
Abstract
Background
Mediterranean-type ecosystems are fire-prone environments where species have evolved in the presence of seasonal summer conditions and frequent fires. In these environments, many species have seeds with impermeable hard coats that impose physical seed dormancy which prevents seed germination until it is broken by any factor such as fire or high summer temperatures. Most studies have usually focused on the isolated effects of fire or summer temperatures on the release of physical seed dormancy. However, both factors frequently operate together because when a fire occurs, usually in summer, seeds in the soil suffer not only the heat shock during the fire, but also the high summer temperatures. Nevertheless, the combined effect of both factors is almost unknown. In this investigation, we studied the isolated and combined effects of heat shock and the high temperatures of summer. The main objective was to evaluate whether the timing in which seeds are exposed to a heat shock during the period of high summer temperatures affects seed germination.
Results
We exposed the seeds of 12 Cistaceae species to three different treatments, in which heat shock (100 °C for 10 min) was applied before, after, or before + after they were exposed to summer temperatures (50/20 °C for 1 month). Although fire was the main factor breaking physical dormancy, its effects were regulated by the timing when it was simulated. We found that summer temperatures following heat shock had negative effects on seed germination since a high proportion of seeds remained dormant. On the contrary, when summer temperatures preceded heat shock, physical dormancy was broken, and germination increased.
Conclusions
Timing of fire during summer plays an important role in controlling seed germination of Cistaceae. Our results provide a novel explanation for the higher Cistaceae seedling emergence found in the field after late season fires. Additionally, we can carefully draw conclusions for fire management since prescribed burnings would have different effects depending on when they were carried out.
Resumen
Antecedentes
Los ecosistemas mediterráneos son ambientes proclives al fuego, en los cuales las especies han evolucionado bajo la presencia de condiciones estacionales en verano y fuegos frecuentes. En estos ambientes, muchas especies tienen semillas con una cubierta dura e impermeable que les impone una dormancia física que impide su germinación hasta que es rota por algún factor como el fuego o las altas temperaturas del verano. La mayoría de los estudios s han enfocado normalmente en los efectos aislados del fuego o las temperaturas del verano en la liberación de la dormancia física de las semillas. Sin embargo, frecuentemente ambos factores operan juntos dado que cuando un fuego ocurre, usualmente en verano, las semillas en el suelo sufren no sólo el choque térmico sino también las altas temperaturas del verano. A pesar de ello, el efecto conjunto de ambos factores es prácticamente desconocido. En esta investigación, estudiamos los efectos aislados y conjuntos del choque térmico del fuego y las altas temperaturas del verano. El objetivo principal fue evaluar si el momento en el que las semillas son expuestas al choque térmico durante el período de altas temperaturas del verano afecta su germinación.
Resultados
Expusimos las semillas de 12 especies de Cistáceas a tres tratamientos diferentes, en los cuales el choque térmico (100 °C durante 10 min) fue aplicado antes, después, o antes + después de que fueran expuestas a temperaturas de verano (50/20 °C durante un mes). Aunque el fuego fue el factor principal en romper la dormancia física, sus efectos fueron regulados por el momento en el cual éste fue simulado. Encontramos que las temperaturas de verano posteriores al choque térmico tuvieron un efecto negativo en la germinación de semillas, dado que una gran proporción permanecieron latentes. Por el contrario, cuando las temperaturas de verano precedieron al choque térmico la dormancia física se rompió y la germinación aumentó.
Conclusiones
El momento en el que ocurren los incendios durante el verano juega un papel importante en la germinación de las Cistáceas. Nuestros resultados proporcionan una explicación novedosa para la mayor emergencia de las Cistáceas encontrada en el campo tras los incendios tardíos del verano. Adicionalmente, podemos extraer conclusiones con prudencia sobre el uso del fuego, dado que las quemas prescritas pueden tener diferentes efectos dependiendo de cuándo éstas sean llevadas a cabo.
Similar content being viewed by others


Introduction
Fire and the Mediterranean climate have coexisted since the onset of this type of climate in the Neogene-Quaternary (Suc 1984; Hernández-Fernández et al. 2007). High summer temperatures together with the absence of precipitation during this season lead to a regime of recurrent fires (Rundel et al. 2018). Consequently, plants have evolved facing both factors with traits that help them to survive and regenerate in these environments (Trabaud 1987; Keeley et al. 2011a). Hardseededness is one of these traits frequently found among the Mediterranean plant species (Baskin and Baskin 2014). Hardseededness imposes physical dormancy (PY) that prevents germination until it is released by a factor such as fire (Moreira and Pausas 2012) or high summer temperatures (Santana et al. 2010, 2012). Nowadays, there is debate about the origin of this trait (Bradshaw et al. 2010; Keeley et al. 2011b; Jaganathan 2015), and while some authors point to fire (Moreira and Pausas 2012) others point to climate (Santana et al. 2020) as the selective pressures causing PY in seeds. Probably, both evolutionary forces were involved in the selection of this character, but this has yet to be explored.
Cistaceae, one of the 18 angiosperm plant families with PY (Baskin and Baskin 2014), is widely distributed in the Mediterranean Basin where it forms vast extensions of shrubland in open dry sunny habitats, especially after fire occurrence (Allen 2009). This plant family occupies poor and acidic soils, notably in the western part of the Mediterranean region (Rivas-Martínez 1979). Although fire plays an important ecological role in Cistaceae colonization (Trabaud 1995), it is not exclusively dependent on fire for its regeneration. Consequently, Cistaceae plants are considered not as typical pyrophytes but as opportunistic species since they colonize not exclusively burned areas but also abandoned agricultural lands, perturbed woodlands, or other types of degraded areas (Trabaud 1995). This is possible because these species show ample germination ranges, being able to germinate under a wide range of environmental conditions once PY is broken (Thanos and Georghiou 1988; Thanos et al. 1992; Baskin et al. 2000). Despite this broad tolerance to germinate under varied conditions, different factors such as temperature, light and water appear to regulate seed germination after PY release (Luna and Chamorro 2016; Luna et al. 2022).
