
Abstract
Background
The pine processionary moth, Thaumetopoea wilkinsoni Tams, 1926 (Lepidoptera/Notodontidae) is one of the most harmful insects that destroys pine ecosystems by feeding on pine leaves at its larval stage. Because of its urticating setae, the insect also causes severe skin reactions to animals and humans. Instead of chemical control, eco-friendly biological control methods are preferred to combat this species.
Results
The purpose of this study was to evaluate the efficacy of five different Beauveria bassiana Vuill, 1912 (Hypocreales/Cordycipitaceae) isolates (TR-SM-10, TR-SM-11, TR-SM-2, TR-SK-1 and TR-D-1) and one B. pseudobassiana Rehner & Humber (Hypocreales/Cordycipitaceae) isolate (TR-SM-1) against the fourth instar larvae of T. wilkinsoni under laboratory conditions. T. wilkinsoni larvae were collected from the Ondokuz Mayıs University Kurupelit Campus in Samsun, Turkey, in 2021, and the fourth instar larvae were used in the experiment. Two ml of spray of the six fungal isolates were applied to every ten larvae at each concentration (1 × 107 and 1 × 108 conidia ml−1). The experiment was carried out in five replicates per group, and the larvae were observed for 10 days. As a result, all isolates of B. bassiana caused 100% mortality at 1 × 108 conidia ml−1 concentration. B. pseudobassiana isolate also caused 100% mortality at both concentrations. At 1 × 107 conidia ml−1 concentration, the larvae treated with the B. pseudobassiana isolate (TR-SM-1) had the lowest LT50 (2.89 days) and LT90 values (4.79 days), while the larvae treated with TR-SM-10 isolate had the highest LT50 (5.65 days) and LT90 values (9.39 days). At 1 × 108 conidia ml−1 concentration, the larvae treated with TR-SK-1 isolate had the lowest LT50 (2.89 days) and LT90 values (4.79 days), while those treated with TR-SM-10 isolate had the highest LT50 (3.95 days) and LT90 values (8.15 days).
Conclusion
It has been recommended that the five different isolates of B. bassiana and B. pseudobassiana isolates were virulent to T. wilkinsoni larvae and can be used for biological control of T. wilkinsoni.
Similar content being viewed by others
Background
The Pinaceae family has many genera, one of the most important genera is Pinus. The needles of the Pinus genus contain essential oils. Because of their various pharmacological properties, such as anti-ageing and anti-inflammatory effects, the needles are widely used in folk medicine and as food additives.
The Pinus species are distributed almost all over the world, including Turkey. Insect pests lead to the biological factors that threaten the continuity of pine forests in Turkey. The pine processionary moth, Thaumetopoea wilkinsoni Tams, 1926 (Lepidoptera/Notodontidae) is one of the most harmful needle-eating insects in pine ecosystems (Altunışık and Avcı 2016). The pest causes deformities in seedlings and stunting (Onaran and Katı 2010) and even tree deaths in young stands (Keleş et al. 2018). In addition, the larvae of the insect have urticating setae that cause skin reactions in both animals and humans (Rodriguez-Mahillo et al. 2012).
Both mechanical and chemical control methods are used to combat the pest in pine forests in Turkey. Given the adverse effects of chemical control on the ecosystem, biological control methods are preferred to combat the species. The use of entomopathogenic microorganisms as a biological control agent is critical in overcoming harmful species in terms of both targeting only a specific pest and being environmentally friendly. Entomopathogenic fungi (EPF) are one of the most effective biological control agent in pest control. B. bassiana Vuill, 1912 (Hypocreales/Cordycipitaceae), is the most common one (Zibaee et al. 2013). Studies on B. bassiana (Rehman et al. 2020) have revealed that the fungi are effective against a variety of insects. Furthermore, the effectiveness of B. pseudobassiana Rehner & Humber (Hypocreales/Cordycipitaceae), which is a cryptic species morphologically similar to B. bassiana but phylogenetically distant from B. bassiana (Rehner et al. 2011), has also been investigated against various insects (Bedini et al. 2018). There are various studies investigating the use of different B. strains to develop safe biological control strategies in the control of the pine processionary moth (Gök et al. 2018).
Combating with T. wilkinsoni is critical for protecting pine forests, which are important for both ecological and economic reasons, as well as reducing the threat to human and animal health. Therefore, in the present study, the efficacy of five different B. bassiana isolates was evaluated against the fourth instar larvae of T. wilkinsoni under laboratory conditions.
