
Abstract
Background
Bacillus thuringiensis is a Gram-positive, rod-shaped, and spore-forming microbial entomopathogen. The silverleaf whitefly, Bemisia tabaci (Genn.) (Hemiptera: Aleyrodidae), is a worldwide important cryptic species causing serious economic damage to several vegetable crops including tomato. The aim of this study was to characterize and evaluate the bio-insecticidal effects of locally isolated Bacillus thuringiensis (Bt) against Galleria mellonella and Bemisia tabaci.
Results
Thirty-one isolates of Bt were obtained from 70 soil samples based on cultural characterization coupled with insecticidal crystal protein detection. All of the 31 bacterial isolates were pre-screened for their bio-insecticidal property against the greater wax moth, Galleria mellonella L. larvae. Only 20 (64.5%) isolates were virulent to G. mellonella with a percentage mortality that ranged from 2.50 to 95%. The groEL gene sequences of all 20 entomopathogenic indigenous isolates displayed 99–100% similarity with Bt isolates. Bioassay evaluation of 12 selected isolates against 3rd to 4th nymphal instars of B. tabaci with spore-crystal suspensions of 1 × 109 spores/ml caused 0 to 31.25%, 0 to 57%, and 0 to 82.5% percentage mortality within 48, 96, and 144 h accordingly. Among isolates of Bt, isolate AAUDS-16 had the highest virulence, followed by the isolate AAUES-69D with LC50 values of 9.67 × 106 spores/ml and 1.16 × 107 spores/ml, respectively. The virulent isolates were tested for their growth response to a different temperature range between 15 and 40 °C. All isolates showed a maximum growth rate around 30 °C.
Conclusions
Bt isolates of AAUES-69D and AAUDS-16 displayed high insecticidal potential against B. tabaci nymphs and G. mellonella larvae, and showed a maximum growth rate in a wide range of temperature. High virulence and temperature-tolerant isolates should be important candidates for diverse insecticidal toxin studies.
Similar content being viewed by others

Background
In Ethiopia, tomato (Lycopersicon esculentum Mill.) is an important food ingredient in a daily diet, a source of cash-generating to small-scale farmers, and delivers employment in both production and processing industries. Its production is mainly challenged by numerous abiotic and biotic factors (Sora 2018). In Ethiopia, insect pests (Ayalew et al. 2009) and associated diseases (Mandefro et al. 2009) are the most important biotic factors contributing to the lowest yield of tomatoes.
The silver leaf whitefly, Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae), is a global important cryptic species complex, resulting in serious economic damage on tomatoes because of its direct and indirect destruction (Shelby et al. 2020). The wide use of chemical pesticides poses a great threat to the environment and food safety (Sparks and Nauen 2015). The health hazards, environmental problems, and insect resistance associated with excessive use of chemicals necessitate for search-integrated pest management strategy (Aramideh et al. 2010).
Microbial biopesticides are used as a biocontrol agent against a wide variety of agricultural insect pests in many agroecosystems (Ruiu 2018). These microbes are natural enemies of insects, target-specific, and promising in reducing the use of hazardous chemical pesticides (Majeed et al. 2017). Among microbial entomopathogens, Bacillus thuringiensis (Bt) is the most important spore-forming microbial bio-control agent, which produces insecticidal crystal (Cry) and cytolytic (Cyt) protein (δ-endotoxins) encoded by cry and cyt genes (Soberon et al. 2018) and Vip protein (Chakroun et al. 2016). The insecticidal proteins are solubilized in the insect intestine after ingested, freeing protoxins that bind to specific receptors in the intestinal epithelium causing pore formation on the cellular membrane, which leads to an ionic imbalance and larval death (Soberon et al. 2018). The bacterium is the most successfully commercialized bio-control agent worldwide for its high specificity and associated environmental safety (Jurat-Fuentes and Crickmore 2017). However, excessive application of Bt has resulted in reduced efficacy as a result of resistance development with insect pests (Zago et al. 2014). This limited pathogenicity efficacy resolved with isolation and identification of propitious indigenous isolates. Hence, isolation and identification of new Bt strains from soil environment with privileged insecticidal properties is a vigorous research priority in many regions of the world (Reyaz et al. 2017).
Native isolates of Bt from soil environments caused up to 50 to 70% mortality of B. tabaci nymphs (Cabra and Hernandez Fernandez 2019) and significantly reduced G. mellonella within 48 h post-infection (Grizanova et al. 2019). In Ethiopia, Woldetenssay and Ashenafi (2008) studied the distribution of Bt on different agroecological soils of Ethiopia and found that 32% of the tested 503 soil samples harbored Bt. They showed that 44 (21%) isolates killed 50–100% of Anopheles arabiensis larvae within 48 h.
Therefore, there is a great potential to look for isolation and characterization of new Bt strains to find out strains with novel or high insecticidal activities. In this context, the present study was initiated to characterize the Bt isolates from soil samples of Menagesha forest and Rift-valley farmlands and evaluate their insecticidal activity against B. tabaci and G. mellonella under in vitro conditions.
Methods
Collection of soil samples
A total of seventy (70) soil samples were collected from Dalota, Kality, Erobgebaya, Koka, and Menagesha forest sites, Ethiopia. Samples were collected by scraping off the surface soil, and from each site, approximately 1.5 kg of soil samples were collected from 2 to 5-cm depth and placed in alcohol-sterilized polyethylene plastic bags. Collected soil samples were brought to the Applied Microbiology laboratory and stored at 4 °C for further processing.
Isolation of entomopathogenic bacterium Bt from soil
A modified version of the temperature selection method was used to isolate Bt from soil samples as described by Travers et al. (1987). Soil samples (10 g) were individually mixed in 100 ml sterile water and homogenized with an orbital shaker (200 rpm) for 4 h at room temperature. After complete homogenization, the samples were prepared to appropriate dilution from which samples were taken and heated at 80 °C for 10 min in a water bath equipped with a shaker to destroy non-spore formers and vegetative Bacillus cells. Then, 0.1 ml soil of aliquot samples was individually inoculated into nutrient agar and incubated at 28 °C for 48–72 h. Colonies were randomly picked and subcultured on nutrient agar and maintained for further investigation.