Fire has been the most studied environmental stimulus in Cistaceae species, and investigations on Cistaceae germination have focused mainly on the effects of heat shock. A high variability of responses to heat exposure has been described, which may be explained as a mechanism of diversification in relation to the different fire intensities experienced by seeds on the soil surface, as well as variations in burial depth (Luna et al. 2019). The response to other fire cues, such as smoke, is not so clear. In fire-prone habitats, many species require smoke to germinate (Keeley and Fotheringham 1998; Çatav et al. 2014; Mackenzie et al. 2016; Moreira and Pausas 2018). Nevertheless, different studies indicate that smoke does not increase germination in species with physical dormancy (including Cistus albidus, C. clusii, C. ladanifer, C. laurifolius, C. monspeliensis, and C. salviifolius) (Reyes and Trabaud 2009; Çatav et al. 2018; Chamorro and Moreno 2019). However, this lack of response to smoke is not always consistent, e.g., Martínez-Baniela et al. (2016) found that Helianthemum tinetense showed physiological seed dormancy released by smoke but not other Helianthemum species.
Physical dormancy is present in over 25% of plant species, especially in ecosystems subject to temporally stochastic events, such as fire and drought (Hudson et al. 2015). Seeds with PY cannot germinate under favorable temperature and water conditions for germination because the palisade cell layer(s) impedes seed imbibition. During dormancy break, an opening is formed in the seed coat through the water-impermeable layer(s) at a specialized morphoanatomical site, usually called a water gap (Gama-Arachchige et al. 2013). Cistaceae shows a chalazal plug that is pushed slightly into the seed and forms a circular opening (oculus) through which water goes into the seed (Baskin et al. 2000; Gama-Arachchige et al. 2013; Geneve et al. 2018). Once PY is broken, seeds cannot revert to the dormant stage (Baskin 2003) and thus, fine tuning to environmental cues must exist to ensure seed survival.
In addition to fire or summer temperatures, other environmental cues can break PY, like temperature fluctuations, microbial action, or animal ingestion among others (Baskin and Baskin 2014). In the case of Cistaceae, the break of PY has usually been related to high temperatures reached during fire as evidenced by the high colonization which happens after fire (Arianoutsou and Margaris 1981; Moreno et al. 2011). However, although temperature thresholds needed for breaking PY have been traditionally related to fire (Herranz et al. 1999), lower temperature thresholds related to summer temperatures can be sufficient to break seed dormancy in some cases (Santana et al. 2012) but not in others (Moreira and Pausas 2012; Luna 2020). In Mediterranean habitats, seed dispersal of Cistaceae occurs frequently during the summer (Bastida and Talavera 2002). Likewise, in these Mediterranean environments, fires occur mainly during the summer season when high temperatures and low humidity favor fire propagation (Vázquez and Moreno 1998; Vecín-Arias et al. 2016). Consequently, dispersed seeds have to withstand the high soil temperatures during summer and eventually the higher temperatures of fire. Despite both factors usually acting together, to our knowledge, only two papers have analyzed their joint effects (Luna 2020; Zomer et al. 2022). Zomer et al. (2022) found that seeds from populations growing at locations with higher summer temperatures showed higher heat thresholds to break PY than those populations from locations with lower summer temperatures. In addition, a novel negative effect of summer temperatures emerged for the germination of 12 Cistaceae species, which disappeared to some extent when heat shock followed summer (Luna 2020).
In the present study, we analyze in detail how both factors, fire and high summer temperatures, regulate germination rates. The main objective was to evaluate whether the timing in which seeds are exposed to a short intense heat shock representative of fire during the period of high summer temperatures affects seed germination. Specifically, we were interested in comparing the effects of heat shock and high summer temperatures on PY break and the effects of heat shock produced before, after, and both before and after the high summer temperatures on seed germination traits.
If the timing of fire determines PY break and subsequent germination, important lessons could be learned for effective fuel management practices such as prescribed burnings. Wildfires are an increasingly serious risk in Mediterranean areas whereby different management techniques are applied to reduce fuel loads and consequently to decrease the fire risk (Fernandes and Botelho 2003). Most of the prescribed burnings are applied in southern Europe for fire hazard reduction, but also can be used for biodiversity conservation (Fernandes et al. 2013). As a consequence of fire application, some species can thrive in association with the release of PY due to fire or to the opening of clearings. More in-depth knowledge of the regeneration of the species will help to give guidelines for landscape management, either avoiding the expansion of some species or promoting their appearance and maintenance.
Material and methods
We studied the germination ecology of 12 common Cistaceae species widely distributed in the western Mediterranean: Cistus albidus L., Cistus clusii Dunnal, Cistus ladanifer L., Cistus laurifolius L., Cistus monspeliensis L., Cistus populifolius L., Cistus psilosepalus Sweet, Cistus salviifolius L., Halimium atriplicifolium (Lam.) Spach, Halimium halimifolium (L.) Willk, Halimium ocymoides (Lam.) Willk and Halimium viscosum (Lam.) Spach. Mature capsules were harvested during dispersal from at least 30 individuals for each species between July and August 2016 in the center of the Iberian Peninsula (Supplementary Table S1). Then, in the laboratory seeds were cleaned and separated in sets of 25 seeds in paper bags. Seeds were stored at dark in laboratory conditions until the beginning of the experiments.