Methods
Sampling
The fourth instar larvae of T. wilkinsoni were collected from a pine forest at Ondokuz Mayıs University Kurupelit Campus in Samsun, Turkey (N 41° 22′ 26.5116″ E 36° 13′ 17.6340″), in 2021.
Preparation of entomopathogenic fungi
A total of six native EPF isolates (TR-SM-10, TR-SM-11, TR-SM-2, TR-SK-1, TR-D-1, and TR-SM-1) molecularly identified and isolated from infected Palomena prasina Linnaeus (Hemiptera/Pentatomidae) in hazelnuts orchards, Black Sea region of Turkey in 2018–2019, were used in this study (Table 1) (Ozdemir et al. 2021). All isolates were obtained from the same host and the same location.
The six native EPF isolates belong to B. bassiana (TR-SM-10, TR-SM-11, TR-SM-2, TR-SK-1, and TR-D-1) and B. pseudobassiana (TR-SM-1). These isolates were inoculated into potato dextrose agar (PDA; Merck Ltd., Darmstadt, Germany) and incubated in complete darkness at 25 ± 1 °C for one week (Incubator; Binder KBWF 240; Germany). At the end of the growth period, each Petri dish was filled with sterile distilled water containing 0.02% Tween 20 and scraped with a glass bag to pass the conidia into the water. Following that, the conidia suspensions were filtered through three layers of sterile cheesecloth to remove mycelium and homogenized for 3 min by vortexing. The resulting conidia suspensions were adjusted to concentrations of the 1 × 107 and 1 × 108 conidia ml−1 using a Neubauer hemocytometer under Olympus CX31 light microscope (Olympus America Inc., Lake Success, NY) (Tuncer et al. 2019).
Application of entomopathogenic fungi on T. wilkinsoni
In the experiment, two layers of sterile filter paper were lightly moistened and placed in 1-L plastic cups. Two ml of the 1 × 107 and 1 × 108 conidia ml−1 were sprayed on the fourth instar larvae placed in plastic cups (ten larvae per dish) using a Potter spraying tower (Burkard, Rickmansworth, Hertz UK), and the appropriate amount of sterilized pine leaves were placed in the dishes to feed the larvae. After each application of EPF suspension, the spray tower was cleaned by 70% ethanol and sterile distilled water. Subsequently, the fourth instar larvae of the insect were released in plastic cups. Only sterile-distilled water containing 0.02% Tween 20 was sprayed to the control group. For 10 days, all plastic cups were incubated at 25 ± 1 °C, 70 ± 5% RH, and a 16:8 h light/dark period. The mortality rates were monitored for ten consecutive days. The dead larvae in each cup were counted 10 days after the application, and the mortality rates were determined. The same procedure was repeated every day for the control groups (Ozdemir et al. 2019).
Statistical analyses
Daily mortality rates were corrected using the Abbott formula when the mortality rate in the control group exceeded 5% (Abbott 1925). The LT50 and LT90 values were determined by Probit analysis using the log-Probit method (POLO-PLUS ver.2.0). The slopes of the regression lines were compared to each. The LT50 and LT90 values of the isolates were compared using confidence intervals (95%).
Results
Before bioassays, the conidial viability of B. bassiana (5) and B. pseudobassiana (1) isolates was determined, as well as the germination rate (100%). Beauveria isolates were applied to the fourth instar larvae of T. wilkinsoni at different concentrations, and the LT50 and LT90 values for each isolates differed (Tables 1 and 2). The LT50 and LT90 values for TR-SM-1 isolate against T. wilkinsoni were 2.89 and 4.79 days, respectively, at 1 × 107 conidia ml−1 concentration, whereas they were 3.01 and 7.47 days at 1 × 108 conidia ml−1. The LT50 and LT90 values for TR-D-1 isolate were 4.42 and 7.46 days at 1 × 107 conidia ml−1 while 3.27 and 6.39 days at 1 × 108 conidia ml−1, respectively. While the LT50 and LT90 values of TR-SK-1 isolate were 4.23 and 8.68 days at 1 × 107 conidia ml−1 concentration, the LT50 and LT90 values were 2.89 and 4.79 days at 1 × 108 conidia ml−1 concentration, respectively. The LT50 values of the 1 × 107 and 1 × 108 conidia ml−1 concentration of TR-SM-2 isolate were 4.22 and 3.64 days, whereas the LT90 values of the same concentrations of the same isolate were 7.46 and 6.99 days, respectively. Similarly, the LT50 values of the 1 × 107 and 1 × 108 conidia ml−1 concentrations of TR-SM-10 isolate were 5.65 and 3.95 days, respectively. The LT90 value of the 1 × 107 conidia ml−1 concentration of the same isolate was 9.39 days, while the LT90 value for the 1 × 108 conidia ml−1 of the same isolate was 8.15 days. The LT50 and LT90 values for TR-SM-11 isolate at the 1 × 107 conidia ml−1 concentration were 4.01 and 8.57 days, whereas at 1 × 108 conidia ml−1 for the same isolate, they were 3.75 and 7.66 days, respectively (Tables 2 and 3).
