limitations, flagging of modern species, and genus delimitation.
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Abbreviations
- ISSRs:
-
Inter-simple sequence repeats
- SRAP:
-
Sequence-related amplified polymorphism
- ORFs:
-
Open reading frames
- ISTR:
-
Inverse sequence-tagged repeat
- TWINSPAN:
-
A hierarchical dichotomous analysis
- PIC:
-
Polymorphic information content
- MI:
-
Marker index
- PCA:
-
Principal coordinates analysis
- HCA:
-
Heatmap cluster analysis
References
Gepts P (1995) Genetic markers and core collections. In: International Plant Genetic Resources Institute (IPGRI) (ed) Core collections of plant genetic resources, pp 127–146
Hofmeister W (1851) Vergleichende untersuchungen der keimung, entfaltung und fruchtbildung höherer kryptogamen:(Moose, Farrn, Equisetaceen, Rhizocarpeen und Lycopodiaceen) und der samenbildung der coniferen. The families of flowering plants - I. Macmillan & Co. Ltd, London
Hutchinson J (1926) The families of flowering plants. Dicotyledonae. Clarendon Press, London
Hutchinson J (1967) The genera of flowering plants. Clarendon Press, Oxford
Kaer A, Malver O, El-Menshawi B, Reischt J (1979) Isothiocyanates in myrosinase-treated seed extracts of Moringa peregrina. Phytochemistry 18(9):1485–1487
Tackholm V (1974) Students flora of Egypt. Cairo University Cairo
Hedge IC, Kjaeer A, Malver O (1980) Dipterygium: Cruciferae or Capparaceae. Royal Botanic Garden Edimberg 38(2):247–250
Boulos L (1999) Flora of Egypt 1st. Vol. 1. Cairo, Egypt: Al Hadara Publishing; p. 170. deserts of Ladakh. Curr Sci 96:1563–1564
Zahran MA, Willis AJ (1992) The vegetation of Egypt. Chapman and Hall, London UK
Abd El-Ghani MM, Marei AH (2006) Vegetation associates of the endangered Randonia africana Coss. and its soil characteristics in an arid desert ecosystems of western Egypt. Acta Bot Croat 65(1):83–99
Boulos L, El-Hadidi MM (1984) The weed flora of Egypt. The American university in Cairo Press Cairo
El Gohary IH (1997) Biosystematic studies of Cleomaceae in Egypt.1. the seed morphology and its taxonomic significance. Desert Inst Bull Egypt 47(2):423–440
El Gohary IH (1982) Morphological studies on the Capparaceae in Egypt MSc thesis, Botany Department, Faculty of Science, Ain shams University
Khafagi A, Al-Gohary IH (1998) Biosystematic studies of Cleomaceae in Egypt. II. Taxonomic significance of some micromorphological characters of the leaf and pollen grains. Al-Azhar Bull Sci 19(2):1027–1036
El-Karemy ZAR (2001) Capparaceae in the flora of Egypt. Taeckholmia 21(2):257–267
Zohary M (1966) Flora Palaestina, vol 1. Israel Academy of Sciences and Humanities, Jerusalem
Hedge IC, Lamond J (1970) Capparidaceae. In Rechinger K.H. ed., Flora Iranica 68:1-9 Graz
Mabberley DJ (1987) The plant book. Cambridge University Press, Cambridge, New York
Lawg (1999) Manual of leaf architecture-morphological description and categorization of dicotyledonous and net-veined monocotyledonomous angiosperms. Leaf Architecture Working Group. Washington
Stech M, Quandt D (2014) 20,000 species and five key markers: the status of molecular bryophyte phylogenetics. Phytotaxa 9(1):196–228
Liu MZ, Chen NF, Liu XZ, Deng H, Li YT, He XL (2009) Molecular authentication of Dendrobium huoshanense from its allied species. J Biol 26(5):34–36
Wang X (2010) Inter-simple sequence repeats (ISSR) molecular fingerprinting markers for authenticating the genuine species of rhubarb. J Med Plant Res 5(5):758–764
Cao PJ, Yao QF, Ding BY, Zeng HV, Zhong YX, Fu CX, Jin XF (2006) Genetic diversity of Sinojackia dolichocarpa (Styracaceae), a species endangered and endemic to China, detected by inter-simple sequence repeat (ISSR). Biochem Syst Ecol 34:231–239
Syamkumar S, Sasikumar B (2007) Molecular marker based genetic diversity analysis of Curcuma species from India. Sci Hortic 112:235–241
Xia T, Chen S, Chen S, Zhang D, Zhan D, Gao Q, Ge U (2007) ISSR analysis of genetic diversity of the Qinghai-Tibet Plateau endemic Rhodiola chrysanthemifolia (Crassulaceae). Biochem Syst Ecol 35:209–214
Li G, Quiros CF (2001) Sequence-related amplified polymorphism (SRAP), a new marker system based on a simple PCR reaction: its application to mapping and gene tagging in Brassica. Theor Appl Genet 103:455–461