Cultural characterization
A total of 280 bacillus colonies were picked and characterized for their cultural characteristics such as colony’s color, shape, margin, elevation, and surface. Bacterial colonies showing typical Bt were preliminary identified based on cultural and morphological characteristics regarding Bergey’s Manual of Determinative Bacteriology (Halt et al. 1994).
Morphological characterization
Gram staining
Gram staining of isolates was carried out following the protocol of Provine and Gardner (1974). Thin bacterial smear was made on a clean glass slide, dried in the air, and heat-fixed. The smear was covered with crystal violet and kept for 1 min. The slide was washed, covered with Gram’s iodine, and let stand for 1 min. The slide was washed, decolorized with acetone, rocking the slide gently. The decolorization step was done very quickly. Immediately, the slide was washed with water, counterstained with safranine and let stand for 30 s, washed with water, blot dried, and examined under the microscope (× 100 magnifications).
Coomassie brilliant blue staining
The presence of parasporal bodies of isolates was detected, using Coomassie brilliant blue (CBB) staining, following the protocol of Rampersad et al. (2002). Selected isolates were inoculated into a sterile 50-ml conical flask containing nutrient broth and incubated in an orbital shaker (250 rpm) for 90 to 110 h at room temperature. Samples were smeared onto glass slides, followed by an air dry and heat fixing. Then, the slides were stained using 0.133% Coomassie blue stain in 50% acetic acid for 5 min. The dried slides were then observed under a phase-contrast microscope (× 100) to visualize the formation of parasporal bodies and spores.
Preparation of bacterial suspension
Isolates from nutrient agar slants were transferred into the fresh nutrient broth and incubated at 28 °C aerobically for complete sporulation and lysis of all vegetative cells. Completion of sporulation and lysis (spore-crystal formation) was detected by using a hemocytometer with the aid of a phase-contrast optical microscope (Soares-da-Silva et al. 2015). The spore-crystal mixture of each isolate was standardized at a concentration of 1 × 109 spores/ml for bio-assay.
Preliminary pathogenicity screening of bacterial isolates using G. mellonella under laboratory condition
Rearing of Galleria mellonella
Rearing of larvae of Galleria mellonella L. (Lepidoptera, Pyralidae) was undertaken according to Meyling (2007) at the Ambo Plant Protection Agricultural Research Center. Adult moths were kept in 500-ml flasks containing folded tissue paper to facilitate their mating and egg-laying potential. Eggs were laid on folded tissue paper, and each tissue paper was transferred from a flask into rearing plastic containers containing honey, wheat bran, and glycerol as dietary components for hatching larvae. The plastic containers were incubated at 20 °C for 4 weeks under darkness. The resulting third to fourth instar larvae were used for bio-assay evaluation.
Bio-assay evaluation
Insecticidal activities of isolates were preliminarily bio-assayed on the G. mellonella larvae, using the diet contamination technique (Navon et al. 1990). From each culture, 5 ml of spore-crystal suspension containing 1 × 109 spores/ml prepared and mixed with 15 g artificial diet containing honey, wheat bran, and glycerol. Then, 10 larvae of G. mellonella were transferred into an artificial diet containing a spore-crystal suspension mixture for feeding. The same number of larvae in an artificial diet without spore-crystal suspension was included as a control. All experiments were repeated 4 times. Periodic mortality was recorded every 48 h for 144 h.
Molecular characterization of entomopathogenic bacterial isolates
DNA extraction
The genomic DNA of isolates was extracted by the heat shock method using TE buffer (Ricieto et al. 2013). Isolates were grown on the nutrient agar at 30 °C for 15 h. Collected 10–20 mg bacterial isolates from the growing colony of approximately 1–2 mm in diameter was transferred to 200 μl of Tris–EDTA buffer (10 mM Tris; 1 mM EDTA; pH 8.0), using a sterile toothpick. The suspension was homogenized and incubated for 10 min in a boiling water bath (100 °C) for lysis of bacterial cells. Then, the suspension was centrifuged at 10,000g for 5 min to remove debris. The supernatant was transferred to a new microcentrifuge tube and stored at − 20 °C for PCR amplification reactions (polymerase chain reaction), and 5 μl of the supernatant was used as a template in the PCR reaction.
PCR amplification
The amplification of 533 bp groEL gene of Bt isolates was carried out, using B. cereus group-specific primer BalF (5′-TGCAACTGTATTAGCACAAGCT-3′) and BalR (5′TACCACGAAGTTTGTTCACTACT-3′) as described by Chang et al. (2003). Five microliters of the template was amplified in 25 μl of reaction mixture consisting of 10 mM Tris-HCl (pH-9), 50 mM KCl, 1.5 mM MgCl2, 200 μM of each dNTP, 1 U Taq DNA polymerase (Fermentas), and 10 pmol of respective primers. The PCRs were performed using a thermocycler (Mastercycler, Eppendorf, Germany) using BalF and BalR primers consist of 30 cycles of 94 °C for 45 s (denaturation), 55 °C for 45 s (annealing), and 72 °C for 45 s (extension). The amplified PCR products were analyzed in 1.5% agarose gel prepared in TAE buffer (40 mM Tris-acetate and 1 mM EDTA, pH 8.0) containing ethidium bromide (0.3 μg/ml). Electrophoresis was carried out at 7 V/cm for 1 h and 30 min in a submarine gel electrophoresis system (Bangalore Genei, India), and the gel was photographed by UV gel documentation system (Alpha Innotech Corporation, USA). The PCR product was purified by using the QIAquick PCR purification kit according to the manufacturer’s instructions (QIAGEN, Germany) and sequenced. Finally, sequenced data were submitted to NCBI (National Center for Biotechnological Information) and compared to publish sequence of NCBI database for the identification of bacterial isolates at the species level.
