
Abstract
Background
Shortage of water is a major problem facing Egypt; thus, it becomes necessary to use non-traditional sources of water such as saline water in irrigation. Overcoming the adverse effects of saline water and enhancing plant tolerance to salinity stress is the main challenge for increasing plant growth and productivity. Using a natural compound such as cysteine amino acid has an important effect in alleviating the adverse effect of salinity stress on different plant crops.
Materials and methods
Two pot experiments were carried out during two successive summer seasons to study the beneficial role of cysteine (0, 20, and 40 mg/l) in enhancing growth, some metabolic process, and seed yield quality and quantity of soybean plant grown under salinity stress (0, 3000, and 6000 mg/l).
Results
Salinity stress (3000 and 6000 mg/l) caused decreases in soybean growth criteria (plant height, number of branches and leaves/plant, dry weight of leaves and stem/plant), photosynthetic pigments and some element contents (nitrogen (N), phosphorus (P), and potassium (K)), and seed yield and yield components (number of pods/plant, weight of pods/plant and number of seeds/plant) as well as oil%. Meanwhile, proline, H2O2 and MDA contents, and superoxide dismutase activity were gradually increased by increasing salinity level. On the other hand, cysteine treatments improved growth and yield of soybean plant either irrigated with tap water or saline water. Cysteine treatments could alleviate the adverse effect of salinity stress on growth and yield of soybean plant through increasing photosynthetic pigments; proline content; N, P, and K contents; superoxide dismutase and catalase activities; and oil% accompanied by decreases in H2O2 and MDA contents as compared with their corresponding controls. In addition, cysteine treatments and/or salinity stress exhibited differences in protein pattern from 112 to 19 kD molecular weight. The appearance of new protein bands reflected the expression of cysteine treatments and salinity stress.
Conclusion
Cysteine treatments had a beneficial role in alleviating the adverse effect of salinity stress on soybean plant. Forty milligrams per liter of cysteine was the most effective treatment in enhancing salinity tolerance of soybean plant.
Similar content being viewed by others


Introduction
All over the world, soybean (Glycine max L. Merril) is one of the most important and cheap protein sources and vegetable oil seed crops. Thus, in many developing countries, soybean is used as an important component of human diets and animal feed. Soybean seeds contain about 20% edible oil, 35% protein, and 35% carbohydrates (17% of which is dietary fiber), and about 5% ash as well as vitamins and minerals (Thomas and Erostus 2008; Dixit et al. 2011). Soybean oil plays an important role in nutrition of human as it has high quantity of essential unsaturated fatty acids (omega-3, omega-6, and omega-9) (Nikolic et al. 2009). Likewise, soybean seeds are considered as a good source of protein due to the presence of different essential amino acids. Moreover, soybean helps in improving fertility of soil through fixing atmospheric nitrogen by Rhizobium bacteria which is present in the root nodules (Bakhoum et al. 2019).
Salinity stress is one of the serious abiotic stresses which adversely affect plant growth and productivity via changes in different metabolic and physiological processes and depending on the duration and severity of the stress. Salinity affect plant growth through osmotic stress followed by ion toxicity (James et al. 2011). Osmotic stress caused disorders in different physiological processes, such as nutrient imbalance, membrane interruption, and impaired ability in detoxifying reactive oxygen species (ROS) which affect antioxidant enzyme activity and decline photosynthetic activity and stomatal aperture (Munns and Tester 2008). The production of ROS as singlet oxygen, superoxide, hydroxyl radical, and hydrogen peroxide is increased in response to salinity stress. The increases in ROS production caused oxidative damages to different cellular components such as proteins, lipids, and RNA (Apel and Hirt 2004). Plant develops different biochemical and physiological mechanisms to survive under salinity stress. The main mechanisms are compartmentalization and ion homeostasis, uptake and transport of ions, osmoprotectants and compatible biosynthesis, synthesis of antioxidant compounds and antioxidant enzyme activation, etc. (Gupta and Huang 2014).
To overcome the deleterious effects of salinity stress and enhance plant tolerance to salt stress, certain compounds which are present naturally in plants and have positive effects on plant growth and yield may be used. One of these compounds is amino acids that have the ability to scavenge ROS which is produced during photosynthesis and respiration processes (Foyer et al. 1991). Amino acids are important bio-regulators which have promotive effects on plant growth and productivity. Moreover, amino acids could act as a part of co-enzymes and precursors of certain plant bio-regulators that promote plant growth through enhancing photosynthesis (Bakhoum et al. 2019), synthesis of carbohydrates and protein (Maxwell and Kieber 2004), as well as spermine and spermidine (Singh 1999). Cysteine is an α-amino acid with a thiol side chain which participates in enzymatic reactions. Cysteine plays a structural main function in proteins as well as its role as a precursor for essential biomolecules, such as glutathione (antioxidant) and vitamins and some defense compounds, such as glucosinolates and thionins (Alvarez et al. 2012). Cysteine is the precursor molecule of glutathione, the predominant non-protein thiol, which plays an important role in plant stress responses (Azarakhsh et al. 2015).
Thus, the present study aimed to investigate the physiological role of cysteine treatments in alleviating the deleterious effect of salinity stress on growth, some biochemical aspects, and yield quantity and quality of soybean plant.
Materials and methods
Experimental procedure
Soybean seeds of Giza 111 cultivar were obtained from the Agricultural Research Center, Giza, Egypt. Two pot experiments were conducted at the greenhouse of National Research Centre, Dokki, Giza, Egypt, during two successive summer seasons.