Amar M, El-Zayat M (2017) Utilization of ISTR, ISSR and SRAP molecular markers to reveal and classify Egyptian pomegranates (Punica granatum L). POJ 10(05):237–246
Rhode W (1996) Inverse sequence-tagged repeat (ISTR) analysis. A novel and universal PCR (polymerase chain reaction)-based technique for genome analysis in plant and animal kingdom. J Genet Breed 50:249–261
Torres MMI, Almaraz AN, Escoto DM (2012) Review article: ISTR, a retrotransposons-based marker to assess plant genome variability with special emphasis in the genera zea and agave. Am J Plant Sci 3(12A):1820–1826
Amar M, El-Zayat M (2013) Retro transposon-markers: an overview of the recent progress in Citrus germplasm. J Biodiv Env Sci 3(10):31–41
Kalendar R (2011) The use of retrotransposon-based molecular markers to analyze genetic diversity. Field Veg Crop Res 48(2):261–274
Velascoramirez AP, Torresmoran MI, Molinamoret S, Sanchezgonzalez JD, Santacruzruvalcaba F (2014) Efficiency of RAPD, ISSR, AFLP and ISTR markers for the detection of polymorphisms and genetic relationships in camote de cerro (Dioscorea spp.). Electron J Biotechnol 17:65–71
Scariot V, De Keyser E, Handa T, De Riek J (2007) Comparative study of the discriminating capacity and effectiveness of AFLP, STMS and EST markers in assessing genetic relationships among evergreen azaleas. Plant Breed 126:207–212
Hammer HDAT, Ryan PD (2001) PAST: Paleontological statistics software package for education and data analysis. Palaeontol Electron 4(1):9
Sankar AA, Moore GA (2001) Evolution of inter simple sequence repeat analysis from mapping in Citrus and extension of the genetic linkage map. Theor Appl Genet 102:206–214
Aga E, Bryngelsson T (2005) Inverse sequence-tagged repeat (ISTR) analysis of genetic variability in forest coffee (Coffea arabica L.) from Ethiopia. Genet Resour Crop Ev 53:721–728
Powell W, Morgante M, Andre C, Hanafey M, Vogel J, Tingey S, Ralfski A (1996) Comparison of RFLP, RAPD, AFLP and SSR (microsatellite) markers for germplasm analysis. Mol Breed 2:225–238
Roldán-Ruiz I, Dendauw J, Van Bockstaele E, Depicker A, DeLoose M (2000) AFLP markers reveal high polymorphic rates in ryegrasses (Lolium spp.). Mol Breed 6:125–134
Metsalu T, Vilo J (2015) ClustVis: A web tool for visualizing clustering of multivariate data using principal component analysis and heatmap. Nucleic Acids Res 43:W566–W570
Kamel W, Abd El Ghani M, El-Bous M (2010) Cleomeaceae as a distinct family in the flora of Egypt. Afr J Plant Sci Biotechnol 4(1):11–16
Ettingshausen CV (1861) Die blatt-skelette der dikotyledonen, mit besonderer Ru cksicht auf die Untersuchung und Bestimmung der fossilen Pflanzenreste. Kais. Kon. Hof- und Staatsdr, Wien
Hickey JL (1973) Classification of the architecture of dicotyledonous leaves. Amer J Bot 60(1):17–33
Dilcher DL (1974) Approaches to the identification of angiosperm leaf remains. Bot Rev 4:1–157
Ll H, Wolfe 1A (1975) The bases of angiosperm phylogeny: vegetative morphology. Ann Mo Bot Gard 62:538–589
Banerjee G, Deshpande BD (1973) Foliar venetion and leaf histology of certain members of Compositae. Flora 162:529–532
Inamdar JA, Murthy GSR (1978) Leaf architecture in some Solanaceae. Flora 176:269–272
Jain DK (1978) Studies in Bignoniaceae. III. Leaf architecture. J Indian Bot Soc 57:369–386
Li HM, Hickey LJ (1988) Leaf architecture and systematics of the Hamamelidaceae sensu lato. Acta Phytotaxonomica Sinica 26:96–110
Sun H, Chen J, Zhou Z, Fei Y (1991) The leaf architecture and its taxonomic significance in the genera Albizia and Cylindrokelupha from China. Acta Bot Yunnanica 13:241–253
Rao SS, Narmada K (1994) Leaf architecture in some Amaranthaceae. Feddes Repertorium 105(1-2):37–44
Wang YF, Ferguso DK, Zetter R, Denk T, Garp G (2001) Leaf architecture and epidermal characters in Zelkova (Ulmaceae). Bot J Linn Soc 136:255–265
Luo Y, Zhou ZK (2002) Leaf architecture in Quercus subgenus Cyclobalanopsis (Fagaceae) from China. Bot J Linn Soc 140:283–295
Inamdar JA, Shenoy KN, Rao NV (1983) Leaf architecture of some monocotyledons with reticulate venation. Ann Bot 52:725–735
