
Abstract
Background
The main objectives of this study were to compare two in vitro selection procedures for developing drought-tolerant cell lines from rice cultivars, regenerate plants from the selected cells, and use the random amplified polymorphic DNA (RAPD) method for genetic characterization of somaclons. Callus was derived from mature embryo cultures of tow Egyptian rice cultivars, Sakha 102 and Giza 178.
Results
The direct in vitro selection method indicated that the callus weight of the two cultivars was significantly decreased in response to increasing ABA concentrations. The shoot formation revealed significant differences among ABA concentrations and highly significant for interaction between rice cultivars and ABA concentrations. The percentage of shoot formation either increased or decreased with increasing ABA levels in rice cultivars. However, Giza 178 cultivar was slightly stable for shoot formation than the cultivar Sakha 102. The obtained results for stepwise selection method indicated also that the shoot formation was obtained till 6 mg/L ABA. The results of the RAPD analysis indicated that the rice cultivars and their somaclons were classified into tow clusters. The polymorphic information content (PIC) per primer ranged from 0.715 to 0.906 with an average of 0.860. The results of UPGMA clustering were consistent with those of the PCoA.
Conclusions
The percentage for Sakha 102 was 10.8, 7.7, and 2.2% on 2, 4, and 6 mg/L ABA, respectively, and was 16.4, 10.0, and 3.4% for Giza 178 cultivar, at the ABA concentrations of 2, 4, and 6 mg/L, respectively. The genetic similarity among the fifteen genotypes ranged from 0.30 to 0.82. Rice cultivars and their somaclons were classified into five clusters.
Similar content being viewed by others

Background
Rice (Oryza sativa L.) is one of the most important food crops, providing a stable diet for most of the world’s population. However, rice yield and quality are severely compromised by physiological and environmental stresses, as well as by insects and diseases. Rice production underminimizing the irrigation requirement became an objective in Egypt, as well as many areas worldwide, due to the increasing limitation on water supply.
Abiotic stresses resulting from drought, salinity, cold, and heat are the major environmental factors which affect plant growth and commonly constitute serious threats to agriculture (Lafitte et al. 2006; Verma et al. 2013; Alhasnawi et al. 2016; Banerjee et al. 2017; Jahan et al. 2017). Drought is a multifaceted stress condition with respect to timing and severity, ranging from long drought seasons where rainfall is much lower than demand to short periods without rain where plants depend completely on available soil water (Lafitte et al. 2006). However, water availability in the soil varies with respect to amount and distribution (Clark et al. 2002). Rice cultivation varies from flooded wetland to rain-fed dry land. Drought is the major environmental factor in rain-fed areas that occupy about one third of the total area and reduces productivity to 13–35% (Lafitte et al. 2006). The genotypic variation in drought tolerance together with the genetic tools available for rice (Matsumoto et al. 2005; Rensink and Buell 2005) and possibility to test their agronomic relevance (Xu et al. 2006) make rice a model system for research in drought tolerance. Studies of rice plants in response to drought stress and identification of drought tolerance mechanisms towards the development of drought-adapted crops can make more crops per drop a reality (Degenkolbe et al. 2009).
In vitro culture may be used to obtain drought-tolerant plants, assuming that there is a correlation between cellular and in vitro plant responses (Biswas et al. 2002; Vennapusa et al. 2015). This method is based on the induction of genetic variation among cells and tissues in cultured and regenerated plants, although there are genetic, biochemical, and physiological constraints in obtaining stress-tolerant plants through in vitro culture. Nabors and Dykes (1985) pointed out that this technique has been successfully used to produce stress-tolerant plants from several plant species. Recently, in vitro selection for cells, exhibiting increased tolerance to water stress and the successful regeneration of drought-tolerant plants, has been achieved (Vennapusa et al. 2015; Jahan et al. 2017).
Maintaining genetic uniformity is one of the major concerns of the tissue culture. RAPD polymorphism results from either a nucleotide base change that alters the primer binding site or from an insertion or deletion within the amplified region (Mahmood et al. 2010). Among the different tools available in molecular biology, RAPD is the simplest, cheaper, sensitive, and useful technique for the analysis of genetic fidelity of commercially micro-propagated plants (Chuang et al. 2009; Mallon et al. 2010; Verma et al. 2010; Elshafei et al. 2019) ranging from economically valuable clones (Valladares et al. 2006) to endangered plant species (Guo et al. 2006). Keeping the above facts in view, an attempt was made to standardize the protocol for somatic embryogenesis from mature embryo-derived calli of diverse rice varieties to determine the basis for somaclonal variation and to examine the responses of rice varieties to varying degrees of PEG-induced drought stress.
