
Abstract
The aim of the present study is to investigate the effect of three entomopathogenic fungi (EPF) (Beauveria bassiana, Cladosporium cladosporioides, and Verticillium alfalfae) on the aphid species, Metopolophium dirhodum (Walker) (Hemiptera: Aphididae). The selected EPF were isolated from the agricultural soil of the National Institute of Plant Protection (INPV) in Constantine, Algeria, and were tested against the aphid insects that were collected from the same area. The aphid species M. dirhodum were exposed to each fungal spore suspensions 107 conidia/ml for 10 s. Percent mortality was recorded at 1, 3, 5, and 7 days post treatment. Percentage mortalities, 7 days post treatment, were 95.83, 63.98, and 51.83% by B. bassiana, C. cladosporioides, and V. alfalfae, respectively. The higher protease activities were observed for isolate V. alfalfae with 95 U/ml, followed by B. bassiana with 38.26 U/ml, and finally C. cladosporioides with 35, 65 U/ml. The results presented in this study revealed that there was no relation between high alkaline protease activities and high virulence isolates.
Similar content being viewed by others

Background
The aphids are the most harmful insects that affect wheat plants. Diuraphis noxia (Mordvilko), Sitobion avenea (F.), Schizaphis graminum (Rondani), Rhopalosiphum padi (L.), Metopolophium dirhodum (Walker), and Sipha maydis (Passerini) are the most inimical insect pests that depreciate substantially cereal cultures (Blackman and Eastop 2000).
M. dirhodum is the most important species that could destroy grains all over the world (Dixon 1987). It chooses its host during the cold season and then emigrates toward drought cereals in summer and is able to transmit a Luteovirus and provides a wide resistance against exterminators (Carter et al. 1980). The biocontrol methods, using entomopathoganic fungi (EPF), are among the most effective ones (Wells et al. 2000).
The efficiency of these fungi lies in the fact that they can fight against exterminators by invading insect crusts. They usually infect insects by active penetration through the cuticle and are able to control insects regarding their feeding environment.
The insect cuticle is the first barrier that fungi encounter. Accordingly, a wide variety of extracellular enzymes are synthesized for the enzymatic processes involved in the degradation of protein, chitin and lipids, which are the basic cuticle components (St. Leger et al. 1986). Different studies suggested that proteases and chitinases are major determinants of fungal virulence in the complex and multifactorial phenomenon insect host/pathogen relationship (Fang et al. 2007).
Protease Pr1 was also found as a virulence indicator in EPF (Castellanos-Moguel et al. 2007). High pathogenicity on Heliothis armigera and cotton aphid has been observed by locally isolated EPF grown on Sabouraud dextrose medium by laboratory bioassay methods (Shahid et al. 2012). These experiments were designed to check similarities at the level of virulence-determining factors. VCP1 (VCP1 as abbreviation means “Verticillium chlamydosporium”) serine-like protease virulence factors, which are pathogenic toward both Heliothis and aphids, were grown in liquid medium at a large scale (Shahid et al. 2003).
The activities of protease have been determined in M. anisopliae var. anisopliae, B. bassiana, and Verticillium lecanii when grown on the cuticle of the locust Schistocerca gregaria (St. Leger et al. 1986).
Thereby, the main objective of the present study was to evaluate the most effective fungi against M. dirhodum and the production of an alkaline protease using Locust meal of Ocneridia voleximii (Pamphaginae, Orthoptera) as a basal substrate, which suggested to select the most widespread strains for biological control of aphids.
Materials and methods
Insects’ rearing
Metopolophium dirhodum was collected from a wheat variety (Cirta HD.122) (Triticum eastivum L.). The aphid was cultured on wheat for several generations, before the commencement of the experiment, at the National Institute of Plant Protection’ suburb (INPV) of Constantine-Algeria.
Fungal isolates
The virulence of three fungal isolates (Beauveria bassiana, Cladosporium cladosporioides, and Verticillium alfalfae) was tested. The fungi used in this study were isolated from an agricultural soil of the INPV. Isolation of EPF was determined according to the method described by (Vinayaga Moorthi et al. 2015), with minor modification. One gram (1 g) of the soil was diluted in 9 ml of distilled water, and then, 100 μl of the dilutions (10−3, 10−4, 10−5) was planted in Potatoes Dextrose Agar (PDA: 200 g potatoes, 20 g D. Glucose, and 20 g Agar), supplemented with Chloramphenicol (10 mg/1 L).The dishes were incubated for 2 weeks at 28 °C. The macroscopic and microscopic of resultant fungal growths were compared in terms of taking into account the standard description of mycelium and spore (De Hoog 1972; Samson et al. 1988).