Soybean seeds of equal size and the same color were selected and washed with distilled water, sterilized with 1% sodium hypochlorite solution for about 2 min and washed again with distilled water. Clay soil is mixed with yellow sand in a proportion of 3:1 in order to improve drainage. N and P fertilizers were mixed into the soil before sowing. Ten soybean seeds were sown along a center row in each plastic pots of about 7 kg clay soils. Ten days after sowing, the seedlings were thinned to three seedlings per pot. The experimental design was a factorial experiment in a complete randomized block design with six replicates. Soybean plants were exposed to foliar spraying with cysteine (0, 20, and 40 mg/l) at 30 and 45 days from sowing. Three saline water levels (0, 3000, and 6000 mg/l) were prepared by dissolving sea salt with tap water. The plants were regularly irrigated with saline water after 30 days from sowing.
Data recorded
Plant samples were taken after 60 days from sowing for measurements of growth criteria as plant height (cm), number of branches and leaves/plant, and dry weight of leaves and stem/plant (g). Plant samples were taken for biochemical analysis as photosynthetic pigments, proline, hydrogen peroxide, lipid peroxidation contents and activity of two antioxidant enzymes (superoxide dismutase and catalase), mineral content (N, P, K), as well as protein pattern. At harvest, the following characters were recorded: number of pods/plant, weight of pods/plant (g), seed number and weight of seeds/plant (g), and oil%.
Biochemical analysis
Photosynthetic pigments (chlorophyll a and b and carotenoid) in fresh leaf tissues were estimated according to the method of Lichtenthaler and Buschmann (2001). Proline content was determined according to Bates et al. (1973). Hydrogen peroxide content was determined using the method of Velikova et al. (2000) in fresh leaf tissues. The level of lipid peroxidation was measured by determining the levels of malondialdehyde (MDA) content using the method of Hodges et al. (1999). Enzyme extracts were collected following the method described by Chen and Wang (2006). Fresh leaf tissues were homogenized in ice-cold phosphate buffer (50 mM, pH 7.8), followed by centrifugation at 8000 rpm and 4°C for 15 min. The supernatant was used immediately to determine the activities of enzymes. Superoxide dismutase activity (SOD) (EC 1.12.1.1) activity was spectrophotometrically assayed at 560 nm by nitro blue tetrazolium (NBT) reduction method (Chen and Wang 2006). Catalase activity (CAT) (EC 1.11.1.6) activity was determined spectrophotometrically by following the decrease in absorbance at 240 nm (Chen and Wang 2006). The enzyme activities were calculated by Kong et al. (1999). Protein pattern (SDS-PAGE) was studied using sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) as described by Laemmli (1970). Data matrix was prepared taking 0 and 1 value for the absence and presence of a particular protein band. Protein pattern (SDS-PAGE) was studied using SDS-PAGE as described by (Laemmli 1970). Data matrix was prepared taking 0 and 1 values for the absence and presence of a particular protein band. Phosphorus (P), potassium (K), and nitrogen (N) were measured in dry leaf tissues. The plant material was digested using an acid mixture consisting of nitric, perchloric, and sulfuric acids in the ratio of 8:1:1 (v/v). Phosphorus and potassium were estimated photometrically using the flame photometer method described by the methods of Chapman and Pratt (1978). Total nitrogen (N) was determined using the micro-Kjeldahl method, as described by AOAC (Association of official agriculture chemists) (2000). The oil content of soybean seeds were extracted and determined according to Horwitz and Latimer (2005).
Statistical analysis
The data obtained were statistically analyzed using MSTAT-C statistical software. The differences between treatment means were compared by Duncan’s multiple range test (DMRT) at 5% probability level according to Steel et al. (1997).
Results
Changes in growth criteria
Irrigation of soybean plant with saline water (3000 and 6000 mg/l) caused non-significant decreases in different growth criteria (plant height, number of branches and leaves/plant, dry weight of leaves/plant) except for dry weight of stem that showed non-significant increases at salinity stress (3000 mg/l) compared with control plant (Table 1). On the other hand, cysteine treatments at 20 and 40 mg/l could improve and alleviate salinity tolerance of the plant by increasing the abovementioned growth criteria as compared with corresponding controls. These increases in different growth criteria are not only in plants irrigated with saline water but also in those irrigated with tap water (Table 1).
Changes in photosynthetic pigments
Effect of different salinity levels on photosynthetic pigments of soybean is shown in Table 2. Increasing salinity levels from 3000 to 6000 mg/l caused gradual decreases in all photosynthetic pigments (chlorophyll a, chlorophyll b, and carotenoid) of soybean leaves compared with control plants (plants irrigated with tap water). On the other hand, cysteine treatments at 20 and 40 mg/l could alleviate the harmful effect of salinity via increases of photosynthetic pigment components as compared with their corresponding controls. The highest increases of the photosynthetic pigments were achieved by application of cysteine treatment at 40 mg/l.
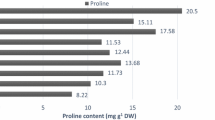
Changes in proline content
The data in Table 2 obviously indicate that increasing salinity levels caused significant increases in soybean leaf proline content, as compared with control plants (plants irrigated with tap water). In addition, cysteine treatments (20 and 40 mg/l) induced proline content in stressed plants as well as in unstressed plants relative to their corresponding controls. Forty milligrams per liter of cysteine was the most effective treatment in increasing proline content of soybean plant.