The objectives of this study were to (1) compare two in vitro selection procedures for developing drought-tolerant cell lines from rice cultivars, which had the high frequency of shoot formation, and to regenerate plants from the selected cells, and to (2) study the genetic differentiation among somaclons using the RAPD molecular marker.
Methods
Plant material and culture conditions
The two Egyptian rice cultivars, Sakha 102 and Giza 178, were used for in vitro selection methods. Both cultivars were chosen as they had the highest capacity to regenerate plants from mature embryo cultures. Only one medium protocol (Murashige and Skoog (MS) supplemented 2,4-D 1.0 Kinteine 1.2 mg/L for callus induction and 2,4-D 0.5 ABA 0.1 mg/L YE 1.36 g/L), which showed the highest potential for shoot formation, was used (Elshafei 2002).
Mature embryo culture was performed for the induction of somatic embryos. Mature rice seeds, dehusked manually, were washed with detergent Teepol for 2–5 min and thereafter with sterile distilled water. Seeds were then surface sterilized for 1 min in ethanol (70%), followed by sodium hypochlorite (1% w/v) for 15 min and then with mercuric chloride (0.1% w/v) for 10 min, and finally rinsed five times with sterile distilled water. The seeds were inoculated on MS (Murashige and Skoog 1962) supplemented 2,4-D 1.0 Kinteine 1.2 mg/L and solidified with 6 g L−1 agar to study the callogenic response in seed explants of rice. The cultures were incubated at 28 ± 2 °C with 16 h light and 8 h dark with 60 μmol m−2 s−1 light intensity provided by cool white fluorescent tubes (40 W; Phillips). The relative humidity within culture room was maintained at 60%. The callus induction frequency was measured as the percentage of seeds that produced a callus (Lee et al. 2009).
Selection methods
Direct selection and stepwise selection
In the direct selection method, the mature embryos were transferred to a medium containing 0, 2, 4, 6, 8, 10, and 12 mg/L abscisic acid (ABA). The calli were maintained on each respective treatment for two subcultures. The internal time (period) for each subculture was 5 weeks. The embryogenic calli were, then, transferred to the regeneration medium. The in vitro traits were recorded. In the stepwise selection method, the embryogenic calli, derived from mature embryos, were transferred to a medium containing 4 mg/L ABA. They were subcultured on a fresh medium with gradually increasing concentration of ABA by 2 mg/L until it reached 12 mg/L. In the meantime, eight replicates were taken into regeneration media of each ABA concentration. Then, the calli were transferred to the regeneration medium (2,4-D 0.5 ABA 0.1 mg/L YE 1.36 g/L) without ABA. Regenerated shoots were transferred to MS medium devoid of growth regulators for further development.
Callus induction and morphogenetic response
After two subcultures of incubation, callus induction and morphogenetic response were recorded and the following determinations were made for each petri dish: the percentage of embryo explants, which produced callus on the initiation medium; the fresh weight (mg) of the initiated callus; and the percentage of embryogenic callus, derived from the embryo explants. Regenerated shoots were transferred to MS medium devoid of growth regulators for further development.
Transfer of in vitro grown plantlets to soil
In vitro raised healthy plantlets were taken out from the culture bottles and washed gently with sterile water to remove the adhering medium completely. Thereafter, they were transferred to disposable thermocol glasses containing autoclaved soil mixture and vermiculite (3:1 v/v) and irrigated with 50 ml of 0.5× MS solution. The plantlets were covered with transparent polythene bags to prevent desiccation and were maintained in a greenhouse at a temperature of 28 ± 2 °C for 1 month. Completely developed plantlets were transferred to pots filled with 1:1 v/v mixture of soil and organic manure and were successfully established in field conditions.
Statistical analysis
Data were statistically analyzed as a 2-factor experiment (cultivars and ABA concentrations) in a completely randomized design (CRD) with eight replicates. All in vitro traits data, except for the callus weight, were subjected to arcsine transformation, before statistical analysis. Comparisons among means were made by using the LSD test of significance.
DNA extraction
PCR analyses were carried out by using the genomic DNA from the rice cultivars, Sakha 102 and Giza 178, and their somaclons lines, which were obtained by in vitro selection. Genomic DNA extraction of end cervical samples was performed using the Wizard Genomic DNA Purification Kit (Promega Biotechnology Corporation, Madison, USA). The extracted DNA samples were treated with RNase and stored at − 20 °C until further use.