Pathogenicity bioassay
The aphids were immersed for 10 s in each fungal suspension, which was prepared at the concentration (1.107conidia/ml) of sterile distilled water and supplemented with a drop of Tween 80 (0.05%). Dishes were kept at room temperature of 25 °C and 60% R.H. The mortality rates of the aphids were evaluated on 1st, 3rd, 5th, and 7th day after inoculation. All the treatments were repeated six times. Obtained data were analyzed. Tukey test was applied after one-way ANOVA for numbers of live individuals, using SPSS® for data analysis (version 17.00 Software. 2006 SPSS.Inc.Chicago.il, USA). The percentage effects of the three fungal isolates were calculated and corrected by Abbott’s (1925) formula:
Enzymes’ essays
Screening for protease enzyme production
The three EPF (B. bassiana, C. cladosporioides, and V. alfalfae) were tested for their ability to produce the protease enzyme. Caseins hydrolysis was added to 20% of agar and then 0.2% of skimmed milk (called Candia) was also added to the agar plates (Larsen et al. 1998). The plates were then incubated at 28 °C for 3 days. The enzyme activity was indicated by the formation of a clear zone around the colonies.
Fungi and inoculum’s preparation
The tested fungal isolates were propagated on PDA medium in plates at 28 °C. After 2 weeks, the plates were scraped with 10 ml of sterile water that contained 0.05% Tween 80. The spore suspension was adjusted to the desired concentration (107 conidia/ml) after counting with Malassez cell.
Fermentation condition
The culture medium used for production of alkaline protease was carried out by using Locust meal of Ocneridia voleximii (Pamphaginae: Orthoptera) which is considered as a basic substrate provided by the laboratory of biosystems and ecology of Arthropod at Mentouri Brothers University of Constantine, Algeria. Locust material was dried in shade at room temperature and then ground by using a blender. Two grams of powdered locust was soaked in 50 ml of distilled water. A fungal suspension (1 ml) containing 107 of conidia/ml was inoculated into Erlenmeyer flasks containing 50 ml of culture medium with an adjusted pH (pH = 4.5). Flasks were kept at 28 °C under agitation at 120 g for 3 days. The mycelia mass was obtained by filtering the content of each flask and then drying it at 105 °C until it reached a constant weight. The filtrate was used as a crude enzyme extract for the enzyme’s assay. The experiment was triplicated, and the mean values were calculated.
Alkaline protease activity
Based on Mechakra et al. 1999 method, a proteolytic activity assay was done using casein as the substrate. The enzyme activity was determined by incubating 1 ml of crude enzyme extract (EE) with 2.5 ml of casein sodium citrate (0.02 M) and 1.5 ml of Tris-HCl (0.2 M/0.1 M) with pH = 8. The mixture was incubated for 30 min at 40 °C, and the reaction was stopped by adding 5 ml of Trichloro-acetic (TCA). Thereafter, it left for 30 min in ice. The precipitated casein was then filtered off. 0.5 ml of the filtrate was taken in a test tube with 2.5 ml of NaCO3 in 2% NaOH (0.1 N), and then, 0.25 ml of the follin Ciocalten reagent (1 ml diluted with 3-ml distilled water) was added. A proteolytic activity was determined by the absorbance reading at 750 nm. Blanks of the samples were prepared by adding the TCA before the addition of substrate. Tyrosine was used as standard. One unit of protease activity was defined as the amount of enzyme that produced 1 mM of Tyrosine per minute above conditions. Tyrosine was taken as the standard for all the enzyme assay calculations.
Results and discussion
Pathogenicity bioassay
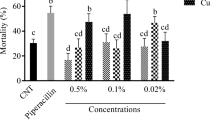
As illustrated in Fig. 1, 7 days post treatment, the rate of mortality of M. dirhodum varied relatively between 52 and 96%. The results revealed that B. bassiana had the most effective toxicity (95.83%), followed by C. cladosporioides (63.98%) and then V. alfalfae (51.83%).

Percentage mortality of three fungi on Metopolophium dirhodum
The scale values of the population density of M. dirhodum are given in Table 1. Aphid population densities at the treatments with the fungal isolates were compared with control at 1, 3, 5, and 7 days post treatment. Before treatment, the population densities were not significantly different at any treatment. The lowest population densities (1.00 and 0.017 individuals) were recorded on B. bassiana at 5 and 7 days post treatment, respectively, while the highest ones (2.67 and 1.67 individuals) were observed on V. alfalfae on the same dates. The respective population densities on C. cladosporioides were (2.33 and 1.33 individuals) at 5 and 7 days post treatment.