The timing of fire and the influence of summer temperature were analyzed in a factorial experiment. Fire was simulated by exposing seeds to a heat shock of 100 °C during 10 min in an air-forced oven. This temperature can be reached both in high as well as moderate intensity fires depending on the position in the soil profile. We chose this specific temperature and time of exposure because it coincides with the records commonly found in the Mediterranean fire shrublands (Moreno et al. 2011; Céspedes et al. 2012) and it was the optimal for breaking the seed dormancy of many Cistaceae species (Luna et al. 2019). Summer temperatures were simulated by exposing unimbibed seeds to daily cycles of 12 h to alternating temperatures 50/20 °C for 1 month in a refrigerated precision cabinet (JP Selecta Hotcold-UB). When air temperatures are around 20–25 °C, soil temperatures are similar but, when the mean of maximum temperatures is around 35 °C, soil surface temperature can reach 60 °C (Ooi et al. 2009). In the Mediterranean, air summer temperatures are usually above 40 °C and daily maximum temperatures recorded in soil can be very high, up to 50 °C on the soil surface of fire breaks in eastern Spain (Baeza and Roy 2008) or up to 40–60 °C in south-eastern Australia (Tieu et al. 2001) or 60–70 °C in south-west western Australia (Hnatiuk and Hopkins 1981). Thus, it is common to use this range of temperatures to simulate current summer temperatures. Furthermore, both temperatures and duration of the treatment applied in this investigation have been widely used in other studies (Santana et al. 2010; Ooi et al. 2012, 2014; Cochrane 2017). In a recent investigation, the effects of summer length on seed germination were studied for the same species analyzed in the present article. No differences between 1 and 2 months of exposure to summer emerged, so we chose the shortest length.
Heat shock was produced before seeds were exposed to summer temperatures (HB), after summer temperatures (HA), and before + after summer temperatures (HBA). A control without heat shock was also included in the experiment (H −). Additionally, a control without exposing seeds to summer temperatures was added for each treatment. That is, while seeds were exposed to summer temperatures, seeds of the control treatments for summer were stored under laboratory conditions, with temperatures of around 20 °C (Fig. 1).

Diagram of the experimental design. The effects of summer temperatures and the timing of heat shock were analyzed in a factorial experiment. Seeds were exposed to a heat shock (100 °C for 10 min) before, after, and before + after a month of exposure to summer temperatures (50/20 °C) together with a control without heat shock. Additionally, a control without exposing seeds to summer temperatures was added for each treatment by repeating the experiment at the same time but under laboratory conditions (around 20 °C) instead of summer temperatures
After these treatments, the seeds were put to germinate on two sheets of filter paper moistened with 1.2 ml of distilled water, in plastic Petri dishes of 5.5 cm diameter. For germination tests, four replicates of 25 seeds were used for each species and treatment. Petri dishes were laid at random in a temperature and humidity-controlled chamber (Model G-21, Ibercex), where seeds were incubated for 6 weeks at 20 °C and a 12 h photoperiod, the optimal germination conditions for many Mediterranean species (Thanos and Georghiou 1988; Luna et al. 2012). Germination was checked daily during the first week and 3 days a week during the last 5 weeks of the experiment. At the end of the experiment, ungerminated seeds were checked for viability by tetrazolium test (Moore 1985).
Four variables were obtained: final germination percentage at the end of the experiment corrected for viability (i.e., germination percentages were estimated in relation to viable seeds), germination speed characterized by the number of days to initiate germination (T0) and to produce 50% of the total germination obtained (T50), and finally, seed viability (i.e., viability percentages were assessed by considering germinated seeds plus non-germinated but tetrazolium colored seeds).
Data were analyzed with generalized linear models (GLMs). Based on the error structure, we used a binomial error distribution and logit link function for final germination and seed viability. In the case of T0 and T50, a Poisson error distribution with log link function was considered most appropriate in relation to the data. We tested for the effects of summer temperatures (two levels, without and with summer temperatures) and timing of heat shock (four levels, without heat shock, before summer, after summer, before + after summer). When timing of heat shock had significant effects, pair-wise comparisons among treatments were performed using the Bonferroni correction. All statistical analyses were performed using the SPSS Statistics version 24.0 (SPSS, Chicago, IL, USA).
Results
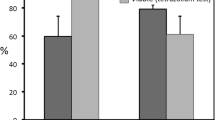
Under control conditions, i.e., seeds were not exposed to summer temperatures, heat shock increased germination of all species whatever the timing of heat shock was (Figs. 2 and 3). When seeds were incubated at summer temperatures, heat shock also increased germination of many species. However, in other species, C. clusii, C. monspeliensis, C. psilosepalus, and the four Halimium species, the heat shock had no effect when it was produced before summer temperatures. Additionally, for most species (except C. populifolius and C. salviifolius), post-summer heat shock (HA) had a significant positive effect on seed germination in comparison to pre-summer heat shock (HB). Finally, for most species, no germination differences emerged between heat shock produced after (HA) and before + after summer temperatures treatments (HBA) (Figs. 2 and 3). Moreover, germination after the two heat shocks (HBA) increased in relation to the lower germination percentages observed after pre-summer heat shock treatment (HB), except for C. albidus, C. laurifolius, C. populifolius, and C. salviifolius.

Germination percentages (mean ± standard error) of Cistus species after being stored under laboratory control conditions (Control) and at summer temperatures (50/20 °C) (Summer). Seeds were exposed to a heat shock before summer temperatures (or lab conditions) (HB), after summer temperatures (HA), and before + after summer temperatures (HBA). A control without heat shock was also included in the experiment (H −). Different letters show significant differences for seed germination (P < 0.05) between heat shock treatments for seeds exposed to summer and control laboratory conditions, based on pairwise comparisons with the Bonferroni correction after GLM analysis

Germination percentages (mean ± standard error) of Halimium species after being stored under laboratory control conditions (Control) and at summer temperatures (50/20 °C) (Summer). Seeds were exposed to a heat shock before summer temperatures (or lab conditions) (HB), after summer temperatures (HA), and before + after summer temperatures (HBA). A control without heat shock was also included in the experiment (H −). Different letters show significant differences for seed germination (P < 0.05) between heat shock treatments for seeds exposed to summer and control laboratory conditions, based on pairwise comparisons with the Bonferroni correction after GLM analysis
Summer temperatures had the opposite effect to heat shock, decreasing germination except in the cases of C. ladanifer, C. laurifolius, and C. monspeliensis, which were not affected (Table 1, Figs. 2 and 3). A significant interaction emerged between heat shock and summer temperatures for more than half of the species (Table 1), which means that the germination response to timing of heat shock was different when seeds went through summer temperatures in relation to the control. In most of these cases, heat shock before summer (HB) did not break PY leading to similar low germination percentages than control treatment (H −) contrary to results under lab control conditions (Figs. 2 and 3).