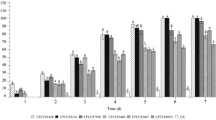
It was determined that the two tested concentrations of nearly all isolates began to cause mortality 2 days after the application, and the mortality increased with time (Figs. 1 and 2). Except for TR-SM-10, TR-SM-11 and TR-SK-1 isolates, the 1 × 107 conidia ml−1 concentration of the isolates caused 100% mortality 10 days after the application, while the 1 × 108 conidia ml−1 concentration of all isolates caused 100% mortality. Although TR-D-1 and TR-SM-10 isolates caused 100 and 93.3% mortality (at 1 × 107 conidia ml−1 concentration), respectively, at 1 × 108 conidia ml−1 concentration, the two isolates caused 100% mortality 5 and 9 days after the treatment. Similarly, while the mortality rates by the two isolates, TR-SM-11 and TR-SK-1 at 1 × 107 conidia ml−1 concentration, were 96% within 10 days, these rates for the two isolates at 1 × 108 conidia ml−1 concentration were 100% 7 days after the treatment. The mortality rates in both concentrations of TR-SM-1 were recorded as 100% 10 days after the treatment. Similarly, the 1 × 107 and 1 × 108 conidia ml−1 concentrations of TR-SM-1 isolate caused 100% mortality within 10 and 6 days, respectively (Figs. 1 and 2).

Mortality rates of Thaumetopoea wilkinsoni treated with Beauveria bassiana and Beauveria pseudobassiana isolates at 1 × 107 conidia ml−1 concentration

Mortality rates of Thaumetopoea wilkinsoni treated with Beauveria bassiana and Beauveria pseudobassiana isolates at 1 × 108 conidia ml−1 concentration
Discussion
B. bassiana, as one of the EPFs, is one of the most preferred species in terms of its effectiveness in pest control. The efficacy of five different B. bassiana and one B. pseudobassiana isolates against the fourth instar larvae of T. wilkinsoni was investigated. Since the pine processionary moth often overwinters as the fourth instar larvae (Ozdemir et al. 2019), the fourth instar larvae were chosen in this study.
It is critical to determine the lethal times (LT50 and LT90 values) to reduce the overall cost of pest control while maintaining a high level of control. While the most effective isolate was TR-SM-1 at 1 × 107 conidia ml−1 concentration, TR-SK-1 was the most effective isolate at 1 × 108 conidia ml−1 concentration. Both the LT50 and LT90 values decreased with the increasing conidial concentrations in all isolates (except TR-SM-1). This situation has indicated that the highest conidial concentrations of B. bassiana isolates were more virulent for T. wilkinsoni larvae. The results of the previous studies showed that they were consistent with the findings obtained from this study (Tuncer et al. 2019). Ozdemir et al. (2019) determined that the B. bassiana strain applied to T. pityocampa Denis & Schiffermüller, 1775 (Lepidoptera/Notodontidae), larvae caused a decrease in LT50 values with increasing conidial concentration. Similar results were obtained in studies where B. bassiana was treated with insect species from other orders other than Lepidoptera.
Ten days after the application, it was determined that B. bassiana isolates, except TR-SM-10, TR-SM-11, and TR-SK-1, caused 100% mortality at 1 × 107 conidia ml−1 concentration. All three isolates led to high mortality rates of 93.3, 96, and 96%, respectively. At the end of the experiment, it was determined that all isolates caused 100% mortality at 1 × 108 conidia ml−1 concentration. This situation proved that the concentration of the 1 × 108 conidia ml−1 for all isolates was highly virulent to T. wilkinsoni larvae. There are several studies showing the effectiveness of different Beauveria strains against the pine processionary moth. Draganova et al. (2013) determined that the pine processionary moth was affected by the B. bassiana strain. Sönmez et al. (2017) applied B. bassiana KTU-24 strain to the pine processionary moth, and noted that the strain caused more mortality with increasing concentration. Gök et al. (2018) showed that the B. bassiana LD2016 strain sprayed on the pine processionary moth caused 100% death on the fifth day, while the B. bassiana BMAUMM6-4 strain caused 100% death on the third day. In studies with different species, the efficacy of B. bassiana isolates was evaluated.