In vitro bioassay evaluation of bacterial isolates against B. tabaci
Rearing of whiteflies
Whitefly adults were collected from Koka in the tomato field using a manual aspirator. The area is characterized by a mean minimum and maximum temperature of 12.14 °C and 27.39 °C, respectively Ethiopian sugar development agency (ESDA 2010). After collection, insects were reared on young tomatoes planted in pots in whitefly-proof cages (Safavi and Bakhshaei 2017). Tomato seedlings with 2 or 4 leaves were placed into a cage and allowed to be infested by adult whiteflies (sex ratio ≈ 1:1). Adults were allowed to oviposit for 24 h on leaves. Then, adults were removed, and plants with whitefly eggs were transferred to other insect-free cages. Whitefly eggs were allowed to develop into the nymphal stage, and 3rd and 4th nymphal instars were applied in bioassays
Single-concentration bioassay
Virulent isolates were evaluated against the nymph of B. tabaci under laboratory conditions with the leaf disc method (Ateyyat et al. 2009). The leaf discs with 3rd to 4th nymphal instars were cut from plants and immersed in 1 × 109 spores/ml concentration for 10 s and placed upside down onto 0.2% plain water agar medium in Petri plates. Four replicates (Petri dishes) were set per treatment. Nymphs were considered dead if body color changed from yellowish to dark brown or if the body appeared dry. Periodic mortality was recorded every 48 h for 144 h. Isolates with mortality greater than 50% were selected to estimate the median lethal concentration (LC50).
Multiple-concentration bioassays
The multiple-concentration bioassays were evaluated to estimate LC50, following the leaf disc method as before (Ateyyat et al. 2009). For this study, four (4) highly virulent isolates were selected and evaluated. Leaf discs with 3rd to 4th nymphal instars were immersed in the 5 concentration ranges from 1 × 105 to 1 × 109 spores/ml of each isolate. For each concentration, 4 repetitions were performed, and periodic mortality was recorded every 48 h for 144 h.
Effect of temperature on the growth of isolates
The growth rate response of isolates to different temperatures was evaluated as described by Hamedo (2016) with a little modification. Isolates were inoculated on nutrient broth and incubated for 24 h. One milliliter of each broth culture (cell suspensions) was inoculated into 50 ml sterilized nutrient broth in a 250-ml Erlenmeyer flask and incubated at 15, 20, 25, 30, 35, and 40 °C. The vegetative growth phase of isolates was determined by the growth curve experiment. The maximum growth rate of isolates was measured by taking optical density reading at 600 nm in 6-h intervals starting from 0 h, using a spectrophotometer. Optical density reading and colony-forming units (CFU) were used to plot the regression line, and the maximal growth rate of each isolates in response to different temperatures was calculated.
Data analysis
Mortality data were corrected using Abbot’s formula (Abbott, 1925), arcsine transformed, and subjected to the ANOVA procedure of SPSS version 20.0. Means were separated using Tukey’s honestly significant difference (HSD) at 5% significance level for screening experiments against G. mellonella and B. tabaci, respectively. The LC50 and LC90 values of isolate were determined by using probit analysis (Finney, 1971).
Results
Cultural characterization of bacterial isolates
A total of 280 Bacillus isolates were detected using heat treatment methods from 70 soil samples collected from Menagesha forest and various tomato-cultivated land of the central rift valley of Ethiopia. The heat treatment method eliminates non-spore formers and vegetative Bacillus cells in the soil samples. All of the 280 Bacillus-like colonies were culturally characterized by examining the different characters including colony color, elevation, margin, shape, and colony surface. The cultural study showed that a total of 40 isolates were carefully chosen based on typical colonial characteristics of Bt showing white to off white colony color, regular to irregular colony margin, circular colony shape, wrinkled to wavy colony surface, and opaque and flat to slightly raised colony elevation. The result showed that the different isolates showed variations among one another in their cultural characteristics.
Morphological characterization
Gram staining
Culturally characterized isolates were subjected to gram staining to approve the bacterial colonies to be Gram-positive. About 40 isolates selected based on colony morphology were evaluated for gram staining. Isolates that were rod-shaped and blue indicate Gram-positive isolates. Based on this result, all isolates were found to be Gram-positive, rod-shaped, and spore formers. The isolate absorbed the crystal violet and did not decolorize with the decolorized agent indicating that the cell wall of the organism was composed of peptidoglycan.
Coomassie brilliant blue staining
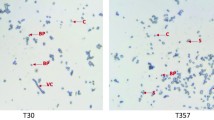
After Gram staining, isolates were subjected to Coomassie brilliant blue staining as the best method to detect Bt isolates from the Bacillus cereus group. The phase-contrast microscope analysis revealed that from the total of 40 isolates stained with Coomassie brilliant blue staining, 31 isolates were found to have parasporal inclusion (insecticidal proteins) and classified as Bt (Table 1). In contrast, the rest of the 9 isolates were not grouped under Bt due to they failed to have parasporal inclusion. Isolates did not have any crystal bodies might be grouped under Bacillus cereus or other B. cereus group. The parasporal crystalline inclusions of Bt isolates were stained as dark blue (Fig. 1). The crystal morphology results of Bt isolates were indicated 8 (25.8%) bipyramidal, 5 (16.13%) pyramidal, 7 (25%) spherical, 5 (16.13%) oval, 4 (12.9%) rod, and 2 (6.5%) amorphous crystal shapes. The crystal protein morphology variation of isolates might be due to the genetic variation of entomopathogenic bacterial isolates.

Microscopic images of Coomassie brilliant blue stained vegetative cells, spores and crystal proteins of B. thuringiensis isolates AAUMS-16 (a) and AAUKoS-12 (b)
The Bt index was calculated as the number of identified Bt isolates divided by the total number of Bacillus-like colonies examined. Based on the result, the Kality sampling site showed the highest Bt index with 0.17, followed by Koka with 0.13 Bt index. Menagesha forest sampling site showed the lowest Bt index with 0.09, and an average Bt index of the total sampling sites was 0.11 (Table 2).