Summer temperatures increased T0 and T50 (Figs. S1–S4), that is, delayed the onset of germination and decreased germination speed in most species (Table 2). On the contrary, timing of heat shock hardly had effects on T0 and T50 (Table 2, Supplementary Table S2). A significant interaction also emerged between heat shock and summer temperatures for seven species (Supplementary Table S2). Cistus ladanifer was the species with the lowest values of T0 and T50 (fastest germination rates) while Halimium viscosum was the species with the highest ones (Table 2).
Overall, seed viability was high, hovering around 80%, except for C. populifolius, which did not reach 30% (Figs. 4, 5, and S5). Seed viability of seven species decreased after the double heat shock (HBA) in relation to the unheated seeds (H −) under control conditions, though not so in those stored under summer temperatures (Supplementary Figs. S5–S6). C. psilosepalus, C. salviifolius, and H. viscosum were quite sensitive to the double heat shock, showing a significant viability decrease in relation to the unheated seeds and to those exposed to only one heat shock (Supplementary Figs. S5–S6).

Stacked proportion of germinated and ungerminated viable seeds (mean and standard errors) are shown in white and colored bars respectively for the eight Cistus species studied and the different heat shock treatments: H − (without heat shock), HB (heat shock before one month of storage), HA (heat shock after one month of storage), HBA (heat shock before + after storage)

Stacked proportion of germinated and ungerminated viable seeds (mean and standard errors) are shown in colored and gray bars respectively for the Halimium species studied and the different heat shock treatments: H − (without heat shock), HB (heat shock before one month of storage), HA (heat shock after one month of storage), HBA (heat shock before + after storage)
For most species, a proportion of viable seeds remained ungerminated in all treatments (Figs. 4 and 5). C. ladanifer, C. laurifolius, and C. psilosepalus showed the highest proportion of seeds germinated without heat shock, around 30% when seeds stayed under laboratory control conditions. When seeds had been kept under summer temperatures, the highest proportion of ungerminated viable seeds was reached both in the control without heat shock (H −) and in the pre-summer heat shock treatment (HB) (Figs. 4 and 5).
Discussion
Fire was the main factor breaking PY since heat shock alone was enough to break dormancy if it was followed by seed watering. On the contrary, summer temperatures did not break PY and even had a negative effect on seed germination. The timing of heat shock, before or after high summer temperatures, determined seed germination in most species. When seeds were exposed to heat shock before being stored under summer temperatures germination was lower than when they were exposed to heat shock after summer. The loss of seed germination after pre-summer heat treatment was not associated with seed viability loss but with an increase in the proportion of hard-coated seeds. This could be explained by the acquisition of secondary physical dormancy, that is, the ungerminated seeds would become hard again after the lack of primary physical seed dormancy (Rolston 1978; Lamont et al. 2022). However, since it is widely considered that the process of PY loss is not reversible (Baskin 2003; Baskin and Baskin 2014), the lack of germination in HB treatment should be explained by a cycling of sensitivity to physical dormancy break. Traditionally, PY break is considered as a simple physical event, in which PY is released directly by one factor, in the case of Cistaceae, by the opening of the water gap in the chalaza region in response to an environmental cue such as high or fluctuating temperatures (Baskin et al. 2000; Van Assche et al. 2003; Jayasuriya et al. 2008a). However, in different cases, loss of PY can occur in two stages, which evidences that environmental control of the germination of seeds with water-impermeable seed coats may be much more complex (Taylor 1981; Jayasuriya et al. 2009). This mechanism has been described in species of Convolvulaceae and Fabaceae (Jayasuriya et al. 2008a, 2008b; Rodrigues-Junior et al. 2018) and proposed for Cistaceae (Luna 2020). According to this model, dormant seeds can cycle between insensitive (hard seeds which are not sensitive to a dormancy-breaking treatment) and sensitive (latent soft seeds, which are sensitive to a dormancy-breaking treatment) stages (Taylor 2005). PY break occurs in two steps, the first is a pre-conditioning step that softens the seeds and makes them sensitive to dormancy breaking, and the second, the actual dormancy-breaking step, during which seeds imbibe water (Baskin and Baskin 2014; Jaganathan 2016). In the case of Cistaceae, fire would be the first requirement for breaking PY but, after fire, favorable conditions would be needed for germination completion (Fig. 6). In the present study, Cistus and Halimium seeds did not complete PY break if after the heat shock they were not imbibed but kept under inappropriate conditions for germination, such as is shown in Fig. 6. Furthermore, HBA had similar effects to HA, which suggest that the negative effect of HB on seed germination can be reverted. This means that the high proportion of hard-coated seeds found in HB would decrease after an additional heat shock, which supports the model of sensitivity cycling to break PY. That is, insensitive seeds could be sensitive again after a new heat shock (Fig. 6). Future investigations should address the examination of seeds after the different treatments by electron microscopy to confirm these arguments.

Diagram of sensitivity cycling to physical dormancy (PY) release for studied Cistaceae species. According to this model, PY break occurs in two steps, the first is a pre-conditioning step that softens the seeds and makes them sensitive to dormancy breaking, and the second, the actual dormancy-breaking step, during which seeds imbibe water. Seeds would not complete PY break if after the heat shock they are kept under inappropriate conditions for germination such as high summer temperatures. That is, the second step of PY break would not have been completed. A generalized model of dormancy-breaking requirements and sensitivity cycling in seeds with PY can be reviewed in Jayasuriya et al. (2009)
In some cases, we found in the present study that under lab control conditions seed germination, when heat shock was produced one month before seeds were moistened (HB), was lower than when seeds were immediately moistened after heat shock (HA). The germination decrease in HB and control, supports the notion that seed dormancy break may not be completed if a long time passes before seed imbibition. However, this lack of germination was not as intense or consistent among the species as under the summer treatment, which evidences that not only time but also conditions present during the time elapsed between heat shock and imbibition must determine completion of PY break.