Studies on the effectiveness of B. pseudobassiana in insects of the order Lepidoptera are few. Of these, Schemmer et al. (2016) reported that six different B. pseudobassiana strains were applied to Cameraria ohridella Deschka & Dimić (Lepidoptera/Gracillariidae), and all strains caused 100% mortality on the tenth day. Gürlek et al. (2018) stated that B. pseudobassiana caused death in Cydia pomonella Linnaeus (Lepidoptera/Tortricidae). In addition, the effectiveness of B. pseudobassiana against insects in other orders was also discussed in the studies. B. pseudobassiana has been reported to be an entomopathogen for Hypera postica Gyllenhal (Coleoptera/Curculionidae) (Yücel et al. 2018). Álvarez-Baz et al. (2015) demonstrated that B. pseudobassiana was highly virulent against Monochamus galloprovincialis Olivier (Coleoptera/Cerambycidae), the vector of pine wilt disease. B. pseudobassiana isolates have been reported to be virulent against the Eastern pine weevil Pissodes nemorensis Germar (Coleoptera/Curculionidae) (Romon et al. 2017).
Conclusions
T. wilkinsoni is an important forest pest that causes significant allergic reactions in humans and animals. Biological control has an important potential in order to protect the ecosystem against T. wilkinsoni, which is common in forests. The pest is a candidate for the use of EPF with high humidity and low average temperatures. Beauveria isolates obtained from this region were applied against the pest and five different isolates of B. bassiana and B. pseudobassiana isolate were found to be virulent against T. wilkinsoni larvae in this study. Moreover, the study revealed that B. pseudobassiana, which is less studied compared to B. bassiana, is a promising entomopathogen and can be used to control T. wilkinsoni larvae. As a result, these fungal isolates can be applied in the field under controlled conditions.
Availability of data and materials
The data generated and/or analyzed during the current study are available from the corresponding author.
Abbreviations
- EPF:
-
Entomopathogenic fungi
- PDA:
-
Potato dextrose agar
References
Abbott WS (1925) A method of computing the effectiveness of insecticides. J Econ Entomol 18:265–267. https://doi.org/10.1093/jee/18.2.265a
Altunışık A, Avcı M (2016) Impact of pine processionary moth defoliation (Thaumetopoea wilkinsoni Tams, 1926) (Lep.: Notodontidae) on increment in Isparta pine forests. Turk Bull Entomol 6(3):231–244. https://doi.org/10.16969/teb.29367
Álvarez-Baz G, Fernández-Bravo M, Pajares J, Quesada-Moraga E (2015) Potential of native Beauveria pseudobassiana strain for biological control of pine wood nematode vector Monochamus galloprovinciali. J Invertebr Pathol 132:48–56. https://doi.org/10.1016/j.jip.2015.08.006
Bedini S, Sarrocco S, Baroncelli R, Vannacci G, Conti B (2018) Pathogenic potential of Beauveria pseudobassiana as bioinsecticide in protein baits for the control of the medfly Ceratitis capitata. Bull Insectology 71(1):31–38
Draganova S, Takov D, Pilarska D, Doychev D, Mirchev P, Georgiev G (2013) Fungal pathogens on some Lepidopteran forest pests in Bulgaria. Acta Zool Bulg 65(2):179–186
Gök S, Güven O, Karaca I (2018) Effects of the entomopathogenic fungus Beauveria bassiana on different stages of the pine processionary moth (Thaumetopoea wilkinsoni Tams). Turk J Biol Control 9(1):7–18. https://doi.org/10.31019/tbmd.436218
Gürlek S, Sevim A, Sezgin FM, Sevim E (2018) Isolation and characterization of Beauveria and Metarhizium spp. from walnut fields and their pathogenicity against the codling moth, Cydia pomonella (L.) (Lepidoptera: Tortricidae). Egypt J Biol Pest Control 28:50. https://doi.org/10.1186/s41938-018-0055-y
Keleş C, Voyvot S, Bilgili E (2018) The structure of egg batches, number of eggs and egg hatching rates of pine processionary moth Thaumetopoea pityocampa (Denis & Schiffermüller, 1775)/Thaumetopoea wilkinsoni Tams, 1924 (Lepidoptera: Notodontidae). JNHE 4:1–7. https://doi.org/10.21324/dacd.446545