Preliminary pathogenicity screening of isolates using G. mellonella larvae under laboratory condition
Isolates found to have insecticidal proteins were selected and primarily assessed for their toxicity against the G. mellonella larvae. The preliminary toxicity screening of 31 isolates with concentrations of spore-crystal suspensions (1 × 109 spores/ml) exhibited a wide range of toxicity toward 3rd instar larvae of G. mellonella. From the total of 31 isolates evaluated, 20 (64.5%) isolates showed insecticidal effect, and the mortality of larvae ranged from 2.50 to 62.50%, 10 to 75%, and 20 to 95% within, 48, 96, and 144 h, respectively (Table 3).
The mortality percentage of larvae after 48 h of treatment application ranged between 2.50 and 62.50%. The maximum percentage of mortality was recorded (62.5%) with isolate AAUES-20, whereas the minimum one (2.5%) was recorded by isolate AAUDS-18. Isolate AAUES-20 and AAUMS-11 showed the highest (75%) and lowest (10%) percentages larval mortality after 96 h of treatment accordingly. After 144 h of application of spore-crystal mixture, isolate AAUES-36A caused the maximum mortality (95%), whereas isolate AAUMS-11 triggered the least mortality (20%). There was a discrepancy in toxicity against G. mellonella among tested isolates at P < 0.01. The result indicated that the percentage of larval mortality increases when larval exposure time of spore-crystal mixture increases (Table 3).
Molecular characterization of bacterial isolates
A total of 20 isolates of Bt screened from cultural characterization, crystal inclusion analysis, and preliminary bioassay evaluation were selected for molecular characterization study. The entomopathogenic bacterial isolates were identified based on groEL gene detection with the heat-shock method. All isolates (100%) produced 533 bp amplified PCR products of groEL genes by using B. cereus group-specific primers, BalF, and BalR (Fig. 2 a and b). The sequences of the groEL gene of all indigenous isolates showed 99–100% BLAST search similarity with Bt isolates excavated in GeneBank.

Agarose gel electrophoresis of amplified PCR products of groEL genes using Bacillus cereus group specific primer of BalF and BalR from twenty selected isolates of Bacillus thuringiensis from Ethiopia a and b. M, 100 bp ladder; 0, blank; B1, AAUDS-11, B2, AAUKS-15, B3, AAUDS-16, B4, AAUKS-9, B5, AAUKS-92, B6, AAUKS-16, B7, AAU KS-13, B8, AAUES-6, B9, AAUMS-54, B10, AAUMS-11, B11, AAUDS-18, B12, AAUES-8, B13, AAUES-36A, B14, AAUES-69D a and M, 100 bp ladder; 0, blank; B15, AAUES-20; B16, AAUMS-23; B17, AAUMS-16; B18, AAUKOS-12; B19, AAUKS-19; B20, AAUDS-12 b
The phylogenetic tree of indigenous isolates was constructed by neighbor-joining methods using the mega software (mega4.1) with 1000 bootstrap replications. The neighbor-joining examination showed that isolates of Bt from different soil sample sites of Ethiopia were closely related. The bootstrap displayed at the nodal branches and their corresponding accession numbers of isolates were indicated in aside (Fig. 3). Phylogenetic analysis showed that all isolates were positioned as part of the species of Bt, and their comparable accession number placed in digression. Isolates of AAUDS-11, AAUKS-15, AAUDS-16, AAUKS-9, AAUKS-92, AAUKS-16, AAUKS-13, AAUES-6, and AAUMS-54 proved to be very close to each other, and they were all clustered in the Bt clade. Besides strains of AAUMS-11, AAUDS-18, AAUES-8, and AAUES-36A were in the same clade even if the genetic distance of these strains in respect to Bt AY112850 might be suggested a new variety or subtype.

Phylogenetic tree of groEL gene sequences of 20 Bacillus thuringiensis isolates from Ethiopia related to the sequence of bacterial isolates found in Gene bank
Bioassay evaluation of Bt against B. tabaci under in vitro conditions
Isolates showed greater than 80% mortality on G. mellonella larvae at 144 h were selected and further screened against whitefly (B. tabaci). Bioassay evaluation of twelve (12) isolates of Bt against nymphs of B. tabaci with a concentration of spore-crystal suspension (1 × 109 spores/ml) caused 0–31.25%, 0–57.5%, and 0–82.5% percentage mortality within 48, 96, and 144 h, respectively (Table 4).
In the 48-h treatment against nymph of B. tabaci, the highest mortality was achieved by the isolate AAUES-69D (31.25%), followed by isolate AAUES-20 (27.50%), whereas isolates AAUKS-13 and AAUMS-54 caused the lowest mortality (0%). Isolates of AAUDS-16 and AAUMS-54 caused the maximum (57.5%) and the minimum mortality (0%) against B. tabaci nymph after 96-h treatment with spore-crystal mixture respectively. The maximum percentage mortality (82.5%) was recorded by the isolate AAUES-69D, and the minimum mortality (0%) was reached by the isolate AAUMS-54 after 144 h of application. The percentage mortality of whitefly infected with Bt isolates showed significant variation (P < 0.01).
Isolates with mortality greater than 62% against whitefly nymphs were selected to estimate the LC50 values. Multiple concentration (1 × 105–1 × 109) response evaluations of the 4 most promising isolates of Bt were performed on B. tabaci nymphs under laboratory conditions for 144 h. The LC50 values of isolates were observed in the range of 9.67 × 106 to 1.21 × 108 spores/ml (Table 5). The maximum LC50 value (1.21 × 108 spores/ml) was recorded by isolate AAUMS-23, followed by isolate AAUES-36A (3.20 × 107 spores/ml), whereas the minimum LC50 value of 9.67 × 106 spores/ml was indicated by isolate AAUDS-16. The percentage mortality of B. tabaci nymphs was recorded after144 h of post-application of Bt isolates in multiple-concentration assessment ranged from 1 × 105 to 1 × 109 spore/ml. The highest mortality was achieved by isolate AAUES-69D at 1 × 105, 1 × 108, and 1 × 109 spore/ml, while at the concentrations of 1 × 106 and 1 × 107 spores/ml, the maximum mortality of B. tabaci was displayed by isolate AAUDS-16. The percentage mortality of B. tabaci was observed to increase with a consequent increase in the concentrations of toxin-spore suspension of selected isolates of Bt (Fig. 4). All the treatments exhibited significant differences in percent mortality of B. tabaci from different treatments.