Around the Mediterranean, fires commonly take place in summer (Vázquez and Moreno 1998; Levin and Saaroni 1999). In the Iberian Peninsula, lightning-caused fires occur mainly from June to September with a peak during July and August (Vázquez and Moreno 1998; Vecín-Arias et al. 2016). In these areas, germination occurs as soon as soils have appropriate moisture, usually following the first autumn rains (Espigares and Peco 1993; Traba et al. 2006; Céspedes et al. 2012). Consequently, the timing of fire, early or late in the summer may determine the degree of physical dormancy release. After early season fires in the summer, seeds would not moisten and would not complete physical dormancy break, contrary to seeds exposed to late season fires, which in a short time would probably find the wet conditions needed to break dormancy and germinate. However, sporadic rains during summer storms could perhaps manage to complete the release of PY after early season fires although not to complete germination because of inappropriate conditions for it (Luna and Chamorro 2016). Germination of seeds may be affected by the time passed between fire and the arrival of the rains (Daskalakou and Thanos 2004; Ooi et al. 2004). Future investigations should be aimed at identifying the threshold time elapse between heat shock and seed imbibition to complete physical seed dormancy break.
Overall, in Mediterranean-type climates, late season fires result in higher seedling emergence than early season ones (Papanastasis 1980; Bond 1984; Midgley 1989; Grant and Loneragan 1999). Two arguments are frequently used to explain this higher germination after late season fires than after early season fires: fire intensity and phenology. The fire season may influence the moisture content of fuels and therefore the intensity of a fire (De Luis et al. 2004). Late season fires are expected to be more severe than early ones, because they occur at the end of the dry season, when the water content of vegetation, dead fuel, and soil is low (Neary et al. 1999). Consequently, since fire intensity can regulate the dormant proportion of seed banks for species with physical seed dormancy (Parker and Kelly 1989), the higher temperatures reached during late season fires would lead to a greater release of physical seed dormancy and subsequent higher seedling emergence.
Furthermore, although fire intensity is similar among fires, season also affects population dynamics of species (Bond and Van Wilgen 1996) due to phenological aspects. Seed dispersal in the Mediterranean often happens during summer and autumn (Bastida and Talavera 2002; Traba et al. 2006). Consequently, the soil seed bank is larger at the time of late season fires than early ones (Bond et al. 1984; Cowling and Lamont 1987). According to this, both in the western as well as the eastern Mediterranean Basin, Cistus germination was higher after late season fires than after early ones, despite fire intensities being similar (Céspedes et al. 2012; Tsafrir et al. 2018). Our research adds a novel interpretation for the higher germination after late season fires in comparison to early ones (Manela et al. 2022). After early season fires, a lower proportion of seeds may germinate because of lower rates of PY break completion.
When extrapolating our results to the actual situation in the countryside, we should bear in mind that laboratory studies represent a simplification of natural conditions. In this study we simplified the effect of fire and the diversity of the temperatures reached and how long they lasted in a single homogenous thermal shock, we reduced the temperatures that the soil experiences during the summer to a constant alternance of temperatures, and we limited the summer to 1 month in which there was no precipitation due to storms. Even so, in spite of the difficulty and the risks that are inherent in the extrapolation of the conclusions of laboratory experiments to explain the world around us, it is important to give careful thought to the possible implications that basic science can have to be used by those who manage the natural environment.
Our results can have implications for making decisions on effective fuel management treatments such as prescribed burnings. Prescribed burnings are increasingly used, especially as a tool for reducing fuel load to fight against increasingly severe wildfires. A balance must be pursued between conserving biodiversity and reducing fire risk. The use of fire can lead directly to the success of some species due to the PY break, but some of these species, like some Cistaceae, can be considered a problem for fuel management. As an example, Cistus ladanifer and Cistus laurifolius form monospecific shrublands that usually do not progress towards more mature stages of vegetation (Mendes et al. 2015) because of their great capacity for regeneration together with the release of phytotoxic compounds into the soil that inhibit the development of other plants (Tárrega et al. 2001; Herranz et al. 2006; Sosa et al. 2010). Additionally, these homogeneous landscapes show a high accumulation and continuity of dead fuel, which leads to situations of high flammability (Saura-Mas et al. 2010; Baeza and Santana 2015). Prescribed burnings are conducted out-of-season, usually during spring or autumn, avoiding the summer season and seeking easy conditions for fire control. Therefore, soils are frequently wet or will be wet early after burnings, which will favor the expansion of Cistaceae species. According to our findings, conducting prescribed burnings just before the summer drought could help to control the expansion of the studied species in relation to later fires followed by rains. Therefore, it is essential to choose the appropriate timing for applying managed burnings to guarantee the achievement of aims.
Conclusions
Fire and summer play a role in determining germination of studied Cistaceae, which supports the general idea that in many cases physical seed dormancy has allowed plant survival under hard and stochastic environments (Hudson et al. 2015). Fire was the main factor breaking physical dormancy, contrary to summer temperatures, which had a negative effect on seed germination. Therefore, the timing of heat shock, before or after high summer temperatures, determined seed germination in most species, and consequently, implications for fire management can be extracted.
Availability of data and materials
Most data that support this study are available in the article and accompanying online supplementary material. However, if someone needs additional information it will be shared upon reasonable request to the corresponding author.
References
Allen, H. 2009. Vegetation and ecosystem dynamics. In The physical geography of the Mediterranean, 203–27. Oxford: Oxford University Press.
Arianoutsou, M., and N.S. Margaris. 1981. Early stages of regeneration after fire in a phryganic ecosystem (East Mediterranean). I. Regeneration by Seed Germination. Biologie-Ecologie Méditerraneenne 8: 119–128.
Baeza, M.J., and J. Roy. 2008. Germination of an obligate seeder (Ulex parviflorus) and consequences for wildfire management. Forest Ecology and Management 256: 685–93.
Baeza, M.J., and V.M. Santana. 2015. Biological significance of dead biomass retention trait in Mediterranean Basin species: an analysis between different successional niches and regeneration strategies as functional groups. Plant Biology 17: 1196–1202.
Baskin, C., and J. Baskin. 2014. Seeds: Ecology, biogeography, and evolution of dormancy and germination. San Diego: Academic Press.
Baskin, C.C. 2003. Breaking physical dormancy in seeds - focussing on the lens. New Phytologist 158: 229–232.