Changes in H2O2 content
Salinity stress significantly increased the hydrogen peroxide (H2O2) content in soybean leaves (Table 3). The H2O2 level is increased by 43.7% in plants irrigated with low saline water (3000 mg/l) and 71.3% in plants irrigated with higher level of saline water (6000 mg/l) as compared with control plants irrigated with tap water. On the other hand, cysteine treatment significantly decreased H2O2 content in the normally irrigated plants and salinity stressed plants compared with their corresponding control. The percentages of decreases were 20.4, 40.4, and 37.8% in response to 40 mg/l cysteine treatment as compared with their corresponding controls in each salinity level (0, 3000, and 6000 mg/l) respectively.
Changes in lipid peroxidation
Salinity stress induced oxidative damage of cell membrane assessed by increasing in lipid peroxidation in soybean plant as compared with unstressed control plants (Table 3). The level of malondialdehyde (MDA) was increased by 42.0% in plants irrigated with low saline water (3000 mg/l) and 78.9% in plants irrigated with higher level of saline water (6000 mg/l) as compared with control plants irrigated with tap water. Meanwhile, gradual and significant decreases in lipid peroxidation by cysteine treatments (20 and 40 mg/l) occurred in unstressed plants and salinity stressed plants as compared with their corresponding controls. The percentages of decreases were 20.8, 33.0, and 28.8% in response to 40 mg/l cysteine treatment as compared with their corresponding controls in each salinity level (0, 3000, and 6000 mg/l) respectively.
Changes in antioxidant enzyme activities
The changes in antioxidant enzyme activities such as superoxide dismutase (SOD) and catalase (CAT) in soybean plants grown under salinity stress are presented in Table 3. Salinity stress caused gradual and significant increases in SOD activity as compared with control plants. CAT activity was significantly decreased due to salinity stress. On the other hand, cysteine treatments at 20 and 40 mg/l caused significant increases in SOD and CAT activity either in plants irrigated with tap water or those irrigated with saline water as compared with corresponding controls.
Changes in nitrogen, phosphorus, and potassium content
Table 4 shows the effect of cysteine treatments and/or salinity stress on N, P, and K contents of soybean plants. The data clearly show that irrigation of soybean plants with different salinity levels caused significant decreases in N, P, and K contents as compared with plants irrigated with tap water. Meanwhile, cysteine treatments at 20 and 40 mg/l caused a significant increase in the content of the abovementioned elements as compared with their corresponding controls.
Protein pattern (SDS-PAGE)
The electrophoretic pattern of leaf soluble protein of soybean treated with cysteine at 20 and 40 mg/l and grown under salinity stress (0, 3000, and 6000 mg/l) was shown in Table 5 and Fig. 1. The data show the appearance of different protein bands with molecular weights from 112 to 19 kD. The variations in protein electrophoretic pattern under salinity stress and cysteine-treated plants were presented as the appearance of new polypeptide bands and disappearance of other polypeptide bands. Salinity stress at 3000 mg/l expressed new polypeptide bands of molecular weight 35 and 23 kD comparing with control plant. In contrast, bands of molecular weights 112 and 21 kD appeared under control condition but is absent under salinity stress. Treatment with 6000 mg/l salinity level showed the appearance of new protein bands at 57 and 44 kD and disappearance of three bands at 112, 90, 50, and 21 kD comparing with control plants.

Protein pattern of leaf-soluble protein of soybean grown under salinity stress and/or cysteine treatments by using SDS-PAGE. (1) control (tap water), (2) 20 mg/l cysteine + tap water, (3) 40 mg/l cysteine + tap water, (4) 3000 mg/l sea salt, (5) 20 mg/l cysteine + 3000 mg/l sea salt, (6) 40 mg/l cysteine + 3000 mg/l sea salt, (7) 6000 mg/l sea salt, (8) 20 mg/l cysteine + 6000 mg/l sea salt, (9) 40 mg/l cysteine + 6000 mg/l sea salt
Soybean plants grown under 3000 mg/l of salinity and treated with cysteine at 40 mg/l showed the appearance of a new band with a molecular weight of 84 kD. While soybean plant is treated with cysteine at 40 mg/l and grown under salinity stress at 6000 mg/l, it showed a new band of 64 kD. It was observed that cysteine treatment at 40 mg/l and salinity stress at 3000 and 6000 mg/l showed a new polypeptide band at 102 kD. On the other hand, our results showed there was a new band of 26 kD due to exogenous application of cysteine at 20 mg/l under unstressed and stressed conditions. A new band of 19 kD appeared in soybean plant treated with cysteine at 20 mg/l and irrigated with tap water.
Changes in seed yield and yield components
Data show that salinity stress caused significant decreases in seed yield (Table 6). A significant decrease in yield components (pods’ weight and seeds’ number/plant) appeared due to irrigation with 6000 mg/l saline water as compared with those plants irrigated with tap water (Table 6). Salinity stress at 6000 mg/l caused the highest significant decreases in seed yield and yield components and oil content of soybean. On the other hand, treated soybean plant with cysteine at 20 and 40 mg/l could alleviate the deleterious effect of salinity stress via increases that occurred in seed yield and yield components as well as oil content when compared with plants irrigated with saline water.
