
Abstract
Dengue, a global health threat, is a leading cause of morbidity and mortality in most tropical and subtropical countries. Dengue can range from asymptomatic, relatively mild dengue fever to severe and life-threatening dengue hemorrhagic fever. Disease severity and outcome is largely associated with the host immune response. Several candidate vaccines in clinical trials appear promising as effective measures for dengue disease control. Vaccine development has been hampered by safety and efficacy issues, driven by a lack of understanding of the host immune response. This review focuses on recent research findings on the dengue host immune response, particularly in humans, and the relevance of these findings to challenges in vaccine development.
Similar content being viewed by others

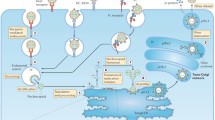

Background
Dengue virus (DENV) annually infects up to 390 million people living in tropical and subtropical areas [1]. It is estimated that up to half of the world’s population is at risk for DENV infection [2]. In recent years, several countries with subtropical regions, including France, Croatia, and Taiwan, reported autochthonous transmission of DENV [3–5]. DENV cases in Japan have been on the rise; 9 imported cases were reported in 1999 but increased to 249 cases in 2013. In 2014, the first autochthonous DENV transmission in 70 years was reported in Japan, where a total of 162 local DENV patients were reported. In addition, there were 179 imported cases in 2014, bringing the total DENV cases in 2014 to 341, the highest number of cases in recent years [6, 7]. Factors contributing to the rapidly increasing DENV cases over the past 70 years include expanding habitats of the mosquito vectors, Aedes aegypti and Aedes albopictus, due to urbanization and climate change, and increase in population. A. aegypti, a mosquito prevalent in the tropics, is closely associated with large dengue epidemics due to its ability to efficiently transmit DENV. In contrast, the habitat range of A. albopictus is not limited to the tropics but includes the temperate subtropical regions of Europe, North America, and East Asia. Because of the widespread distribution of A. albopictus, there is a potential threat of dengue spreading to wider geographical areas.
DENV belongs to the genus Flaviviridae, a group comprised of antigenically closely related viruses that cause disease in humans, including the Japanese encephalitis virus (JEV), yellow fever virus (YFV), and West Nile virus (WNV). The genome of DENV is a single, plus-stranded RNA which encodes three structural proteins (capsid [C], envelope [E], and preM proteins) and seven nonstructural proteins (NS1, NS2A, NS2B, NS3, NS4A, NS4B, and NS5). Infection with any of the four DENV serotypes that are antigenically and genetically related causes symptoms ranging from acute febrile illness to severe manifestations that include bleeding and organ failure. Without appropriate treatment, the mortality rate of dengue hemorrhagic fever (DHF) is approximately 25 %. Currently, there is no specific treatment or vaccine for dengue.
Clinical signs of dengue
DENV causes a wide range of clinical symptoms, ranging from asymptomatic infection and dengue fever (DF) to the severe forms of illness, DHF, and dengue shock syndrome (DSS). Symptoms typically appear 4–7 days after an incubation period following a bite from an infected mosquito. DF is a self-limited febrile illness characterized by a sudden onset of fever accompanied by a generalized morbilliform rash, headache, myalgias, and retro-orbital pain during the acute phase. The febrile phase generally lasts for a week. Symptoms are also accompanied by thrombocytopenia and leukopenia. A small proportion of dengue patients develop severe dengue (DHF or DSS). In severe dengue, life-threatening symptoms of plasma leakage, severe thrombocytopenia, hemorrhagic manifestations, and coagulation disorders may lead to shock and multiple organ failure during the critical phase of the disease. After the critical phase, most patients recover without any sequelae, although some develop fatigue and depression that lasts for several weeks after the acute phase. In severe dengue, an estimated 500,000 require hospitalization, and the mortality rate is 2.5 %, particularly in infants and young children [2]. Atypical clinical signs and symptoms of dengue include encephalitis, myocarditis, and respiratory distress [8].