Baskin, C.C., J.M. Baskin, and X. Li. 2000. Taxonomy, anatomy and evolution of physical dormancy in seeds. Plant Species Biology 15: 139–152.
Bastida, F., and S. Talavera. 2002. Temporal and spatial patterns of seed dispersal in two Cistus species (Cistaceae). Annals of Botany 89: 427–434.
Bond, W.J. 1984. Fire survival of Cape Proteaceae-influence of fire season and seed predators. Vegetatio 56: 65–74.
Bond, W.J., and B.W. Van Wilgen. 1996. Fire and plants. London: Chapman & Hall.
Bond, W.J., J. Vlok, and M. Viviers. 1984. Variation in seedling recruitment of Cape Proteaceae after fire. Journal of Ecology 72: 209–221.
Bradshaw, S.D., K.W. Dixon, S.D. Hopper, H. Lambers, and S.R. Turner. 2010. Little evidence for fire-adapted plant traits in Mediterranean climate regions. Trends in Plant Science 16: 69–76.
Çatav, S.S., K. Kucukakyuz, K. Akbas, and C. Tavsanoglu. 2014. Smoke-enhanced seed germination in Mediterranean Lamiaceae. Seed Science Research 24: 257–264.
Çatav, S.S., K. Kucukakyuz, C. Tavsanoglu, and J.G. Pausas. 2018. Effect of fire-derived chemicals on germination and seedling growth in Mediterranean plant species. Basic and Applied Ecology 30: 65–75.
Céspedes, B., I. Torres, B. Luna, B. Pérez, and J.M. Moreno. 2012. Soil seed bank, fire season, and temporal patterns of germination in a seeder-dominated Mediterranean shrubland. Plant Ecology 213: 383–393.
Chamorro, D., and J.M. Moreno. 2019. Effects of water stress and smoke on germination of Mediterranean shrubs with hard or soft coat seeds. Plant Ecology 220: 511–521.
Cochrane, A. 2017. Are we underestimating the impact of rising summer temperatures on dormancy loss in hard-seeded species? Australian Journal of Botany 65: 248–256.
Cowling, R.M., and B.B. Lamont. 1987. Post-fire recruitment of four co-occurring Banksia species. Journal of Applied Ecology 24: 645–648.
Daskalakou, E., and C. Thanos. 2004. Postfire regeneration of Aleppo pine – the temporal pattern of seedling recruitment. Plant Ecology 171: 81–89.
De Luis, M., M.J. Baeza, J. Raventós, and J.C. González-Hidalgo. 2004. Fuel characteristics and fire behaviour in mature Mediterranean gorse shrublands. International Journal of Wildland Fire 13: 79–87.
Espigares, T., and B. Peco. 1993. Mediterranean pasture dynamics: The role of germination. Journal of Vegetation Science 4: 189–194.
Fernandes, P.M., and H.S. Botelho. 2003. A review of prescribed burning effectiveness in fire hazard reduction. International Journal of Wildland Fire 12: 117–128.
Fernandes, P.M., G.M. Davies, D. Ascoli, C. Fernández, F. Moreira, E. Rigolot, C.R. Stoof, J.A. Vega, and D. Molina. 2013. Prescribed burning in southern Europe: Developing fire management in a dynamic landscape. Frontiers in Ecology and the Environment 11: E4–E14.
Gama-Arachchige, N.S., J.M. Baskin, R.L. Geneve, and C.C. Baskin. 2013. Identification and characterization of ten new water gaps in seeds and fruits with physical dormancy and classification of water-gap complexes. Annals of Botany 112: 69–84.
Geneve, R.L., C.C. Baskin, J.M. Baskin, K.M.G.G. Jayasuriya, and N.S. Gama-Arachchige. 2018. Functional morpho-anatomy of water-gap complexes in physically dormant seed. Seed Science Research 28: 186–191.
Grant, C.D., and W.A. Loneragan. 1999. The effects of burning on the understorey composition of 11–13 year-old rehabilitated bauxite mines in Western Australia—vegetation characteristics. Plant Ecology 145: 291–305.
Hernández-Fernández, M., M.A. Álvarez Sierra, and P. Peláez-Campomanes. 2007. Bioclimatic analysis of rodent palaeofaunas reveals severe climatic changes in Southwestern Europe during the Plio-Pleistocene. Palaeogeography Palaeoclimatology Palaeoecology 251: 500–526.
Herranz, J.M., P. Ferrandis, and J.J. Martínez-Sánchez. 1999. Influence of heat on seed germination of nine woody Cistaceae species. International Journal of Wildland Fire 9: 173–182.
Herranz, J.M., P. Ferrandis, M.A. Copete, E.M. Duro, and A. Zalacaín. 2006. Effect of allelopathic compounds produced by Cistus ladanifer on germination of 20 Mediterranean taxa. Plant Ecology 184: 259–272.
Hnatiuk, R.J., and A.J.M. Hopkins. 1981. An ecological analysis of kwongan vegetation south of Eneabba, Western-Australia. Australian Journal of Ecology 6: 423–438.
Hudson, A.R., D.J. Ayre, and M.K.J. Ooi. 2015. Physical dormancy in a changing climate. Seed Science Research 25: 66–81.
Jaganathan, G.K. 2015. Are wildfires an adapted ecological cue breaking physical dormancy in the Mediterranean basin? Seed Science Research 25: 120–126.
Jaganathan, G.K. 2016. Influence of maternal environment in developing different levels of physical dormancy and its ecological significance. Plant Ecology 217: 71–79.
Jayasuriya, K.M.G.G., J.M. Baskin, and C.C. Baskin. 2008a. Cycling of sensitivity to physical dormancy-break in seeds of Ipomoea lacunosa (Convolvulaceae) and ecological significance. Annals of Botany 101: 341–352.
Jayasuriya, K.M.G.G., J.M. Baskin, R.L. Geneve, C.C. Baskin, and C. Chien. 2008b. Physical dormancy in seeds of the holoparasitic angiosperm Cuscuta australis (Convolvulaceae, Cuscuteae): Dormancy-breaking requirements, anatomy of the water gap and sensitivity cycling. Annals of Botany 102: 39–48.