Moubasher H, Abd El-Ghani M, Kamel W, Mansi M, El-Bous M (2011) Taxonomic considerations among and within some Egyptian taxa of Capparis and related genera (Capparaceae) as revealed by RAPD fingerprinting. Collect Bot 30:29–35. https://doi.org/10.3989/collectbot.v30.003
Zhang QD, Jia RZ, Meng C, Ti CW, Wang YL (2015) Diversity and population structure of a dominant deciduous tree based on morphological and genetic data. AoB Plants:7
Abdel-Mawgood A, Ahmed M, Ali B (2006) Application of molecular markers for hybrid maize (Zea mays L.) identification. J Food Agric Env 4:176–178
Inocencio A, Kikuchi M, Merrey D, Tonosaki M, Maruyama A, de Jong I, de Vries FP (2005) Lessons from irrigation investment experiences: cost-reducing and performance-enhancing options for sub-Saharan Africa. International Water Management Institute. Final Report, Battaramulla, Sri Lanka
Belaj AZ, Satovic G, Cipriani L, Baldoni R, Testolin L, Rallo I, Trujillo (2003) Comparative study of the discriminating capacity of RAPD AFLP and SSR markers and of their effectiveness in establishing genetic relationships in olive. Theor Appl Genet 107:736–744
Amar MH, El Wahab MA (2013) Comparative genetic study among Origanum L. plants grown in Egypt. J Biol Environ Sci 3:208–222
Biswas MK, Xu Q, Deng XX (2010) Utility of RAPD, ISSR, IRAP and REMAP markers for the genetic analysis of Citrus spp. Sci Hortic 124:254–261
Du D, Du X, Mattia MR, Wang Y, Yu Q, Huang M, Yu Y, Grosser JW, Gmitter FG (2018) LTR retrotransposons from the Citrus x clementina genome: characterization and application. Tree Genet Genomes 14:43
Kalendar R, Schulman AH (2014) Transposon-based tagging: IRAP, REMAP, and iPBS. In Molecular plant taxonomy. Humana Press, Totowa NJ, pp 233–255
Inocencio C, Rivera D, Obon C, Alcaraz F, Barrena JA (2006) A systematic revision of Capparis section Capparis (Capparaceae). Ann Missouri Bot Gard 93:122–149
Al-Safadi B, Faouri H, Elias R (2014) Genetic diversity of some Capparis L. species growing in Syria. Braz Arch Biol Technol 657:916–926
Fici S (2014) A taxonomic revision of the Capparis spinosa group (Capparaceae) from the Mediterranean to Central Asia. Phytotaxa 174(1):001–024
Tan K (2002) Capparaceae. In: Strid A. & Tan, K. (Eds.) Flora Hellenica. 2. A. R. G. Gantner Verlag K. G., Ruggell:113–115
Özbek O, Kara A (2013) Genetic variation in natural populations of Capparis from Turkey, as revealed by RAPD analysis. Plant Syst Evol 299:1911–1933
Rivera RM, Kelley KL, Erdos GW, Hansen PJ (2003) Alterations in ultrastructural morphology of two-cell bovine embryos produced in vitro and in vivo following a physiologically-relevant heat shock. Biol Reprod 69:2068–2077
Saifi N, Ibijbijen J, Echchgadda D (2011) Genetic diversity of caper plant (Capparis ssp.) from North Morocco. J Food Agric Env 9(3&4):299–304
Saadaoui E, Guetat A, Tlili N, El Gazzah M, Khaldi A (2011) Subspecific variability of Tunisian wild populations of Capparis spinosa L. J Med Plants Res 5(17):4339–4348
Nadeem MA, Nawaz MA, Shahid MQ, Doğan Y, Comertpay G, Yıldız M, Hatipoğlu R, Ahmad F, Alsaleh A, Labhane N, Özkan H (2018) DNA molecular markers in plant breeding: current status and recent advancements in genomic selection and genome editing. Biotechnol Biotechnol Equip 32(2):261–285
Acknowledgements
This work is under the activity of the Egyptian Desert Gene Bank, Desert Research Center, Egypt. The authors are thankful to the late Professor Dr. Ahmed Morsy, professor of plant ecology, Desert Research Center, Egypt, for support and advice.
Funding
The authors declare no funding for this research.
Author information
Authors and Affiliations
Contributions
Both MSA and MAZ contributed equally to this paper. MSA formulated the idea, designed, and performed the ecological experiments, while MAZ carried out the practical work of genetic diversity experiments. MHA analyzed the data and wrote the first draft of the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
El zayat, M.A.S., Ali, M.E.S. & Amar, M.H. A systematic revision of Capparaceae and Cleomaceae in Egypt: an evaluation of the generic delimitations of Capparis and Cleome using ecological and genetic diversity. J Genet Eng Biotechnol 18, 58 (2020). https://doi.org/10.1186/s43141-020-00069-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s43141-020-00069-z