PCR amplification
Nine primers (Table 3), obtained from Pharmacia Biotech (Amersham Pharmacia Biotech UK Limited, England HP79 NA), were tested in the present study to amplify the template DNA. Amplification reaction volumes were 25 μl, each containing 1× PCR buffer with MgCl2 (50 mM KCl, 10 mM Tris-HCl (pH = 9.0), 2 mM MgCl2, and 1% Triton X-100); 200 μM each of dATP, dCTP, dGTP and dTTP; 50 PM primer; 50 ng template DNA; and 1.5 μ of tag polymerase. Reaction mixtures were overlaid with 15 μl mineral oil and exposed to the following conditions: 94 °C for 3 min, followed by 45 cycles of 1 min at 94 °C, 1 min at 36 °C, 2 min at 72 °C, and a final 7 min extension at 72 °C. Amplification products were visualized with DNA marker on 1.6% agarose gel with 1× TBE buffer and were detected by staining with an ethidium bromide solution for 30 min. The amplified PCR products were applied to 2 % (m/v) agarose gel, containing 0.1 μg cm−3 ethidium bromide in TBE buffer. Gels were analyzed by UVI Geltec version 12.4; 1999-2005 (USA).
Data handling and cluster analysis
RAPD data were scored for computer analysis on the basis of the presence of the amplified products for each primer. If a product is present in a rice tow cultivars and their somaclons, it will be designated as “1,” if absent, it will be designated as “0,” after excluding the unreproducible bands. Pairwise comparisons of rice tow cultivars and their somaclons, based on the presence or absence of unique and shared polymorphic products, were used to determine similarity coefficients, according to Jaccard (Jaccard 1908). The similarity coefficients was used to construct dendograms, using the unweighted pair group method with arithmetic averages (UPGMA), employing the SAHN (sequential, agglomerative, hierarchical, and nested clustering) from the NTSYS-pc (Numerical Taxonomy and Multivariate Analysis System), version 1.80 (Applied Biostatistics) program (Rohlf 1993). Bootstrap analysis with PAUP version 4.0b10 software (Swofford 2003) was performed to evaluate the tree topology reliability for 1000 simulations to investigate the discriminatory power of each RAPD primer; the polymorphic information content (PIC) was calculated according to Smith et al. (2000) as follows: PIC = PIC = 1 − Σn fi2 where fi is the frequency of the ith allele in the set of 16 rice tow cultivars and their somaclons. Principal coordinate analysis (PCoA) from the RAPD was subsequently made using Jaccard modules in NTSYS-pc.
Results
Direct selection for tolerance to ABA
Analysis of variance of the effects for callus induction (%), callus weight (mg), embryogenic callus (%), shoot formation (%), root formation (%), and their interactions, presented in (Table 1), indicated that the percentage of callus induction was either significantly or highly significantly influenced by rice cultivars and ABA concentrations. The interaction was not significant between cultivars and ABA concentrations for callus induction. The analysis of variance for the effects of cultivars and ABA concentrations used, and their interaction, on callus weight is presented in Table 1. Such analysis indicates that the callus weight only was highly significantly affected by ABA concentrations. However, the rice cultivars and the interaction between them and ABA concentrations did not significantly affect their callus weight. The analysis of variance, presented in (Table 1), indicated that the percentage of embryogenic callus was either significantly or highly significantly affected by rice cultivars, ABA levels, and their interaction. The interaction was highly significant between rice cultivars and ABA concentrations (Table 1). It further reveals that shoot formation was significantly and highly significantly affected by ABA concentrations and their interaction with rice cultivars, respectively. However, the effect of rice cultivars was insignificant Table 1. The analysis of variance, presented in Table 1, indicated that the percentage of root formation was highly significantly influenced only by ABA concentrations, but was not significantly affected by either the rice cultivars or the interaction between the two tested variables.
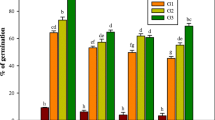
Means of callus induction (%), callus weight (mg), embryogenic callus (%), shoot formation (%), and root formation (%) as influenced by cultivar, ABA concentration, and their interaction were shown in Figs. 1 and 2 from which it is clear that the percentage of callus induction markedly decreased, in response to increasing ABA concentration. However, the reduction was slightly lower for the cultivar Giza 178 than for Sakha 102. Furthermore, the effects of increasing ABA concentrations on callus weight, for the two rice cultivars, are shown in Figs. 1 and 2.