Jayasuriya, K.M.G.G., J.M. Baskin, and C.C. Baskin. 2009. Sensitivity cycling and its ecological role in seeds with physical dormancy. Seed Science Research 19: 3–13.
Keeley, J.E., W.J. Bond, R.A. Bradstock, J.G. Pausas, and P.W. Rundel. 2011. Fire in Mediterranean ecosystems: Ecology, evolution and management. Cambridge: Cambridge University Press.
Keeley, J.E., and C.J. Fotheringham. 1998. Smoke-induced seed germination in California chaparral. Ecology 79: 2320–2336.
Keeley, J.E., J.G. Pausas, P.W. Rundel, and R. Bradstock. 2011b. Fire as an evolutionary pressure shaping plant traits. Trends in Plant Science 16: 406–411.
Lamont, B.B., G.E. Burrows, and D. Korczynskyj. 2022. High summer temperatures do not interact with fire to promote germination among seeds of Cistaceae: A reinterpretation of Luna (2020) with extra data on wet/dry conditions. Plant Ecology 223: 144–149.
Levin, N., and H. Saaroni. 1999. Fire weather in Israel — synoptic climatological analysis. GeoJournal 47: 523–538.
Luna, B. 2020. Fire and summer temperatures work together breaking physical seed dormancy. Scientific Reports 10: 1–10.
Luna, B., D. Chamorro, and B. Pérez. 2019. Effect of heat on seed germination and viability in species of Cistaceae. Plant Ecology and Diversity 12: 151–158.
Luna, B., and D. Chamorro. 2016. Germination sensitivity to water stress of eight Cistaceae species from the western Mediterranean. Seed Science Research 26: 101–110.
Luna, B., P. Piñas-Bonilla, G. Zavala, and B. Pérez. 2022. Effects of light and temperature on seed germination of eight Cistus species. Seed Science Research 32: 86–93.
Luna, B., B. Pérez, I. Torres, and J.M. Moreno. 2012. Effects of incubation temperature on seed germination of Mediterranean plants with different geographical distribution ranges. Folia Geobotanica 47: 17–27.
Mackenzie, B.D.E., T.D. Auld, D.A. Keith, F.K.C. Hui, and M.K. Ooi. 2016. The effect of seasonal ambient temperatures on fire-stimulated germination of species with physiological dormancy: A case study using Boronia (Rutaceae). Plos One. 11 (5): e0156142.
Manela, N., H. Shemesh, Y. Osem, Y. Carmel, C. Soref, A. Tsafrir, and O. Ovadia. 2022. Seasonal fires shape the germinable soil seed bank community in eastern Mediterranean woodlands. Journal of Plant Ecology 15: 13–25.
Martínez-Baniela, M., L. Carlón, T.E. Díaz, A. Bueno, and E. Fernández-Pascual. 2016. Plant-derived smoke and temperature effects on seed germination of five Helianthemum (Cistaceae). Flora 223: 56–61.
Mendes, P., C. Meireles, C. Vila-Vicosa, C. Musarella, and C. Pinto-Gomes. 2015. Best management practices to face degraded territories occupied by Cistus ladanifer shrublands - Portugal case study. Plant Biosystems 149: 494–502.
Midgley, J.J. 1989. Season of burn of serotinous fynbos Proteaceae: A critical review and further data. South African Journal of Botany 55: 65–170.
Moore, R.P. 1985. Handbook on tetrazolium testing. Zurich: International Seed Testing Association.
Moreira, B., and J.G. Pausas. 2012. Tanned or burned: the role of fire in shaping physical seed dormancy. Plos One. 7 (12): e51523.
Moreira, B., and J.G. Pausas. 2018. Shedding light through the smoke on the germination of Mediterranean Basin flora. South African Journal of Botany 115: 244–250.
Moreno, J.M., E. Zuazua, B. Pérez, B. Luna, A. Velasco, and V.R. de Dios. 2011. Rainfall patterns after fire differentially affect the recruitment of three Mediterranean shrubs. Biogeosciences 8: 3721–3732.
Neary, D.G., C.C. Klopatek, L.F. DeBano, and P.F. Ffolliott. 1999. Fire effects on belowground sustainability: A review and synthesis. Forest Ecology and Management 122: 51–71.
Ooi, M.K.J., T.D. Auld, and R.J. Whelan. 2004. Delayed post-fire seedling emergence linked to season: A case study with Leucopogon species (Epacridaceae). Plant Ecology 174: 183–196.
Ooi, M.K.J., T.D. Auld, and A.J. Denham. 2009. Climate change and bet-hedging: Interactions between increased soil temperatures and seed bank persistence. Global Change Biology 15: 2375–2386.
Ooi, M.K.J., T.D. Auld, and A.J. Denham. 2012. Projected soil temperature increase and seed dormancy response along an altitudinal gradient: Implications for seed bank persistence under climate change. Plant and Soil 353: 289–303.
Ooi, M.K.J., A.J. Denham, V.M. Santana, and T.D. Auld. 2014. Temperature thresholds of physically dormant seeds and plant functional response to fire: Variation among species and relative impact of climate change. Ecology and Evolution 4: 656–671.
Papanastasis, V.P. 1980. Effects of season and frequency of burning on a phryganic rangeland in Greece. Journal of Range Management 33: 251–255.
Parker, V.T., and V.R. Kelly. 1989. Seed bank in California chaparral and other Mediterranean climate shrublands. In Ecology of Soil Seed Bank, 231–56. San Diego: Academic Press.
Reyes, O., and L. Trabaud. 2009. Germination behaviour of 14 Mediterranean species in relation to fire factors: Smoke and heat. Plant Ecology 202: 113–121.
Rivas-Martínez, S. 1979. Brezales y jarales de Europa occidental (revisión fitosociológica de las clases Calluno-Ulicetea y Cisto-Lavanduletea). Lazaroa 1: 5–127.
Rodrigues-Junior, A.G., C.C. Baskin, J.M. Baskin, and Q.S. Garcia. 2018. Sensitivity cycling in physically dormant seeds of the Neotropical tree Senna multijuga (Fabaceae). Plant Biology 20: 698–706.