Discussion
Changes in growth and yield of soybean plant
The adverse effects of salinity stress on plant growth and yield may be attributed to low osmotic potential of soil solution, imbalance of various ions and ion toxicity (Pasternak 1987), reduction in carbon fixation (Niu et al. 1995), partial stomata closure that caused reduction of photosynthesis (Pasternak 1987), and nutritional imbalances which caused limitation of growth (Grattan and Grieve 1999). Moreover, salinity stress caused some changes in physiological, biochemical, and molecular processes which adversely reduced growth and yield of plant (Ashraf and Harris 2004). Since, the decreases in growth of different crops could be referred to the destructive role of salinity stress on different biochemical processes as antioxidant enzyme activity, photosynthesis, and ion homeostasis. The reduced effects of salinity on plant growth and yield of soybean plant (Tables 1 and 6) are congruent with those reported by Dawood et al. (2016) on lupine plant and El-Awadi et al. (2017) on faba bean plant. On the other hand, cysteine treatments increased soybean growth and yield (Tables 1 and 6). The enhancement effect of cysteine amino acid on growth criteria of soybean plant might be attributed to GSH synthesis from cysteine (Nasibi et al. 2016). In general, amino acids application have a high integrity with various metabolic processes in cells that promote growth and yield of different plants (Coruzzi and Last 2000). The promotive effect of various amino acids on growth and yield of different plant species was detected by Nasibi et al. (2016) on wheat plant and El-Awadi et al. (2016) on faba bean plant.
Changes in photosynthetic pigments
Total chlorophyll (chlorophyll a + chlorophyll b) is an important symptom correlated with photosynthetic efficiency (Kirschbaum 2011). Chlorophyll a, chlorophyll b, and carotenoid contents were decreased gradually with increasing salinity levels (Table 2). These decreases might be referred to the increased production of reactive oxygen species (ROS) and oxidative stress that damaged chloroplast membranes and/or due to the antagonistic effects of sodium ion on magnesium ion absorption (Smirnoff 1993). In addition, these decreases could be considered as an important regulatory step to avoid the absorbance of high light and thus reduced the production of ROS (Munné-Bosch and Alegre 2000; Miller et al. 2010). Salinity stress caused osmotic stress which adversely affect photosynthetic pigments. Osmotic stress induced water deficit and consequently closed stomata, which decreased CO2 availability and reduced photosynthesis leading to excess production of ROS in chloroplasts (Parida et al. 2004). These reductions in different photosynthetic pigment components in response to salt stress were confirmed by Sadak et al. (2015) and Ismail et al. (2019) on faba bean plant. Cysteine treatments caused marked increases of all photosynthetic pigment components as compared with the corresponding controls (Table 2). This promotive effect of cysteine in increasing photosynthetic pigments of soybean might be due to the fact that the chloroplast is the main source of sulfide via sulfate reduction in the sulfur assimilation pathway (Takahashi et al. 2011). The increased contents of carotenoids play an important role of plant defense against salinity stress as it is well recorded that carotenoids have vital importance as they act as photoreceptors pigments (Lawlor 1989) and as modulator for eliminating singlet oxygen (Fyfe et al. 1995). Cysteine treatments increased the photosynthetic pigment of different species such as faba bean (El-Awadi et al. 2016), wheat plant (Nasibi et al. 2016), and Egyptian clover using tryptophan (El Karamany et al. 2018) and soybean cultivars using methionine (Bakhoum et al. 2019).
Changes in proline content
Salinity stress and/or cysteine treatments increased proline content of soybean (Table 2). Proline accumulates in large amounts in salt-stressed plants (Yamada et al. 2005; Abd El-Samad et al. 2011). Its accumulation in plants could be due to de novo synthesis or decreased degradation or both and depends upon the type of species and extent of stress (Kavi-Kishor et al. 2005). The accumulation of the proline in plant tissues in response to different abiotic stresses may play an important role against oxidative damages (Alia et al. 2001), take part in cellular osmotic adjustment (Yamada et al. 2005), and stabilize the membrane and protein 3D structure (Kavi-Kishor et al. 2005; Ashraf and Foolad 2007).
Changes in H2O2 and MDA contents
H2O2 and MDA contents were increased in soybean plant subjected to different salinity levels (Table 3). H2O2 is used in different biochemical processes and signaling cascades required for plant growth and development. Generally, H2O2 contents increased in various plants subjected to stress (Dawood and Sadak 2014; Sadak 2016; Kaur et al. 2017). Under osmotic stress, plants produce ROS such as H2O2, which are harmful to plant growth through their effects on subcellular components and plant metabolism causing oxidative destruction of cells (Gunes et al. 2007; Maksup et al. 2014). The increased levels of ROS caused deterioration of lipids of membranes and thus increased MDA content and leakage of solutes from membranes (Gunes et al. 2007). The increases in MDA level caused lipid membrane peroxidation and were used as an indicator of oxidative damage (Jain et al. 2001). The level of MDA is considered as a biomarker of stress and increased as severity of stress increased. Guimarães et al. (2011) showed that increased MDA content might result in electrolyte leakage, indicating a loss of membrane integrity. These obtained results are similar to those obtained by Sadak et al. (2017) on chickpea plant and by El-Awadi et al. (2017) on sunflower plant.