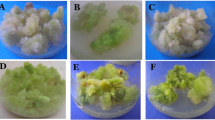
Callus induction, somatic embryogenesis, and plant regeneration derived from mature embryo explants of rice. a Yellow-white callus. b Nodular callus. c Shoot formation. d Whole plant regeneration at the flowering stage

Callus induction, callus weight, embryogenic callus, shoot formation, and root formation of rice cultivars transferred to MS after tow subcultures treatment with various A concentration. Bars with different lower case letters indicate a significant difference by LSD test (p < 0.05)
The callus weight markedly decreased with increasing ABA concentration. In fact, the reduction was almost similar for the two cultivars. The effects of increasing ABA concentration on the percentage of embryogenic callus, for the two rice cultivars, are shown in Figs. 1 and 2. It is obvious that the percentage of embryogenic callus either decreased or increased with different ABA levels in both cultivars. However, Sakha102 cultivar showed markedly the lowest response to the percentage of embryogenic callus (49.18%) in the case of 6 mg/L ABA level. The cultivar, Giza 178, showed the highest percentage to the embryogenic callus (64.25%) on the medium containing 6.0 mg/L ABA. However, no embryogenic calli (0.00 mg/L) were observed on the medium containing 12 mg/L ABA in Sakha 102 cultivar (Figs. 1 and 2).
The effect of increasing ABA concentration on the percentage of shoot formation, for the two cultivars, is shown in Fig. 2. Such figure clarifies that the percentage of shoot formation either increased or decreased with increasing ABA levels in both cultivars. However, Giza 178 cultivar was slightly less variable for shoot formation than Sakha 102 (Figs. 1 and 2). The root formation effects of increasing ABA concentration on such percentage either decreased or increased with increasing ABA concentrations in both cultivars. However, the highest percentage of root formation was obtained by the cultivar, Sakha 102, on the medium containing 8 mg/L ABA while, Giza 178 gave a high percentage of root formation on the medium of 4 mg/L ABA (Fig. 2).
Stepwise selection for tolerance to ABA
In the method of increasing ABA concentration, stepwise selection, on calli growth of rice cultivars, was examined. The calli showed a healthy and less variable growth till the medium contained 12 mg/L ABA. These calli did not exhibit any browning. Shoot formation was obtained till the medium contained 6 mg/L ABA. The percentages of shoot formation, derived from selected calli, were 10.8, 7.7, and 2.2% for Sakha 102 cultivar and were 16.4, 10.0, and 3.4% for Giza 178 cultivar at the ABA concentrations of 2, 4, and 6 mg/L, respectively (Table 2).
Plant regeneration from selected calli
The numbers of regenerated plants obtained from direct selection were 118 and 105 for the cultivars, Sakha 102 and Giza 178, respectively. The regenerated plants, derived from the selected cells, were grown to maturity and the seeds were obtained. The number of plants that gave seeds was 11 and 42 for Sakha 102 and Giza 178 cultivars, respectively, while the number of plants obtained from stepwise selection was 44 and 48 for Sakha 102 and Giza 178 cultivars, respectively.
RAPD analysis
Nine primers were used to amplify the genome of the two rice cultivars, Giza 178 and Sakha 102, and their somaclons. The nine primers studied amplified a total of 77 DNA fragments. Out of these 77 amplified fragments, 11.7% were not polymorphic, whereas, the remaining bands (88.3%) were polymorphic in one or other of the sixteen rice (two cultivars and their somaclons). The number of bands, amplified per primer, ranged from four (primer 5) to twelve (primer 8) with a mean value of 8.6 bands per primer (Table 3). All of the ten scorable bands were polymorphic across the rice genotypes by primer 9 (Fig. 3).

RAPD polymorphism in rice cultivars, Giza178 and Sakha102, and their somaclons using primer 9
The polymorphic information content (PIC) per primer ranged from 0.715 to 0.906 with an average of 0.860 (Table 3). The lowest value was recorded for RAPD5 primer. The highest value was recorded for RAPD8 (0.906). PIC values were positively correlated (r = 0.91) with a number of amplified alleles per primer.