Rolston, M.P. 1978. Water impermeable seed dormancy. Botanical Review 44: 365–396.
Rundel, P.W., M.T.K. Arroyo, R.M. Cowling, J.E. Keeley, B.B. Lamont, J.G. Pausas, and P. Vargas. 2018. Fire and plant diversification in Mediterranean-climate regions. Frontiers in Plant Science 9: 851.
Santana, V.M., J.G. Alday, I. Adamo, J.A. Alloza, and M.J. Baeza. 2020. Climate, and not fire, drives the phylogenetic clustering of species with hard-coated seeds in Mediterranean Basin communities. Perspectives in Plant Ecology Evolution and Systematics 45: 125545.
Santana, V.M., R.A. Bradstock, M.K.J. Ooi, A.J. Denham, T.D. Auld, and M.J. Baeza. 2010. Effects of soil temperature regimes after fire on seed dormancy and germination in six Australian Fabaceae species. Australian Journal of Botany 58: 539–545.
Santana, V.M., M.J. Baeza, and M.C. Blanes. 2012. Clarifying the role of fire heat and daily temperature fluctuations as germination cues for Mediterranean Basin obligate seeders. Annals of Botany 111: 127–134.
Saura-Mas, S., S. Paula, J.G. Pausas, and F. Lloret. 2010. Fuel loading and flammability in the Mediterranean Basin woody species with different post-fire regenerative strategies. International Journal of Wildland Fire 19: 783–794.
Sosa, T., C. Valares, J.C. Alías, and N.C. Lobón. 2010. Persistence of flavonoids in Cistus ladanifer soils. Plant and Soil 337: 51–63.
Suc, J.P. 1984. Origin and evolution of the Mediterranean vegetation and climate in Europe. Nature 307: 429–432.
Tárrega, R., E. Luis-Calabuig, and L. Valbuena. 2001. Eleven years of recovery dynamic after experimental burning and cutting in two Cistus communities. Acta Oecologica 22: 277–283.
Taylor, G.B. 1981. Effect of constant temperature treatments followed by fluctuating temperatures on the softening of hard seeds of Trifolium subterraneum L. Australian Journal of Plant Physiology 8: 547–558.
Taylor, G.B. 2005. Hardseededness in Mediterranean annual pasture legumes in Australia: A review. Australian Journal of Agricultural Research 56: 645–661.
Thanos, C.A., and K. Georghiou. 1988. Ecophysiology of fire-stimulated seed germination in Cistus incanus ssp. creticus (L.) Heywood and C. salvifolius L. Plant, Cell and Environment 11: 841–849.
Thanos, C.A., K. Georghiou, C. Kadis, and C. Pantazi. 1992. Cistaceae: A plant family with hard seeds. Israel Journal of Botany 41: 251–263.
Tieu, A., K.W. Dixon, K.A. Meney, and K. Sivasithamparam. 2001. Interaction of soil burial and smoke on germination patterns in seeds of selected Australian native plants. Seed Science Research 11: 69–76.
Traba, J., F.M. Azcárate, and B. Peco. 2006. The fate of seeds in Mediterranean soil seed banks in relation to their traits. Journal of Vegetation Science 17: 5–10.
Trabaud, L. 1987. Fire and survival traits of plants. In Role of Fire in Ecological Systems, 65–89. The Hague: SPB Academic Publishing.
Trabaud, L. 1995. Modalités de germination des cistes et des pins méditerranéens et colonisation des sites perturbées. Revue D’écologie-La Terre Et La Terre Vie 50: 3–14.
Tsafrir, A., Y. Osem, H. Shemesh, Y. Carmel, C. Soref, and O. Ovadia. 2018. Fire season modifies the perennial plant community composition through a differential effect on obligate seeders in eastern Mediterranean woodlands. Applied Vegetation Science 22: 115–126.
Van Assche, J.A., K.L.A. Debucquoy, and W.A.F. Rommens. 2003. Seasonal cycles in the germination capacity of buried seeds of some Leguminosae (Fabaceae). New Phytologist 158: 315–323.
Vázquez, A., and J.M. Moreno. 1998. Patterns of lightning-, and people-caused fires in peninsular Spain. International Journal of Wildland Fire 82: 103–115.
Vecín-Arias, D., F. Castedo-Dorado, C. Ordóñez, and J.R. Rodríguez-Pérez. 2016. Biophysical and lightning characteristics drive lightning-induced fire occurrence in the central plateau of the Iberian Peninsula. Agricultural and Forest Meteorology 225: 36–47.
Zomer, M., B. Moreira, and J.G. Pausas. 2022. Fire and summer temperatures interact to shape seed dormancy thresholds. Annals of Botany 129: 809–816.
Funding
Funding was provided by Junta de Comunidades de Castilla-La Mancha, the European Regional Development Fund (ERDF), and the University of Castilla-La Mancha. This study was supported by the PRESFIRE project (SBPLY/19/180501/000130).
Author information
Authors and Affiliations
Contributions
BL conceived the study idea, and all authors designed the study. BL and PP carried out the experiments. BL was the largest contributor to writing the manuscript. All authors read the manuscript, contributed to the writing, and approved it.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: Table S1.
Coordinates and altitude for locations where seeds were harvested. Table S2. Results from GLM for main effects of fire and summer temperature treatments and their interactions on T0. Table S3. Results from GLM for main effects of fire and summer temperature treatments and their interactions on T50. Fig. S1. T0 for the studied Cistus species and treatments. Fig. S2. T0 for the studied Halimium species and treatments. Fig. S3. T50 for the studied Cistus species and treatments. Fig. S4. T50 for the studied Halimium species and treatments. Fig. S5. Seed viability percentages of Cistus species for each treatment. Fig. S6. Seed viability percentages of Halimium species for each treatment.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Luna, B., Piñas-Bonilla, P., Zavala, G. et al. Timing of fire during summer determines seed germination in Mediterranean Cistaceae. fire ecol 19, 52 (2023). https://doi.org/10.1186/s42408-023-00210-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s42408-023-00210-6