On the other hand, cysteine treatments decreased H2O2 and MDA content in soybean plant subjected to different salinity levels (Table 3). The beneficial role of cysteine in decreasing H2O2 and MDA contents might be attributed to its product, such as glutathione, which has antioxidant activity. It has been reported that exogenous glutathione increased phenol peroxidase activity with the reduction of deleterious effect of salinity in Brassica napus L. as well as it directly scavenge H2O2 and maintain the membrane integrity (Sadak et al. 2017). Nasibi et al. (2016) reported that cysteine treatment ameliorate the reduced effect of salinity stress of wheat plant by decreasing H2O2 and MDA contents.
Changes in antioxidant enzyme activities
Salinity stress significantly increased SOD activity and significantly decreased CAT activity as compared with control plants (Table 4). Oxidative stress is an important sign of abiotic stress and increased SOD activity that correlated with increasing protection from destructive effect of stress (Miller et al. 2010). SOD is one of the ubiquitous enzymes in aerobic organisms’ essential for cellular defense mechanisms against ROS. SOD modulates relative amounts of superoxide anion (O2−) and the two Haber–Weiss reaction substrates and decreases the risk of –OH radical formation, which is highly reactive and may cause severe damage to membranes and proteins (Bowler et al. 1992). Data showed that salinity stress caused reduction in the activity of CAT enzymes. The reduction in catalase activity indicated that this was unable to completely neutralize H2O2 resulting from the oxidative salt stress (Shalata and Neumann 2001). The CAT had the capacity for the decomposition of H2O2 generated by SOD. Thus, CAT activity coordinated with SOD activity can represent a central protective role in O2− and H2O2 scavenging process. In addition, CAT is the main scavenger of strong oxidant H2O2 in peroxisomes and it converts H2O2 to water and molecular oxygen (Bahari et al. 2013). These results of salinity stress are confirmed earlier on different plant species (Ibrahim et al. 2016; Kaur et al. 2017). The beneficial role of cysteine in increasing antioxidant enzymes activities might be attributed to its product, such as glutathione, which has antioxidant activity. It has been reported that exogenous glutathione mitigates the deleterious effect of salinity in chickpea and it directly scavenges H2O2 and maintains the membrane integrity (Sadak et al. 2017). Nasibi et al. (2016) reported that cysteine treatment ameliorate the reduced effect of salinity stress of wheat plant by decreasing H2O2 and MDA contents via the action of different antioxidant enzymes (SOD and CAT). Generally, the increases of CAT activity are a strategy for improving the salt tolerance (Vaidyanathan et al. 2003).
Changes in element contents
Salinity stress significantly reduced the contents of soybean plant (N, P, and K+) gradually as salinity stress increased (Table 5). Salt stress disturbs intracellular ion homeostasis of plants, which leads to membrane dysfunction perturbations of metabolic activity and causes growth inhibition (Very et al. 1998). Similar observations were obtained by Perez-Alfocea et al. (1993) and Bolarin et al. (1995) who reported that the salt-tolerant relative L. pennelli plant was associated with both a high capacity for osmotic adjustment and a greater ability to take up ions and translocate them to leaves. Cysteine treatments caused significant increases in N, P, and K content in soybean plants irrigated with tap water or saline water. Nasibi et al. (2016) and El-Awadi et al. (2016) showed that cysteine treatment resulted in a higher K content of wheat plant and N, P, and K of faba bean.
Protein pattern (SDS-PAGE)
The variations in protein electrophoretic pattern under salinity stress and/or cysteine treatments were presented as the appearance of new polypeptide bands and disappearance of other polypeptide bands. Salinity stress synthesized new proteins knows as stress-specific proteins (Table 5). The expression of protein pattern reflected the plant tolerant to salt stress. Bavei et al. (2011) reported that a specific polypeptide was synthesized as a salt adaptation for plants under salt stress. Other studies revealed that there were specifically proteins synthesized under stress conditions in the rice leaf (Parker et al. 2006) and in faba bean seedlings (Younis et al. 2009). Hamdia et al. (2016) revealed that three bands were induced under salinity stress as compared to untreated plants. Numerous studies have shown that the changes in protein pattern by electrophoretic analysis under abiotic stresses such as heat treatment and a new protein band appeared in treated samples (Polenta et al. 2007). Salinity stress might be identified proteins related to salt stress as comparing with control conditions (Sobhanian et al. 2016). Likewise, cysteine treatment at 20 mg/l showed a new band of 19 kD. On the other hand, exogenous application of cysteine at 20 mg/l showed a new band of 26 kD under unstressed condition or salinity stress. Finally, both of salinity stress and/or cysteine treatment induced changes of the protein patterns with various intensity. The new protein pattern exhibited under salinity stress and cysteine treatments reflected the expression of salinity tolerance of soybean.
Conclusion
This study indicated that the foliar application of cysteine at the concentration of 40 mg/l was the most effective treatment in enhancing the photosynthetic pigments, the N, P, K contents, the oil%, and the activities of antioxidant enzymes (SOD and CAT) and in lowering H2O2 and MDA contents under salinity stress. Also, the appearance of new polypeptide bands exhibited under salinity stress and cysteine treatments reflected the expression of salinity tolerance of soybean. Therefore, the foliar application of 40 mg/l cysteine had a beneficial role in alleviating the adverse effect of salinity stress on soybean plant.
Availability of data and materials
The datasets generated and/or analyzed during the current study are included in this published manuscript.