All isolates produced different mortality values among treatment; the highest mortality value was obtained on 7 days post treatment (F7DAA = 32.698, df = 3, p ≤ 0.00). The results’ statistics in 3rd and 5th were significant [(F3DAA = 8.333, df = 3, p ≤ 0.001); (F5DAA = 14.556, df = 3, p ≤ 0.00)] .
More than 750 fungal species were reported as potent agents against insect propagation (Gillespie et al. 1998). Saranya et al. (2010) stated that C. cladosporioides caused 77.50% mortality in Aphis craccivora (Koch) at a concentration of 108 conidia/ml. B. bassiana killed 96% of the aphids within 7 days after application. These results corroborate those of Murerwa et al. (2014) who reported that B. bassiana developed a considerable activity against M. dirhodum, and the mean mortalities ranged from 84 to 90% 10 days after application. Bensaci et al. (2015) revealed that the maximal mortality rate of C. oxysporum against Aphis fabae was 67.90% at the concentration of 108 conidia/m.
On the contrary, Abdel-Baky and Abdel-Salam 2003showed that the maximal mortality rates of Cladosporium spp. on Aphis gossypii and Aphis craccivora were 37.5 and 38%, respectively, 3 days after application with a concentration of 106 conidia/ml.
In the present study, the results that describe the effect of V. alfalfae on aphids were approximately similar to those reported by Chavan et al. (2008). Their results showed that the efficacy of V. locanii was high in a sense that they had an immense mortality rate ranged between 68.23 and 89.54% on aphids.
Alkaline protease activity
The screening of fungi in order to determine their protease activity was carried out by hydrolysis on Agar plates that contain 0.2% Skim milk in 20% agar. After incubation at 28 °C for 7 days, the enzyme activities were detected by the appearance of a zone around the fungal colonies. V. alfalfae showed the highest zone around the colony with ϕ = 5.1 cm, followed by C. cladosporioides with ϕ = 2 cm, and then B. bassiana with ϕ = 1.3 cm (Fig. 2).

Screening of fungi protease enzyme production, a Verticillium alfalfae, b Beauveria bassiana, c Cladosporium cladosporioides
Alkaline protease activity in the culture of locust meal of O. volexemii was detected at 28 °C for 3 days. The higher protease activities were observed for V. alfalfae with 95 U/ml, followed by B. bassiana with 38.26 U/ml, and then C. cladosporioides with 35, 65 U/ml.
The pH of the medium showed a large change in the case of isolates V. alfalfae (7.89), B. bassiana and C. cladosporioides (6.92) knowing that the initial pH was adjusted to 4.5.
This increase is explained by the release of alkaline molecules such as the ammonia that results from the degradation of proteins and leads to the decreases of the production of organic acids. This indicates also that the strains had certainly used the locust meal of O. vloximii, differently, and as the only source of carbon in the protein production. In effect, the variations of pH are considered as indicators of change in the metabolic process, including enzymes (Sandhya et al. 2005).
Extracellular proteases were even found in the insect haemolymph (Shimizuy et al. 1993). An immense variation in the protease activity was observed among the B. bassiana, C. cladosporioides, and V. alfalfae. A level of protease was observed in V. lecanii, and the highest levels were produced, solely, in cuticle substrate. Insect cuticle stimulated the protease production by both V. locanii and V. alboatum and with the adaptation of Verticillium spp. The protease production occurred in the insect cuticle that does not require substantial changes in the regulatory conditions of protease expression genes. These results are similar to those reported by (Barranco-Florido et al. 2002) when the proteolytic was lower submerged fermentation (cuticule of Sphenarium purpurascens, 20 g/l; (NH4)2 SO4,3 g/l; MgSO4, 0.6 g/l; NaCl, 0.5 g/l; KH2PO4, 10 g/l). They were 66.35 and 92% lower for proteolytic in the strains V. lecanii ATCC26854, ATCC 46578 and wild strain (WS).
In the present study, the protease activity by B. bassiana (38.26 U/ml) was approximately similar to those reported by Ito et al. (2007), demonstrating high protease activity on the 5th day of culture in B. bassiana. The time for maximum protease production does not necessarily depend upon the media constituents. Kucera (1971) observed high amounts of protease release 3 days after inoculation with different nitrogen sources. Proteolytic activity of B. bassiana is influenced by the culture age. This is probably due to the nutrient limitation or the autolysis of the culture (Braga et al. 1999).