Multiple concentrations caused percentage mortality response of spore-crystal Bacillus thuringiensis against Bemisia tabaci nymphs after 144 h of application
Effect of temperature on the growth of Bt isolates
The virulent isolates were tested for their growth response to various temperatures (Fig. 5). The mean growth rate of isolates ranged from 0.08 to 0.83, 0.97 to 1.57, 1.77 to 2.73, 2.56 to 3.06, 2.07 to 2.73, and 0.83 to 1.70 generations/h at 15, 20, 25, 30, 35, and 40 °C accordingly. Even though isolates were grown in all tested temperature range from 15 to 40 °C, all of the isolates showed the maximum growth rate (generations/h) around 30 °C. The height growth rate was recorded by an isolate of AAUES-69D (3.06 generations/h), whereas the lowest growth rate was recorded with isolates of AAUMS-23 (2.56 generations/h) at 30 °C.

The effects of various temperature on the growth rate (generations/h) of selected Bacillus thuringiensis isolates
Average growth rate of isolates in response to all tested temperatures ranged between 1.54 and 2.07 growth rates (generation/h). An isolate of AAUES-69D exhibited the maximum growth rate (2.07 generation/h), followed by AAUDS-16 (1.86 generation/h), whereas the minimum growth rate was recorded with isolate AAUMS-23 (1.54 generation/h). Generally, isolates of AAUES-69D and AAUDS-16 displayed high insecticidal potential against nymphs of B. tabaci with minimum LC50 values of 1.16 × 107 and 9.67 × 106 spores/ml, respectively; also, these isolates showed a maximum growth rate of 2.07 and 1.86 generation/h, respectively, in a response to a wide range of temperature accordingly.
Discussion
In this particular study, cultural, morphological, molecular, and bio-insecticidal activities were used to describe Bt isolates from soil samples collected from Menagesha forest and various tomato-cultivated land of the central rift valley of Ethiopia. A total of 280 Bacillus isolates were detected from 70 soil samples using heat treatment methods. Likewise, Woldetenssay and Ashenafi (2008) isolated a total of 213 Bacillus-like colonies from 503 soil samples collected from the different agro-ecological zone of Ethiopia. Another study showed that about 89 Bacillus-like colonies were isolated from 207 soil samples of different sites (Lone et al. 2017). Captivatingly, soil environment could be inhabited by diverse microbial community and preferred as the main source for the isolation of Bacillus species in general and Bt in particular (Soares-da-Silva et al. 2015). The cultural characterization parameter preliminarily identified 40 isolates with typical colonial characteristics of Bt. In this result, the different isolates showed variations among one another in their colony color, margin, surface, and elevation. The different colony morphology of Bt was described as white to off-white colony color with irregular colony margin and flat to slightly raised colony elevation (Chai et al. 2016 and Padole et al. 2017).
Culturally characterized isolates were further subjected to morphological studies based on Gram staining and Coomassie brilliant blue staining as the suited parameters to distinguish Bt isolates from B. cereus group. All isolates were Gram-positive, spore-former, and rod-shaped bacteria. A similar result showed that Bt isolates were spore-former, rod-shaped, and gram-positive soil-inhabiting bacteria (Kassogué et al. 2015). The Coomassie brilliant blue staining analysis showed that from the total of 40 isolates, only 31 isolates were comprised of parasporal inclusion and classified as Bt whereas 9 isolates failed to have parasporal inclusion. Studies reported that crystal protein inclusions were the unique characteristic of Bt that distinguish Bt from other Bacillus species (Woldetenssay et al. 2009). In this crystal morphology study, Bt isolates achieved 25.8% bipyramidal, 16.13% pyramidal, 25% spherical, 16.13% oval, 12.9% rod, and 6.5% amorphous crystal shapes. The crystal morphology variation was supported by Mukhija and Khanna (2018) who studied the crystal morphology of 140 Bt isolates and reported 52 (37.1%) bipyramidal, 31 (22.1%) pyramidal, 21 (15%) cuboidal, 18 (12.9%) round, and 18 (12.9%) amorphous crystal shapes. The differences in the crystal protein morphology distribution of isolates could be due to genetic variation caused by the difference in the environmental conditions.
The preliminary pathogenicity screening of entomopathogenic bacterial isolates was carried out by using G. mellonella larvae. From the total of 31 tested isolates, 20 (64.5%) isolates were virulent against G. mellonella with the percentage mortality of larvae ranged from 2.50 to 62.50%, 10 to 75%, and 20 to 95% within 48, 96, and 144 h post-application, respectively. There was a difference in toxicity against G. mellonella among tested isolates at P < 0.01. Bio-insecticidal evaluation of Bt revealed a significant reduction against G. mellonella larvae within 48 h post-infection (Grizanova et al. 2019). Research findings supported that G. mellonella larva was used as the best model host to study the interactions between insect pests and Bt (Dubovskiy et al. 2016). Moreover, the insecticidal potential evaluation of Bt isolates resulted in significant mortality against G. mellonella (Basedow et al. 2012) under laboratory and field conditions.