Abbreviations
- CAT:
-
Catalase
- Chl:
-
Chlorophyll
- CV:
-
Cultivar
- K:
-
Potassium
- MDA:
-
Malondialdehyde
- N:
-
Nitrogen
- P:
-
Phosphorus
- ROS:
-
Reactive oxygen species
- SOD:
-
Superoxide dismutase
References
Abd El-samad HM, Shaddad MAK, Barakat N (2011) Improvement of plants salt tolerance by exogenous application of amino acids. J Med Plant Res 5:5692–5699
Alia JM, Mohanty P, Matysik J (2001) Effect of proline on the production of singlet oxygen. Amino Acids 21:195–200
Alvarez C, Bermudez A, Romero M, Gotor LC, Garcia CI (2012) Cysteine homeostasis plays an essential role in plant immunity. New Phytologist 193:165–177
AOAC (Association of official agriculture chemists) (2000) Official methods of analysis, 17th edn. Association of official agriculture chemists, Gaithersburg
Apel K, Hirt H (2004) Reactive oxygen species: metabolism, oxidative stress, and signal transduction. Ann Rev Plant Biol 55:373–399
Ashraf M, Foolad MR (2007) Roles of glycinebetaine and proline in improving plant abiotic stress tolerance. Environ Exp Bot 59:206–216
Ashraf M, Harris PG (2004) Potential biochemical indicators of salinity tolerance in plants. Plant Sci 166:3–16
Azarakhsh M, Asrar RZ, Mansouri H (2015) Effects of seed and vegetative stage cysteine treatments on oxidative stress response molecules and enzymes in Ocimum basilicum L. under cobalt stress. J Soil Sci Plant Nutr 15(3):651–662
Bahari A, Pirdashti H, Yaghubi M (2013) The effects of amino acid fertilizers spraying on photosynthetic pigments and antioxidant enzymes of wheat (Triticum aestivum L.) under salinity stress. Int J Agronomy Plant Product 4:787–793
Bakhoum GS, Badr EA, Sadak MS, Kabesh MO, Amin GA (2019) Improving growth, some biochemical aspects and yield of three cultivars of soybean plant by methionine treatment under sandy soil condition. Int J Enviro Res 13(1):35–43
Bates LS, Waldan RP, Teare LD (1973) Rapid determination of free proline for water stress studies. Plant Soil. 39:205–207
Bavei V, Shiran B, Khodambashi M, Ranjbar A (2011) Protein electrophoretic profiles and physiochemical indicators of salinity tolerance in sorghum (sorghum bicolor L.). Afric J Biotech 10(14):2683–2697
Bolarin MC, Santa-Cruz A, Cayuela E, Perez-Alfocea F (1995) Short-term solute changes in leaves and roots of cultivated and wild tomato seedlings under salinity. J Plant Physiol. 147:463
Bowler C, Montagu MV, Inze D (1992) Superoxide dismutase and salt stress tolerance. Ann Rev Plant Biol 43:83–116
Chapman HD, Pratt PF (1978) Methods of Analysis for Soils, Plant and Water. Univ California Div Agric Sci Publ 4034:162–165
Chen JX, Wang XF (2006) Plant physiology experimental guide. Higher Educ Press 24–25:55–56
Coruzzi G, Last R (2000) Amino acids. In: Buchanan B, Gruissem W, Jones R (eds) Biochemistry and molecular biology of plants. Amer Soc Plant Biol, Rockville, pp 358–410
Dawood MG, El-Metwally IM, Abdelhamid MT (2016) Physiological response of lupine and associated weeds grown at salt-affected soil to α-tocopherol and hoeing treatments. Gesunde Pflanzen 68:117–127
Dawood MG, Sadak MS (2014) Physiological role of glycinebetaine in alleviating the deleterious effects of drought stress on canola plants (Brassica napus L.). Middle East J Agric Res 3(3):943–954
Dixit AK, Antony JIX, Sharma NK, Tiwari RK (2011) Opportunity, challenge and scope of natural products in medicinal chemist, vol. Soybean constituents and their functional benefits. Res Signpost, 37/661(2) 12:367–383
El Karamany MF, Omer HAA, Bakry BA, Bakhoum GS, Sadak MS (2018) Impact of tryptophan treatment on yield and chemical composition of Berseem green fodder. Biosci Res 15(4):3692–3707
El-Awadi ME, Ibrahim SK, Sadak MS, Abd Elhamid EM, Gamal El-Din KM (2016) Impact of cysteine or proline on growth, some biochemical attributes and yield of faba bean. Int J Pharm Tech Res. 9(6):100–106
El-Awadi ME, Sadak MS, Dawood MG, Khater MA, Elashtokhy MMA (2017) Amelioration the adverse effects of salinity stress by using γ-radiation in faba bean plants. Bull. NRC 41(1):293–310
Foyer C, Lelandais M, Edwards EA, Mulineawx PMM (1991) The role of ascorbate in plants, interactions with photosynthesis and regulatory significance. In: Pell EJ, Steffen KL (eds) Active oxygen oxidative stress and plant metabolism, vol 6. Current Topics in plant physiology. American Society of Plant Physiologists, Rockville, pp 131–144
Fyfe P, Cogdell RJ, Hunter CN, Jones MR (1995) Study of the carotenoid binding pocket of photosynthetic reaction center from the purple bacterium rhodobacter sphaeroides. I: Photosynthesis from Light to Biosphere. Mathis (ed) Vol. IV, 47-50 in Proceeding of Xth international photosynthesis congress, Montpellier, France, 20-25 August.