Fungal pathogenesis of insect is related to many different factors such as hosting, parasitizing, and the environment as well. In the initial contact between fungus and insect, after the adhesion process, the penetration of the fungal pathogen into cuticle depends relevantly on the action of the enzymatic activities (St. Leger et al. 1987).
Jakovljević (2016) found that the alkaline protease activities of C. cladosporioides were enhanced by the addition of pollutant for 56.88% in in Czapek-Dox liquid nutrient medium with the addition of 0.5%.
Conclusion
Under the present experimental conditions, the results revealed that there was no relationship between a high alkaline protease activity and high virulence isolates. B. bassiana had the most effective toxicity (95.83%) but the high alkaline protease activity was observed in V. alfalfae (95 U/ml). Many factors influence directly and indirectly protease activity and the virulence isolates in this process. More research is needed about this complex mechanism.
Obtained results showed that the fungal isolates used might be utilized as biocontrol agents against the aphids. Among them, B. bassiana was the most promising one. Further studies should include evaluating the efficacy of the isolate in the field. However, this present work indicated the potentiality of V. alfalfae as a new resource of alkaline protease, which may suggest that protease activities could be used in the selection of candidates for aphids biological control.
References
Abbott WS (1925) A method of computing the effectiveness of an insecticide. J Econ Entomol 18:265–267
Abdel-Baky NF, Abdel-Salam AH (2003) Natural incidence of Cladosporium spp. as a bio-control agent against whiteflies and aphids in Egypt. J Appl Entomol 127(4):228–235
Barranco-Florido JE, Alatorre-Rosas R, Gutiérrez-Rojas M, Viniegra-González G, Saucedo-Castaňeda G (2002) Criteria for the selection of strains of entomopathogenic fungi Verticillium lecanii for solid state cultivation. Enzym Microb Technol 30:910–915
Bensaci OA, Daoud H, Lombarkia N, Rouabah K (2015) Formulation of the endophytic fungus Cladosporium oxysporum Berk. & M.A. Curtis, isolated from Euphorbia bupleuroides subsp. luteola, as a new biocontrol tool against the black bean aphid (Aphis fabae Scop.) J T Plant Prot Res 55(1):2015
Blackman RL, Eastop VF (2000) Aphids on the world’s crops: an identification and information guide. Wiley, Chichester, p 476
Braga GUL, Destefano RHR, Messias CL (1999) Protease production during growth and autolysis of submerged Metarhizium anisopliae cultures. Rev Microbiol 30:107–113
Carter N, McLean IFG, Watt AD, Dixon AFG (1980) Cereal aphids: a case study and review. In: Coaker TH (ed) Applied biology. Acad. Press, London, pp 271–348
Castellanos-Moguel J, Gonzalez-Barajas M, Mier T, Reyes Montes MR, Aranda E, Toriello C (2007) Virulence testing and extracellular subtilisin-like (Pr1) and tripsina-like (Pr2) activity during propagule production of Paecilomyces fumosoroseus isolates from whiteflies (Homoptera: Aeyrodidae). Rev Iberoam Micol 24:62–68
Chavan BP, Kadam JR, Saindane YS (2008) Bioefficacy of liquid formulation of Verticillium lecanii against aphid (Aphis gossypii). Int J Plant Prot 1(2):69–72
De Hoog GS (1972) The genera Beauveria, Isaria, Tritirachium and Acrodontium. Gen Nov Stud Mycol 1:1–41
Dixon AFG (1987) Cereal aphids as an applied problem. Agric Zool Rev 2(1):1–57
Fang WG, Gou SJ, Pei XQ, Zhang YJ, Xiao YH, Li DM, Jin K, Bidochka MJ, Pei Y (2007) Increased insect virulence in Beauveria bassiana strains overexpressing an engineered chitinase. Appl Environ Microbiol 73:295–302
Gillespie J, Bateman PR, Charnley AK (1998) Role of cuticule degrading proteases in the virulence of Metarhizium spp. for the desert locust, Schistocerca gregaria. J.Nvertebr. Pathol 71:128–137
Ito ET, Varea-Pereira G, Miyagui MT, Pinotti MHP, Neves PMOJ (2007) Production of extracellular protease by a Brazilian strain of Beauveria bassiana reactivated on coffee berry borer, Hypothenemus hampei. Braz Arch Biol Technol 50:217–223
Jakovljević VD, Vrvić MM (2016) Potential of pure and mixed cultures of Cladosporium cladosporioides and Geotrichum candidum for application in bioremediation and detergent industry. Saudi J Biol Sci Article in Press. http://www.sciencedirect.com/science/article/pii/S1319562X1600022X. Accessed 13 Feb 2016.