Molecular characterization of Bt is a vigorous technique than morphological, cultural, and physiological classification methods due to microbiological indistinguishable features of Bt isolates from other B. cereus groups (Abo-Bakr et al., 2020). As a result, 20 isolates screened from preliminary bioassay evaluation were selected for molecular characterization study. These entomopathogenic bacterial isolates were identified based on the groEL gene, and all isolates (100%) produced 533 bp amplified PCR products of groEL genes by using B. cereus group-specific primers of BalF and BalR. By the same token, the B. cereus group-specific primers (BalF and BalR) displayed 533 bp amplified products of groEL genes in detection and differentiation of B. cereus group isolates (Chang et al. 2003). According to Wei et al. (2018), Bt and other B. cereus group proficiently identified and evaluated with groEL gene sequences and its primers showed 56 (97%) amplified from 58 B. cereus isolates whereas, among 50 isolates, all (100%) isolates indicated amplified PCR products in Bt. As well, the groEL gene was a potential phylogenetic marker to adequately differentiate the species of B. cereus group compared to 16S ribosomal DNA gene sequence analysis (Chang et al. 2003).
The neighbor-joining examination showed that isolates of Bt from different soil sample sites of Ethiopia were closely related. The phylogenetic tree of indigenous isolates was constructed by neighbor-joining methods using the mega software. Phylogenetic analysis verified all bacterial isolates were positioned as part of the species of Bt as described by El-kersh et al. (2012).
Bioassay evaluation of 12 selected isolates of Bt against 3rd to 4th nymphal instars of B. tabaci with a concentration of spore-crystal suspensions (1 × 109 spores/ml) triggered 0 to 31.25%, 0 to 57.5%, and 0 to 82.5% percentage mortality within 48, 96, and 144 h, respectively. The highest mortality was achieved by isolate AAUES-69D (31.25%) after 48 h, isolate AAUDS-16 (57.5%) after 96 h, and AAUES-69D (82.5%) after 144 h post-applications. This result in lined with the study stated that 34.84 to 51.21% mortality of B. tabaci nymphs after 48 h treated with different isolates of Bt (Ruiz-Sánchez et al. 2019). Salazar-Magallon et al. (2015) reported more than 92% 3rd to 4th nymphal instars of whitefly mortality after treated with culture broth containing spore-crystal complex. In Ethiopia, from the total of 130 isolates of Bt, 39 (30%) isolates revealed insecticidal properties against Aedes africanus (Ashenafi et al., 2001). The different isolates of Bt proved to successfully control insect pests by producing a wide range of insecticidal parasporal crystal proteins (Jurat-Fuentes and Crickmore, 2017).
Multiple concentration (1 × 105–1 × 109 spores/ml) response evaluations of most promising isolates of Bt were performed on B. tabaci nymphs under laboratory conditions for 144 h. The LC50 values of isolates were observed in the range of 9.67 × 106 to 3.32 × 107 spores/ml. Isolates of Bt AAUDS-16 reached the highest virulence, followed by isolate AAUES-69D with LC50 value of 9.67 × 106 and 1.16 × 107 spores/ml accordingly. The percentage mortality of B. tabaci was observed to increase with a subsequent increase in the concentrations of toxin-spore suspension of selected isolates of Bt. Correspondingly, Sivaji and Girija (2017) also described the increase in percent mortality of whitefly pests along with the increase in the toxin-spore concentration of Bt isolates. Significant differences were observed in the percentage mortality of the tested B. tabaci by different treatments. In the same way, variations in the effectiveness among different concentrations of Bt isolates were described (Patel et al. 2018).
The virulent isolates were tested for their growth response to various temperature ranges from 15 to 40 °C, and all of the isolates showed a maximum growth rate around 30 °C. Isolate AAUES-69D exhibited the maximum growth rate of 2.07 generation/h, followed by AAUDS-16 (1.86 generation/h). Woldetenssay and Ashenafi (2009) studied the effects of temperature on the growth rate of isolates from different agroecological zones of Ethiopia, and the result indicated that isolates of Bt showed maximal growth rate at 30 °C even if all isolates grew well at a temperature raged from 12 to 39 °C. Besides, the growth of Bt isolates was also proved at a different temperature range (25–50 °C), and the maximum growth rate of isolates showed between 30 and 35 °C (Abo-Bakr et al. 2020).
Generally, isolates of AAUES-69D and AAUDS-16 displayed high insecticidal potential against nymphs of B. tabaci with minimum LC50 values of 1.16 × 107 and 9.67 × 106 spores/ml, and showed a maximum growth rate of 2.07 and 1.86 generation/h in a response to a wide range of temperature accordingly. These remarkable features of isolates are very important to screen, and they are shown to be competitive for future bio-pesticide applications for sustainable crop production under field conditions with a variable temperature range.
Conclusions
The present study showed the cultural, morphological, and bio-insecticidal diversity of Bt isolates detected from cultivated and forest soil samples. The insecticidal properties of Bt were evaluated against G. mellonella and B. tabaci nymphs. The percentage mortality effects of isolates showed a significant difference in both G. mellonella larvae and B. tabaci nymphs. Isolates of AAUDS-16 and AAUES-69D achieved the lowest LC50 values and exhibited an average maximum growth rate at different temperature ranges. As a result, these isolates had the potential for biological control of whitefly and should be important candidates for different insecticidal toxin studies and use as gene sources for the construction of transgenic tomato plants.
Availability of data and materials
All data and material are stated in the manuscript.
Abbreviations
- Bt:
-
Bacillus thuringiensis
- AAU:
-
Addis Abab University
- CBB:
-
Coomassie brilliant blue
- PCR:
-
Polymerase chain reaction
- TAE:
-
Tris-acetate-EDTA
- NCBI:
-
National Center for Biotechnological Information
- EDTA:
-
Ethylenediaminetetraacetic acid
- UV:
-
Ultraviolet
- LC50 :
-
Median lethal concentration
- SE:
-
Standard error
- CL:
-
Confidence limit
- χ2 :
-
Chi square
References
Abbott WS (1925) A method of computing the effectiveness of an insecticide. J econ Entomol 18:265–267
Abo-Bakr A, Fahmy E M, Badawy F, Abd El-latif, A O, and Moussa S (2020) Isolation and characterization of the local entomopathogenic bacterium, Bacillus thuringiensis isolates from different Egyptian soils. Egypt J Biol Pest Control (2020):30:54.