Cluster analysis
One of the goals of the present study was to investigate the efficiency of RAPD markers in determining, accurately, the genetic relationship between somaclons and their parents. The RAPD markers, produced by nine primers, were used to construct a similarity matrix. Simple matching coefficient, ranging from 0.26 to 0.82, suggested a broad genetic base for rice genotypes. The results indicate the genetic similarity among the rice cultivars and their somaclons, based on the 68 polymorphic bands (Table 3) represents the clustering of rice cultivars generated by UPGMA analysis of the parents, namely Sakha 102, Giza 178, and their somaclons. Tow clusters could be observed. The first cluster comprised the parent, Giza 178, and its somaclons, while the second included the parent, Sakha102, and its somaclons (Fig. 4).

Dendrogram of rice cultivars and somaclons based on genetic distance obtained from RAPD markers using the UPGMA method. Numbers on branches correspond to bootstrap values % (1000 replications)
The dendrogram was confirmed by PCoA analysis (Fig. 5). The first five principal coordinates accounted for 87.6% of the total variation. Accessions in PCoA scatter plot (indicated by ellipses numbered with A, B, and C) seemed to form a very close grouping in the dendrogram (Fig. 5). Accessions clustered in ellipses A, B, and C were basically from group I and II of the dendrogram correspondingly.

The scatter plot of principal coordinate analysis (PCoA) based on the RAPD data. The accessions in ellipses A, B, C are from groups I and II
Discussions
Direct and stepwise selection for tolerance to ABA
Our results in this study were for the percentage of shoot formation either increased or decreased with increasing ABA levels in both cultivars. However, Giza 178 cultivar was slightly less variable for shoot formation than cultivar “Sakha 102” (Figs. 1 and 2). The stepwise method was more effective for shoot formation than the direct selection. This might be explained by the gradual adaptation of the tissue to the step by step increase of ABA concentration. Similar results have been obtained by Barakat and Abdel-Latif (1996), using three in vitro selection methods for NaCl tolerance in wheat callus. They reported that the stepwise method of increasing NaCl in the medium was more effective for plant regeneration than other methods. Previously, studies on in vitro selection for salt tolerance in rice were initiated by Yano and Ogawa (1982). They cultured immature embryos of IR36 cultivar on LS medium, supplemented with 17.5 to 67.5% seawater, and obtained callus showing various degrees of tolerance. After four subcultures (4 months), they obtained tolerant plants from 17.5–37.5% seawater medium. Likewise et al. (1983) observed that four subcultures, in media with different concentrations of seawater, were more than enough to produce salt-tolerant cells. The salt-tolerant progenies were evaluated (Wong et al. 1986).
In vitro screening of rice genotypes for drought tolerance using polyethylene glycol (Joshi et al. 2011) reported that mature embryos of three aromatic genotypes (Pusa Basmati 1, Pant Sugandh Dhan 17, Taraori Basmati) and one non-aromatic genotype (Narendra 359) indica rice (Oryza sativa L.) varieties were used for developing callus on Murashige and Skoog medium supplemented with 2,4-dichlorophenoxy acetic acid (2,4-D) (2.0 mg L−1 for Narendra 359 and 2.5 mg L for Pusa Basmati 1, Taraori Basmati, and Pant Sugandh Dhan 17). Screening of calli was done by subculturing calli for 15 days on Murashige and Skoog (MS) basal medium supplemented with different concentrations of polyethylene glycol (PEG)-6000 as chemical drought inducer. Callus volume decreased, and total proline content was found to be increased significantly with an increase in PEG concentration. Narendra 359 showed the best response in terms of callus growth at 70 g l−1 of PEG. The highest percentage somatic embryogenesis among selected calli was observed in Pusa Basmati 1 and the lowest in Pant Sugandh Dhan 17. Excellent shooting and rooting (94%) was observed in MS + 0.1 mg L−1 naphthalene acetic acid (NAA) and MS + 2.0 mg L−1 2,4-D (Joshi et al. 2011). Kacem et al. (2017) found that the data of three durum wheat indicated that embryogenic calli regenerated at a high frequency on the control medium, particularly Djenah Khetifa (60.7%)and Waha (58.1%). Oued Zenati recorded a mean of 49.2%. These results demonstrated the efficiency of the regene ratio polyethyleneglycol (PEG) on different concentrations of PEG (0%, 10%, and 20%). Masoabi et al. (2018) they reported that sugarcane callus from the NCo310 cultivar was subsequently mutagenized with 16 mM EMS and in vitro selected on 20% (w/v) PEG6000, which resulted in the survival of 18 plantlets. These in vitro selected lines were subjected to preliminary greenhouse pot trials to confirm drought tolerance. Pot trials identified seven lines that outlived NCo310 control plants. In addition, when re-watered after the drought stress period, plants from one mutant line recovered and were able to form new shoots.