Grattan SR, Grieve CM (1999) Salinity—mineral nutrient relations in horticultural crops. Scientia Horticulturae 78:127–157
Guimarães FVA, de Lacerda CF, Marques EC, de Miranda MRA, de Abreu CEB (2011) Calcium can moderate changes on membrane structure and lipid composition in cowpea plants under salt stress. Plant Growth Regul 65:55–63
Gunes A, Inal A, Alpaslan M, Eraslan F, Bagci EG, Cicek N (2007) Salicylic acid induced changes on some physiological parameters symptomatic for oxidative stress and mineral nutrition in maize (Zea mays L.) grown under salinity. J Plant Physiol 164:728–736
Gupta B, Huang B (2014) Mechanism of salinity tolerance in plants: physiological, biochemical, and molecular characterization. Hindawi Publishing Corporation. Int J Genomics, Article ID 701596, 18 pages doi: https://doi.org/10.1155/2014/701596
Hamdia M, Abd ES, Shaddad MAK (2016) Mechanisms of salt tolerance of wheat cultivars. Triticeae Genomics Genet 7:1–16
Hodges DM, De Long JM, Forney C, Prange PK (1999) Improving the thiobarbituric acid reactive substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta 207:604–611
Horwitz W, Latimer J (2005) Official methods of analysis of AOAC international. AOAC international, Gaithersburg
Ibrahim MFM, Faisal A, Shehata SA (2016) Calcium chloride alleviates water stress in sunflower plants through modifying some physio-biochemical parameters. American-Eurasian J Agric Environ Sci 16(4):677–693
Ismail RR, Ahmad ES, Abdel-Fattah M, Omar MN, Azzam CR, Zahran HH (2019) Effect of plant growth-promoting rhizobacteria on growth and symbiotic nitrogen fixation of vicia faba plants under salt stress. Plant Sci J 7(1):01–19
Jain M, Mathur G, Koul S, Sarin NB (2001) Ameliorative effects of proline on salt stress-induced lipid peroxidation in cell lines of groundnut (Arachis hypogaea L.). Plant Cell Rep 20:463–468
James R, Blake IC, Byrt CS, Munns R (2011) Major genes for Na+ exclusion, Nax1 and Nax2 (wheat HKT1; 4 and HKT1; 5), decrease Na+ accumulation in bread wheat leaves under saline and waterlogged conditions. J Exper Bot 62(8):2939–2947
Kaur H, Bhardwaj RD, Satvir K (2017) Grewal. Mitigation of salinity-induced oxidative damage in wheat (Triticum aestivum L.) seedlings by exogenous application of phenolic acids. Acta Physiol Plant 39:221
Kavi-Kishor PB, Sangam S, Amrutha RN, Sri Laxmi P, Naidu KR, Rao KRSS, Rao S, Reddy KJ, Theriappan P, Sreeniv N (2005) Regulation of proline biosynthesis, degradation, uptake and transport in higher plants: its implications in plant growth and abiotic stress tolerance. Curr Sci 88:424–438
Kirschbaum MUF (2011) Does Enhanced Photosynthesis Enhance Growth? Lessons Learned from CO2 Enrichment Studies. Plant Physiol 155(1):117–124
Kong FX, Hu W, Chao WL, Sang WL, Wang LS (1999) Physiological responses of Mexicana to oxidative stress of SO2. Environ Exp Bot 42:201–209
Laemmli UK (1970) Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227:680–685
Lawlor DW (1989) Photosynthesis: metabolism, control and physiology. Longman Scientific and Technical Group, London, pp 217–243
Lichtenthaler HK, Buschmann C (2001) Chlorophylls and carotenoids: measurement and characterization by UV-VIS spectroscopy. In: Wrolstad RE, Acree TE, An H, Decker EA, Penner MH, Reid DS, Schwartz SJ, Shoemaker CF, Sporns P (eds) Current protocols in food analytical chemistry (CPFA). Wiley, New York, pp F4.3.1–F4.3.8
Maksup S, Roytrakul S, Supaibulwatana K (2014) Physiological and comparative proteomic analyses of Thai jasmine rice and two check cultivars in response to drought stress. J Plant Interact 9:43–55
Maxwell B, Kieber J (2004) Cytokinin signal transduction. In: Davies PJ (ed) Plant hormones. Biosynthesis, signal transduction, action. Kluwer, Dordrecht, pp 321–349
Miller G, Suzuki N, Ciftci-yilmaz S, Mittler R (2010) Reactive oxygen species homeostasis and signaling during drought and salinity stresses. Plant Cell Environ 33:453–467
Munné-Bosch S, Alegre L (2000) The xanthophyll cycle is induced by light irrespective of water status in field-grown lavender (Lavandula stoechas) plants. Physiol Plant 108:147–151
Munns R, Tester M (2008) Mechanisms of salinity tolerance. Ann Rev Plant Biol 59:651–681
Nasibi F, Kalantari KM, Zanganeh R, Mohammadinejad G, Oloumi H (2016) Seed priming with cysteine modulates the growth and metabolic activity of wheat plants under salinity and osmotic stresses at early stages of growth. Ind J Plant Physiol 21(3):279–286
Nikolic NC, Cakic SM, Novacovic SM, Cvetkovic MD, Stankovic MZ (2009) Effect of extraction techniques on yield and composition of soybean oil. Maced J Chem Chem Eng 28(2):173–179
Niu X, Bressan RA, Hasegawa PM, Pardo JP (1995) Ion homeostasis in NaCl stress environments. Plant Physiol 109:735–742
Parida AK, Das AB, Mohanty P (2004) Investigations on the antioxidative defence responses to NaCl stress in a mangrove, Bruguiera parviflora: differential regulations of isoforms of some antioxidative enzymes. Plant Growth Regul 42:213–226