Aramideh S, Saferalizadeh MH, Pourmirza AA, Bari MR, Keshavarzi M, Mohseniazar M (2010) Characterization and pathogenic evaluation of Bacillus thuringiensis isolated from West Azerbaijan province-Iran. Afr J Microbiol Res 4(12):1224–1229
Ashenafi A, Gebre-Michael T, Balcha F, and Seyoum A (2001) Isolation and characterization of B. thuringiensis from different agro-ecological zones of Ethiopia and assessment of the insecticidal potential of the isolates. Annual report 2000/2001, Institute of Pathobiology, Addis Ababa University.
Ateyyat MA, Shatnawi M, Al-Mazra’awi MS (2009) Culturable whitefly associated bacteria and their potential as biological control agents. Jordan J Biol Sci 2(3):139–144
Ayalew G, Mulatu B, Negeri M, Merene Y, Sitotaw L, and Tefera, T (2009) Review of research on insect and mite pests of vegetable crops in Ethiopia. In: Proceeding of Plant Protection Society of Ethiopia (PPSE): Increasing Crop Production through Improved Plant Protection volume II, pp. 47–67, (Abreham, T., ed.). Addis Ababa, Ethiopia.
Basedow TH, Elshafie HAF, Abo-Elsaad MM, Al-Ajlan AM (2012) Evaluation of Bacillus thuringiensis and neem for controlling the larvae of the greater wax moth, Galleria mellonella (Lepidoptera). Int J Agricult Biol 14(4):629–632
Cabra ET, Hernandez Fernandez JA (2019) Larvicidal activity of Bacillus thuringiensis Colombian native strains against Bemisia tabaci (Hemiptera: Aleyrodidae). J Plant Protect Res:503–511
Chai P F, Rathinam X, Ghazali1 A H, and Subramaniam S (2016) Characterization of a native Bacillus thuringiensis isolates from Malaysia that produces exosporium enclosed parasporal inclusion. Emirates J Food Agricult 28(9): 653-659.
Chakroun M, Banyuls N, Bel Y, Escriche B, Ferré J (2016) Bacterial vegetative insecticidal proteins (Vip) from entomopathogenic bacteria. Microbiol Mol Biol Rev 80:329–350
Chang YH, Shangkuan YH, Lin HC, Liu HW (2003) PCR assay of the groEL gene for detection and differentiation of Bacillus cereus group cells. Appl Environ Microbiol 69:4502–4510
Dubovskiy IM, Grizanova EV, Whitten MMA, Mukherjee K, Greig C, Alikina T, Kabilov M, Vilcinskas A, Glupov VV, Butt TM (2016) Immuno-physiological adaptations confer wax moth Galleria mellonella resistance to Bacillus thuringiensis. Virulence 7 https://doi.org/10.1080/21505594.2016.1164367
El-kersh TA, Al-sheikh YA, Al-akeel RA, Alsayed AA (2012) Isolation and characterization of native Bacillus thuringiensis isolate from Saudi Arabia. Afr J Biotechnol 11(8):1924–1938
ESDA (Ethiopian Sugar Development Agency) (2010) Research directorate researchers’ orientation manual. Wonji, Ethiopia
Finney DJ (1971) Probit analysis. Cambridge University Press, Cambridge, pp 3–197
Grizanova EV, Krytsyna TI, Surcova VS, Dubovskiy MI (2019) The role of midgut nonspecific esterase in the susceptibility of Galleria mellonella larvae to Bacillus thuringiensis. J Invert Pathol 166:107208
Halt JG, Krig Peter NR, Sneath HA, Staley JT, William ST (1994) Burgey’s manual of determinative bacteriology, 9th edn. William and Wilkins, Baltimore, p 559
Hamedo HA (2016) Identification of Bacillus thuringiensis isolated from different sources by Biologic Gen III system and scanning electron microscopy. Catrina 15(1):51–57
Jurat-Fuentes JL, Crickmore N (2017) Specificity determinants for Cry insecticidal proteins: insights from their mode of action. J Invert Pathol 142:5–10
Kassogué A, Maïga K, Traoré D, Dicko AM, Fané R, Guissou T, Faradji FA, Valicente FH, Hamadoun A, Babana AH (2015) Isolation and characterization of Bacillus thuringiensis strain indigenous to agricultural soils of Mali. Afr J Agricult Res 10(28):2748–2755
Lone SA, Malik A, Padaria JC (2017) Selection and characterization of Bacillus thuringiensis strain from northwestern Himalayas toxic against Helicoverpa armigera. Microbiol Open 6:1–11
Majeed MZ, Fiaz M, Ma CS, Afzal M (2017) Entomopathogenicity of three muscardine fungi, Beauveria bassiana, Isaria fumosorosea and Metarhizium anisopliae, against the Asian citrus psyllid, Diaphorina citri Kuwayama (Hemiptera: Psyllidae). Egypt J Biol Pest Control 27(2):211–215
Mandefro M, Ahmed E, Yesuf M, Lencho A, Hiskias Y, Shehabu M, Abebe F, Hussien T, and Abraham A (2009) Review of vegetable disease research in Ethiopia. In: Proceeding of Plant Protection Society of Ethiopia (PPSE): Increasing Crop Production through Improved Plant Protection volume II, pp. 203–230, (Abreham, T., ed.). Addis Ababa, Ethiopia.
Meyling N V (2007) Methods of isolation of entomopathogenic fungi from the soil environment. A laboratory manual, January 2007. (http:// orgprints.org/11200).