Results in this study demonstrated that plant regeneration was not possible after four subcultures or from long-term cultured calli. Barakat and Cocking (1989) reported that cultured cells lost their capacity for plant regeneration after several subcultures. Cultured cells frequently contained nuclei, showing various degrees of polyploidy, aneuploidy, and chromosome abnormalities. This chromosomal variation often leads to loss of morphogenetic capacity.
Molecular markers analysis
The PCR technique proved to be a powerful tool for the identification of polymorphism in cereals. Using wheat, barley, rye, wheat-barley, and rice addition lines, Weining and Langridge (1991) detected polymorphism, using conserved, semi-random and random primers. With different combinations of primers, they were able to detect both inter- and intraspecific diversities. Ten primers were used in the genetic analysis of three aromatics (Pusa Basmati 1, Pant Sugandh Dhan 17, Taraori Basmati) and one non-aromatic (Narendra 359) indica rice (Oryza sativa L.) (Joshi et al. 2011). They reported that all primers showed a high level of polymorphism with all the somaclons. The results indicated that the somaclons of different varieties showed genetic similarity ranging from 0.46 to 0.76 (Joshi et al. 2011).
These results indicated that the RAPD technique could be successfully applied to crop species with very large genomes, like rice, to obtain a proper characterization of genetic relationship. Nagaoka and Ogihara (1997) stated that RAPD markers were more easily handled and, thus, became more desirable to estimate the genetic relationship among rice genotypes. Hu and Quiros (1991) analyzed the potential of using RAPD markers for cultivar identification in Brassica crops. They found that as few as four primers could be distinguished among fourteen broccoli and twelve cauliflower cultivars. Our results in this study were for the PCoA; the first five principal coordinate accounted for 87.6% of the total variation. The principal component analysis (PCoA) showed that 95.56% of the total variation was explained by the first two principal components Kacem et al. 2017.
In the PCoA results by Hassanein and Al-Soqeer (2018), genotypes were divided into two main clusters presenting Mo genotypes and Mp genotypes. The (PCoA) plot for the first two coordinates explains 51.68% of variance among genotypes. Elshafei et al. (2019) found that genotypes were divided into two main clusters presenting salt-tolerant genotypes and salt-sensitive bred wheat genotypes.
Conclusion
In the selection for tolerance to ABA, the calli showed a healthy and stable growth till the medium contained 12 mg/L ABA. The shoot formation was obtained till 6 mg/L ABA. The percentage of shoot formation, derived from selected calli, was 10.8, 7.7, and 2.2% for Sakha 102 cultivar and was 16.4, 10.0, and 3.4% for Giza 178 cultivar, at the ABA concentrations of 2, 4, and 6 mg/L, respectively. The RAPD markers produced by nine primers were used to construct a similarity matrix. The genetic similarity among the fifteen genotypes ranged from 0.30 to 0.82. Rice cultivars and their somaclons were classified into five clusters. The present study indicated that the use of RAPD technique was sensitive and powerful to detect genetic variation at the level of DNA among rice cultivars and their somaclons. This might be of particular importance in the future, dealing with in vitro selection to stress conditions.
Availability of data and materials
The authors declare that the experimental data and material are available.