Parker R, Flowers TJ, Moore AL, Harpham NVJ (2006) An accurate and reproducible method for proteome profiling of the effects of salt stress in the rice leaf lamina. J Exp Bot 57:1109–1118
Pasternak D (1987) Salt tolerance and crop production—a comprehensive approach. Ann Rev Phytopathol 25:271–291
Perez-Alfocea F, Estan MT, Caro M, Guerrier G (1993) Osmotic adjustement in Lycopersicon esculentum and L. Pennellii under NaCl and polyethylene glycol 6000 iso-osmotic stresses. Physiol Plant 87:493–498
Polenta GA, Calvete JJ, Gonza’lez CB (2007) Isolation and characterization of the main small heat shock proteins induced in tomato pericarp by thermal treatment. FEBS J 274:6447–6455
Sadak MS (2016) Physiological role of signal molecules in improving plant tolerance under abiotic stress. Inter J Chem Tech Res 9(7):46–60
Sadak MS, Abd Elhamid EM, Ahmed MMRM (2017) Glutathione Induced Antioxidant Protection Against Salinity Stress in Chickpea (Cicer arietinum L.) Plant. Egypt J Bot 57(2):293–302
Sadak MS, Abdelhamid MT, Schmidhalter U (2015) Effect of foliar application of aminoacids on plant yield and some physiological parameters in bean plants irrigated with sea water. Acta Biol Colomb 20(1):141–152
Shalata A, Neumann P (2001) Exogenous ascorbic acid (vitamin C) increases resistance to salt stress and reduces lipid peroxidation. J of Experimental Bot 52(364):2207–2211
Singh BK (1999) Plant amino acids: biochemistry and biotechnology. Marcel Dekker, New York, p 648
Smirnoff N (1993) The role of active oxygen in the response of plants to water deficit and desiccation. New Phytol 125:27–58
Sobhanian N, Pakniyat H, Kordshooli MA, Dorostkar S, Aliakbari M, Nasiri ZF (2016) Electrophresis study of wheat (Triticum aestivum L.) protein changes under salinity stress. Science Res 4(2):33–36
Steel RGD, Torrie JH, Dickey DA (1997) Principles and Procedures of Statistics. A Biometrical Approach, 3rd edn. McGraw Hill Book Co. Inc, New York
Takahashi H, Kopriva S, Giordano M, Saito K, Hell R (2011) Sulfur assimilation in photosynthetic organisms: molecular functions and regulations of transporters and assimilatory enzymes. Ann Rev Plant Biol 62:157–184
Thomas D, Erostus NN (2008) Soybean research in Africa for 30 years, by IITA. Research for Development, Nigeria
Vaidyanathan H, Sivakumar P, Chakrabarsty R, Thomas G (2003) Scavenging of reactive oxygen species in NaCl-stressed rice (Oryza sativa L.) differential response in salt-tolerant and sensitive varieties. Plant Science 165:1411–1418
Velikova V, Yordanov I, Edreva A (2000) Oxidative stress and some antioxidant systems in acid rain-treated bean plants. Protective role of exogenous polyamines. Plant Sci 5:59–66
Very AA, Robinson MF, Mansfield TA, Sanders D (1998) Guard cell cation channels are involved in Na induced stomatal closure in a halophyte. Plant J 14:509–521
Yamada M, Morishita H, Urano K, Shiozaki N, Yamaguchi-Shinozaki K, Shinozaki K, Yoshiba Y (2005) Effects of free proline accumulation in petunias under drought stress. J Exp Bot 56:1975–1981
Younis ME, Hasaneen MN, Kazamel AMS (2009) Plant growth, metabolism and adaptation to stress conditions. XXVII. Can ascorbic acid modify the adverse effects of NaCl and manitol on amino acids, nucleic acids and protein patterns in Vicia faba seedlings? Protoplasma 235:37–47
Acknowledgements
The authors are thankful to the National Research Centre for the provision of laboratory facilities to carry out this research.
Funding
There are currently no funding sources in the design of the study and the collection, analysis, and interpretation of data and in the writing of the manuscript.
Author information
Authors and Affiliations
Contributions
All authors proposed the research idea. MEE performed and designed the pot experiments. MSS performed the laboratory analysis and wrote the manuscript. MGD shared in coordinating the data collection and wrote the manuscript. ARA and FSA designed and performed the experiments and analyzed the data. All authors read and approved the final manuscript.
Authors’ information
Mervat Sh. Sadak, Mona G. Dawood, and Mohamed E. El-Awadi are professors of Plant Physiology at the National Research Centre, Egypt.
Asmaa R. Abd El-Hameid and Faten S.A. Zaki are researchers of Plant Physiology at the National Research Centre, Egypt.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable
Consent for publication
Not applicable
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Sadak, M.S., Abd El-Hameid, A.R., Zaki, F.S.A. et al. Physiological and biochemical responses of soybean (Glycine max L.) to cysteine application under sea salt stress. Bull Natl Res Cent 44, 1 (2020). https://doi.org/10.1186/s42269-019-0259-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s42269-019-0259-7