Onaran MA, Katı M (2010) Biological struggle against pine bag insect (Thaumetopoea pityocampa Schiff). BAÜ Fen Bil Enst Derg 12(2):21–27
Ozdemir IO, Kushiyev R, Erper I, Tuncer C (2019) Efficacy of entomopathogenic fungi Beauveria bassiana and Metarhizium anisopliae against Thaumetopoea pityocampa Shiff. (Lepidoptera: Thaumatopoeidae). Arch Phytopathol Plant Prot 52(5–6):470–480. https://doi.org/10.1080/03235408.2019.1649812
Ozdemir IO, Tuncer C, Ozer G (2021) Molecular characterisation and efficacy of entomopathogenic fungi against the Green shield bug Palomena prasina (L.) (Hemiptera: Pentatomidae) under laboratory conditions. Biocontrol Sci Technol. https://doi.org/10.1080/09583157.2021.1944608
Rehman H, Rasul A, Farooqi MA, Aslam HMU, Majeed B, Sagheer M, Ali Q (2020) Compatibility of some botanicals and the entomopathogenic fungus, Beauveria bassiana (Bals.), against the red flour beetle, Tribolium castaneum (Herbst) (Coleoptera: Tenebrionidae). Egypt J Biol Pest Cont 30:131
Rehner SA, Minnis AM, Sung GH, Luangsaard JJ, Devotto L, Humber RA (2011) Phylogeny and systematics of the anamorphic, entomopathogenic genus Beauveria. Mycologia 103(5):1055–1073. https://doi.org/10.3852/10-302
Rodríguez-Mahillo AI, Gonzalez-Muñoz M, Vega JM, López JA, Yart A, Kerdelhué C, Camafeita E, Ortiz JCG, Vogel H, Toffolo EP, Zovi D, Battisti A, Moneo I (2012) Setae from the pine processionary moth (Thaumetopoea pityocampa) contain several relevant allergens. Contact Derm 67(6):367–374. https://doi.org/10.1111/j.1600-0536.2012.02107.x
Romon P, Hatting H, Goldarazena A, Iturrondobeitia JC (2017) Varriation in virulence of Beauveria bassiana and B. pseudobassiana to the pine weevil Pissoddes nemorensis in relation to mycelium characteristics and virulence genes. Fungal Biol 121(2):189–197. https://doi.org/10.1016/j.funbio.2016.11.008
Schemmer R, Chládeková P, Medo J, Barta M (2016) Natural prevalence of entomopathogenic fungi in hibernating pupae of Cameraria ohridella (Lepidoptera: Gracillariidae) and virulence of selected isolates. Plant Prot Sci 52:199–208. https://doi.org/10.17221/110/2015-PPS
Sönmez E, Demir İ, Bull JC, Butt TM, Demirbağ Z (2017) Pine processionary moth (Thaumetopoea pityocampa, Lepidoptera: Thaumetopoeidae) larvae are highly susceptible to the entomopathogenic fungi Metarhizium brunneum and Beauveria bassiana. Biocontrol Sci Technol 27(10):1168–1179. https://doi.org/10.1080/09583157.2017.1387643
Tuncer C, Kushiyev R, Erper I, Ozdemir IO, Saruhan I (2019) Efficacy of native isolates of Metarhizium anisopliae and Beauveria bassiana against the invasive ambrosia beetle, Xylosandrus germanus Blandford (Coleoptera: Curculionidae: Scolytinae). Egypt J Biol Pest Cont 29(1):28. https://doi.org/10.1186/s41938-019-0132-x
Yucel B, Gozuacik C, Gencer D, Demir I, Demirbag Z (2018) Determination of fungal pathogens of Hyperia postica (Gyllenhall) (Coleoptera: Curculionidae): isolation, characterization, and susceptibility. Egypt J Biol Pest Cont 28:39
Zibaee A, Bandani AR, Sendi J (2013) Pathogenicity of Beauveria bassiana to fall webworm (Hyphantria cunea) (Lepidoptera: Arctiidae) on different host plants. Plant Prot Sci 49(4):169–176. https://doi.org/10.17221/72/2012-PPS
Acknowledgements
Not applicable.
Funding
Not applicable.
Author information
Authors and Affiliations
Contributions
EFT, OY, CT, IOO, and EY conceived, designed, analyzed, wrote, corrected and approved the final draft. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Topkara, E.F., Yanar, O., Tuncer, C. et al. Efficacy of Beauveria bassiana and Beauveria pseudobassiana isolates against the pine processionary moth, Thaumetopoea wilkinsoni Tams, 1926 (Lepidoptera/Notodontidae). Egypt J Biol Pest Control 32, 3 (2022). https://doi.org/10.1186/s41938-021-00501-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s41938-021-00501-7