Abbreviations
- 2,4-D:
-
2,4-Dichlorophenoxyacetic
- ABA:
-
Abscisic acid
- BA:
-
N6-Benzyl adenine
- CRD:
-
Completely randomized design
- IBA:
-
Indole-3-butyric acid
- ISSR:
-
Inter-simple sequence repeats
- KN:
-
6-Furfuryl aminopurine
- MS:
-
Murashige and Skoog
- NAA:
-
1-Naphthaleneacetic acid
- PCoA:
-
Principal coordinate analysis
- PCR:
-
Polymerase chain reaction
- PEG:
-
Polyethylene glycol
- PIC:
-
Polymorphic information content
- RAPD:
-
Random amplified polymorphic DNA
- SSR:
-
Simple sequence repeat
- UPGMA:
-
Unweighted pair group method with arithmetic averages
References
Alhasnawi AN, Radziah CC, Kadhimi AA, Isahak A, Mohamad A, Yusoff WMW (2016) Enhancement of antioxidant enzyme activities in rice callus by ascorbic acid under salinity stress. Biol Plant 60(4):783–787
Banerjee J, Gantait S, Maiti MK (2017) Physiological role of rice germin-like protein 1 (OsGLP1) at early stages of growth and development in indica rice cultivar under salt stress condition. Plant Cell, Tissue Organ Cult (PCTOC) 131(1):127–137
Barakat MN, Abdel-Latif TH (1996) In vitro selection of wheat callus tolerant to high levels of salt and plant regeneration. Euphytica 91:127–140
Barakat MN, Cocking EC (1989) Regeneration of plants from Linum protoplast. In: Biotechnology in agriculture and forestry & (plant protoplast and genetic engineering I), pp 147–159
Biswas J, Chowdhury B, Bhattacharya A, Mandal AB (2002) In vitro screening for increased drought tolerance in rice. In Vitro Cell Dev Biol-Plant 38:525–530
Chuang SJ, Chen CL, Chen JJ, Chou WY, Sung JM (2009) Detection of somaclonal variation in micro-propagated Echinacea purpurea using AFLP marker. Sci Hort 120:121–126
Clark LJ, Cope RE, Whalley WR, Barraclough PB, Wade LJ (2002) Root penetration of strong soil in rain fed lowland rice: comparison of laboratory screens with field performance. Field Crops Res 76:189–198
Degenkolbe T, Do PT, Zuther E, Repsilber D, Walther D, Hincha DK, Kohl KI (2009) Expression profiling of rice cultivars differing in their tolerance to long-term drought stress. Plant Mol Biol 69:133–153
Elshafei AA (2002) Towards in vitro selection studies for drought tolerance in rice. Master of Science Alexandria University, Alexandria
ELshafei AA, Afiah SA, Amer MA, El-enany MAM (2019) Validation of molecular markers linked with salinity tolerance and assessment of multiple salinity tolerance indices in wheat genotypes grown on saline soil (Triticum aestivum L.). Biosci Res 16(2):963–978
Guo WL, Gong L, Ding ZF, Li YD, Li FX, Zhao SP, Liu B (2006) Genomic instability in phenotypically normal regenerants of medicinal plant Codonopsis lanceolata Benth. et Hook. f., as revealed by ISSR and RAPDs markers. Plant Cell Rep 25:896–906
Hassanein AMA, Al-Soqeer AA (2018) Morphological and genetic diversity of Moringaoleifera and Moringaperegrina genotypes. Horticulture Environment, and Biotechnology (19):1–11
Hu J, Quiros CF (1991) Identification of broccoli and cauliflower cultivars with RAPD markers. Plant Cell Rep 10:505–511
Jaccard P (1908) Nouvelles rechearches sur la distribution lorale. Bull Soc Vaud Sci Nat 44:223–270
Jahan N, Javed MA, Manan FA, Khan S, Samad AA, Zaidi MI (2017) Comparative study of physiological and growth parameters for sodium and iron toxicities in upland and lowland cultivars of Oryza sativa L. Pak J Bot 49:249–254
Joshi R, Alok S, Raj KS (2011) In vitro screening of rice genotypes for drought tolerance using polyethylene glycol. Acta Physiol Plant 33:2209–2217
Kacem NS, Delporte F, Muhovski Y, Djekoun A, Watillon B (2017) In vitro screening of durum wheat against water-stress mediated through polyethylene glycol. J Genet Eng Biotechnol 15(1):239–247
Lafitte HR, Li ZK, Vijayakumar CHM, Gao YM, Shi Y, Xu JL, Fu BY, Yu SB, Ali AJ, Domingo J, Maghirang R, Torres R, Mackill D (2006) Improvement of rice drought tolerance through backcross breeding: evaluation of donors and selection in drought nurseries. Field Crops Res 97:77–86
Lee KW, Choi GJ, Kim KY, Ji HC, Park HS, Yoon SH, Lee SH (2009) High frequency plant regeneration from mature seed derived callus of Italian ryegrass (Lolium multiflorum) cultivars. Afr J Biotech 8:6828–6833
Likewise Y, Ogawa YM, Yano S (1983) In cell and tissue culture techniques for cereal crop improvement. Science Press, Beijing, pp 237–254