Kucera M (1971) Toxins of the entomophagous fungus Beauveria bassiana. II. Effect of nitrogen sources on formation of the toxic protease in submerged culture. J Invertebr Pathol 17:211–215
Larsen MD, Kristiansen KR, Hansen TK (1998) Characterization of the proteolytic activity of starter cultures of Penicillium roqueforti for production of blue veined cheeses. Int J Food Microbiol 43(3):215–221
Mechakra A, Auberger B, Remeuf F, Lenoir J (1999) Optimisation d’un milieu de culture pour la production d’enzymes prote’olytiques acides par Penicillium camemberti. Sci Aliment 19:663–675
Murerwa P, Futi AP, Wanjiku KA, Maniania NK (2014) Effect of infection by Metarhizium anisopliae isolate ICIPE 51 on developmental stage, fecundity and intrinsic rate of increase of Rhopalosiphum padi and Metopolophium dirhodum. J Entomol Nematol 6(11):154–160
Samson RA, Evans HC, Latge JP (1988) Atlas of entomopathogenic fungi. Sproner-Verlag, Berlin, Heidelberge, New York, p 187
Sandhya C, Sumantha A, Szakacs AG, Pandey A (2005) Comparative evaluation of neutral protease production by Aspergillus oryzae in submerged and solid-state fermentation. Process Biochem 40:2689–2694
Saranya S, Ushakumari R, Jacob S, Babu Philip M (2010) Efficacy of different entomopathogenic fungi against cowpea aphid, Aphis craccivora (Koch). Journal of Biopesticides 3(1 Special Issue):138–142
Shahid AA, Rao AQ, Bakhsh A, Husnain T (2012) Entomopathogenic fungi as biological controllers: new insights into their virulence and pathogenicity. Arch Biol Sci 64(1):21–42 2012 Belgrade
Shahid AA, Sattar A, Chaudhry B, Riazuddin S (2003) Determination of protein virulence factors and pathogenicity of entomopathogenic fungi. Pak J Biochem Mol Biol 36:100–107
Shimizuy S, Tsuchitan I, Matusumoto T (1993) Production of an extracellular protease by Beauveria bassiana in the haemolymph of the silkworm, Bombyx mori. Lett Appl Microbiol 16:291–294
St. Leger RJ, Charnley AK, Cooper M (1986) Cuticle degrading enzymes of entomopathogenic fungi: mechanisms of interaction between pathogen enzymes and insect cuticle. J Invertebr Pathol 47:295–302
St. Leger RJ, Cooperand M, Charnley AK (1987) Production of cuticle degrading enzymes by the entomopathogen Metarhizium anisopliae during infection of cuticles from Calliphora vomitoria and Manduca sexta. J Gen Microbiol 133:1371–1382
Vinayaga Moorthi P, Balasubramanian C, Selvarani S, Radha A (2015) Efficacy of sub lethal concentration of entomopathogenic fungi on the feeding and reproduction of Spodoptera litura. Springer Plus 4:681
Wells ML, McPherson RM, Ruberson JR, Herzog GA (2000) Effect of fungicide application on activity of Neozygites fresenii (Entomophthorales: Neozygi taceae) and cotton aphid (Homoptera: Aphididae) suppression. J Econ Entomol 93:1118–1126
Acknowledgements
The authors thank Süleyman Demirel Üniversitesi, Ziraat Fakültesi, Bitki Koruma Bölümü, 32260, Isparta-Türkiye, Mentouri Brothers Constantine, Faculty of SNV, Constantine, Algeria, and the National Institute of Agronomic Research Constantine-Algeria (INRA. Constantine), for their kind and great help.
Author information
Authors and Affiliations
Contributions
OA, MMS and AO collaborated in the creation of the manuscript. ALB and participated in the pathogenicity test. In addition to that, OA, MM, AO, FK and AB elaborated in the identification of the fungi and enzyme's test. Finally, BN is the one that produced locusts ( Ocniridia voleximii ). All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Abdelaziz, O., Senoussi, M.M., Oufroukh, A. et al. Pathogenicity of three entomopathogenic fungi, to the aphid species, Metopolophium dirhodum (Walker) (Hemiptera: Aphididae), and their Alkaline protease activities. Egypt J Biol Pest Control 28, 24 (2018). https://doi.org/10.1186/s41938-018-0030-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s41938-018-0030-7