Mukhija B, Khanna V (2018) Isolation, characterization and crystal morphology study of Bacillus thuringiensis isolates from soils of Punjab. J Pure Appl Microbiol 12(1):189–193
Navon A, Klein M, Braun S (1990) Bacillus thuringiensis potency bioassays against Heliothis armigera, Earias insulana and Spodoptera littoralis larvae based on standardized diets. J Invert Pathol 55:387–393
Padole DA, Moharil MP, Ingle KP, Shyamkant Munje S (2017) Isolation and characterization of native isolates of Bacillus thuringiensis from Vidarbha. Int J Curr Microbiol App Sci 6(1):798–806
Patel AS, Shelatl HN, Patel HK (2018) Isolation and insecticidal potential of native Bacillus thuringiensis against Helicoverpa armigera and Spodoptera litura Int. J Curr Microbiol App Sci 7(2):1330–1339
Provine H, Gardner P (1974) The Gram’s stained smear and its interpretation. Hosp Pract 9:85–91
Rampersad J, Ayub K, Ammons D (2002) Usefulness of staining parasporal bodies when screening for Bacillus thuringiensis. J Inverteb Pathol 79(3):203–204
Reyaz A, Gunapriya L, Arulselvi PI (2017) Molecular characterization of indigenous Bacillus thuringiensis strains isolated from Kashmir valley. Biotech 7:143
Ricieto APS, Fazion FAP, Carvalho Filho CD, Vilas-Boas LA, Vilas-Bôas GT (2013) Effect of vegetation on the presence and genetic diversity of Bacillus thuringiensis in soil. Can J Microbiol 59:28–33
Ruiu L (2018) Microbial biopesticides in agroecosystems. Agronomy 8:235
Ruiz-Sánchez E, García-Ramírez A, Reyes-Ramírez A, Gamboa-Angulo M, José María Tun-Suárez J, Cristóbal-Alejo J (2019) Biological activity of Bacillus thuringiensis culture supernatant on Bemisia tabaci and its parasitoid Eretmocerus eremicus. Trop Subtrop Agroecosyst 22:213–218
Safavi SA, Bakhshaei M (2017) Biological parameters of Trialeurodes vaporariorum (Hemiptera: Aleyrodidae) exposed to lethal and sub-lethal concentrations of Calypso®. J Crop Prot 6(3):341–351
Salazar-Magallon JA, Hernandez-Velazquez VM, Alvear-Garcia A, Arenas-Sosa I, Pena-Chora G (2015) Evaluation of industrial by-products for the production of Bacillus thuringiensis strain GP139 and the pathogenicity, when applied to Bemisia tabaci nymphs. Bull Insectol 68(1):103–109
Shelby EA, Moss JB, Andreason SA, Simmons AM, Moore AJ, Moore PJ (2020) Debugging: strategies and considerations for efficient RNAi-mediated control of the whitefly Bemisia tabaci. Insects 11:723
Sivaji M, Girija D (2017) Evaluation of bio-insecticidal property of Bacillus thuringiensis strains isolated from Western Ghats soil. M R J I 21(2):1–12
Soares-da-Silva J, Pinheiro VCS, Litaiff-Abreu E, Polanczyk RA, Tadei WP (2015) Isolation of Bacillus thuringiensis from the state of Amazonas, in Brazil, and screening against Aedes aegypti (Diptera, Culicidae). Rev Bras Entomol 59:1–6
Soberon M, Monnerat R, and Bravo A (2018) Mode of action of cry toxins from Bacillus thuringiensis and resistance mechanisms. In: P. Gopalakrishnakone, B. Stiles, A. Alape-Girón, J.D. Dubreuil And M. Mandal, eds. Microbial toxins: toxinology. Dordrecht: Springer, pp. 1-13.
Sora S (2018) Review on productivity of released tomato (Solanum Lycopersicum Mill.) varieties in different parts of Ethiopia. J Hortic Sci For 1:102
Sparks TC, Nauen R (2015) IRAC: mode of action classification and insecticide resistance management. Pestic Biochem Phys 121:122–128
Travers RS, Martin AW, Reichelderfer CF (1987) Selective process for efficient isolation of soil Bacillus species. Appl Environ Microbiol 53:1263–1266
Wei S, Chelliah R, Park B, Park J, Forghani F, Park Y, Cho MS, Park DS, Oh DH (2018) Molecular discrimination of Bacillus cereus group species in foods (lettuce, spinach, and kimbap) using quantitative real-time PCR targeting groEL and gyrB. Microbial Pathogenesis 115:312–320
Woldetenssay Z, Ashenafi M (2008) Frequency and distribution of Bacillus thuringiensis from Ethiopian soils. Ethiop J Biol Sci 7(1):55–67
Woldetenssay Z, Ashenafi M, Eiler A, Bertilson S (2009) Isolation and characterization of Bacillus thuringiensis from soils in contrasting agroecological zones of Ethiopia. SINET: Ethiop J Sci 32(2):117–128
Zago HB, Siqueira HÁA, Pereira EJG, Picanço MC, Barros R (2014) Resistance and behavioural response of Plutella xylostella (Lepidoptera: Plutellidae) populations to Bacillus thuringiensis formulations. Pest Manag Sci 70:488–495
Acknowledgements
The authors gratefully thank the Healthy Seedling Project granted by both the Ethiopian Biotechnology Institute and the Regional Project supported by the Austrian Development Agency (ADA) for financial support in this study.
Funding
Culture media, chemicals, reagents, substrates, stationary materials, and laboratory and field equipment were supplied by the Ethiopian Biotechnology Institute and the Regional Project.
Author information
Authors and Affiliations
Contributions
AG planned the study, carried out the experiment, analyzed data, and wrote the manuscript. YC and FA supervised the overall activities in the study and editing manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable
Consent for publication
Not applicable
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Gebremariam, A., Chekol, Y. & Assefa, F. Isolation, characterization, and bio-insecticidal efficiency of Ethiopian isolates of Bacillus thuringiensis against Galleria mellonella L. (Lepidoptera: Pyralidae) and tomato whitefly, Bemisia tabaci (Genn.) (Hemiptera: Aleyrodidae). Egypt J Biol Pest Control 31, 28 (2021). https://doi.org/10.1186/s41938-021-00375-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s41938-021-00375-9