Mahmood T, Nazar N, Abbasi BH, Khan MA, Ahmad M, Zafar M (2010) Detection of somaclonal variations using RAPD fingerprinting in Silybum marianum (L.). J Med Plants Res 4:1822–1824
Mallon R, Rodrı’guez-Oubin˜a J, Gonza’lez ML (2010) In vitro propagation of the endangered plant Centaurea ultreiae: assessment of genetic stability by cytological studies, flow cytometry and RAPD analysis. Plant Cell Tissue Organ Cult 101:31–39
Masoabi M, Lloyd J, Kossmann J, van der Vyver C (2018) Ethyl methanesulfonate mutagenesis and in vitro polyethylene glycol selection for drought tolerance in sugarcane (Saccharum spp.). Sugar Tech 20(1):50–59
Matsumoto T, Wu JZ, Kanamori H (2005) The map-based sequence of the rice genome. Nature 436:793–800
Murashige T, Skoog F (1962) A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol Plant 15:473–497
Nabors MW, Dykes TA (1985) Tissue culture of cereal cultivars with increased salt, drought and acid tolerance. Biotechnol Int Agric Res:121–1338
Nagaoka T, Ogihara Y (1997) Applicability of inter-simple sequence repeat polymorphisms in wheat for use as DNA markers in comparison to RFLP and RAPD markers. Theor Appl Genet 94:597–602
Rensink WA, Buell CR (2005) Microarray expression profiling resources for plant genomics. Trends Plant Sci 10:603–609
Rohlf FJ (1993) NTSYS-pc numerical taxonomy and multivariate system, version 1.80. Applied biostatistics Inc, New York
Smith JS, Kresovich CS, Hopkins MS, Mitchell SE, Dean RE, Woodman WL, Lee M, Porter K (2000) Genetic diversity among elite sorghum inbred lines assessed with simple sequence repeats. Crop Sci 40:226–232
Swofford DL (2003) PAUP. Phylogenetic analysis using parsimony (and other methods). Version 4.0b10. Sinauer Associates, Sunderland (MA)
Valladares S, Sa’nchez C, Martı’nez MT, Ballester A, Vieitez AM (2006) Plant regeneration through somatic embryogenesis from tissues of mature oak trees: true-to-type conformity of plantlets by RAPD analysis. Plant Cell Rep 25:879–886
Vennapusa AR, Vemanna RS, Bh RR, Babitha KC, Kiranmai K, Nareshkumar A, Sudhakar C (2015) An efficient callus induction and regeneration protocol for a drought tolerant rice indica genotype AC39020. J Plant Sci 3(5):248
Verma D, Ansari MW, Agrawal GK, Rakwal R, Shukla A, Tuteja N (2013) In vitro selection and field responses of somaclonal variant plants of rice cv PR113 for drought tolerance. Plant Signal Behav 8(4):e23519
Verma P, Srivastava P, Singh SC, Mathur A (2010) Genetic fidelity of long-term micropropagated plantlets of Valeriana wallichii—an endangered medicinal plant. Cur Sci 99:436–438
Weining S, Langridge P (1991) Identification and mapping of polymorphism in cereals based on the polymerase chain reaction. Theor Appl Genet 82:209–216
Wong CK, Sc W, Ko S (1986) Production of rice plantlets on NaCl – stressed medium and evaluation of their progenies. Bot Bull Academia Sinica 27:11–23
Xu K, Xu X, Fukao T, Canlas P, Maghirang-Rodriguez R, Heuer S, Ismail AM, Bailey-Serres J, Ronald PC, Mackill DJ (2006) Sub1A is an ethylene-response factor-like gene that confers submergence tolerance to rice. Nature 442:705–708
Yano SM, Ogawa YY (1982) Plant formation from selected rice cells resistant to salt. In: Fujiwara A (ed) Plant Tissue Culture. Maruzen, Tokyo, pp 495–496
Acknowledgements
The authors thank the Deanship of Scientific Research and RSSU at King Saud University for their technical support. Great appreciations are also due to Prof. M.I. Shaalan and Prof. M. Motawei for their suggestions and proofreading this manuscript.
Funding
This study was funded by Biotechnology Lab, Crop Science Department, Faculty of Agriculture, Alexandria University.
Author information
Authors and Affiliations
Contributions
AE performed the analysis with the molecular markers. MB and SM performed the tissue culture experiments. SK and MAM performed the statistical analysis. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
The authors declare that the work is ethically approved and consented to participate.
Consent for publication
The authors declare that the work has been consented for publication.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Elshafei, A., Barakat, M., Milad, S. et al. Regeneration of rice somaclons tolerant to high level of abscisic acid and their characterization via RAPD markers. Bull Natl Res Cent 43, 107 (2019). https://doi.org/10.1186/s42269-019-0154-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s42269-019-0154-2