Host immune responses to DENV
Humoral and cellular immune responses may play important roles in both disease pathogenesis and protection [9–12]. The paradoxical role of the host immune response in disease outcome is associated with consecutive infection by two different DENV serotypes. There are four serotypes of DENV, characterized by their antigenicity. The basis for host immunity in the disease outcome involves (1) virus-neutralizing antibodies directed against viral proteins and (2) immune cascades triggered by viral epitope-activated memory T cells [9–12].
Epidemiological studies have found that primary dengue patients, after a short cross-protective period, are at higher risk of developing severe dengue during a consecutive infection with another DENV serotype [12, 13]. The increased risk of developing severe dengue is hypothesized to be associated with non-protective serotype cross-reactive immunity induced after primary DENV infection. The basis for this is that after a primary DENV infection, serotype cross-reactive protective immunity wanes over a few months [14]. Neutralization activity in vitro was absent in diluted patient serum samples, indicating that a certain antibody threshold is needed for virus neutralization [14, 15]. Although serotype cross-reactive neutralizing antibodies confer protection at a certain threshold, waning immunity or a decline in antibody concentration induces susceptibility to infection with another DENV serotype.
Upon secondary infection with a heterogeneic serotype during this period of waning immunity, homotypic serotype-specific memory T and B cells are activated. This early serotype-specific immunity is directed against infection by the prior DENV serotype, but not to the current infecting serotype [9, 10]. During secondary infection, T cell responses directed against the previous infecting serotype do not confer protection and are associated with the triggering of immune cascades that induce severe symptoms [9].
As the disease progresses to the convalescent phase, serotype cross-reactive immunity with high avidity to all four DENV serotypes develops. These broadly reactive antibodies confer protection to all four DENV serotypes. A third or fourth infection may occur in hyperendemic areas, although a majority of these infections are hypothesized to be asymptomatic or mild [11].
Role of antibodies in pathogenesis of severe dengue
Epidemiological evidence has demonstrated that two groups of individuals are at higher risk of developing severe dengue: (1) adults with waning homotypic immunity and (2) infants born to dengue-immune mothers [13–15]. Serum samples obtained from these individuals during the period of waning immunity were found to enhance DENV infection in vitro [16]. Antibodies in these serum samples possess the ability to enhance DENV infection of Fcgamma (FcγR)-bearing cells in a phenomenon known as antibody-dependent enhancement (ADE) [16–20]. In ADE, subneutralizing levels of antibodies form a virus immune complex that enhances the infection of DENV target cells, the FcγR-bearing hematopoietic cell lines, leading to high levels of viremia [21–23]. Viremia determined by FcγR-bearing cells is up to 10 times greater than that determined by non-FcγR-bearing cells [13]. The resulting high level of viremia triggers the immune system, which in turn is associated with increased disease severity [22, 23].
Serum samples from convalescent DENV patients typically demonstrate ADE activity at higher dilutions and neutralizing activity at lower dilutions, indicating that antibodies induced after infection possess two contrasting activities [17]. In patients with acute secondary infection, undiluted serum samples demonstrated ADE activity against the infecting serotype, suggesting that ADE occurs in vivo [14–16]. ADE has also been demonstrated in animal models of secondary infection, in animals passively transferred with infection-enhancement antibodies, and in a maternally acquired heterotypic dengue antibody animal model [24–27].
Antibodies in protection against DENV infection
Neutralizing antibodies are considered central in protection against DENV infection. Antibodies targeting DENV possess two competing activities related to pathogenicity: virus neutralization and infection enhancement. At higher avidity, antibodies neutralize DENV, whereas lower levels of antibodies enhance DENV infection and hamper virus neutralization [17]. The concept was confirmed in studies of serum samples from dengue patients, in which neutralizing activity was present to several DENV serotypes using non-FcγR-expressing cells, but ADE activity that lowers the overall neutralizing activity was present to the current infecting serotype (DENV-1 and DENV-3), as demonstrated by FcγR-expressing cells (Table 1) [17]. After the cross-protection phase in patients with primary infection, antibodies demonstrate high levels of neutralizing activity to a homotypic serotype, but neutralizing activity against other serotypes is absent. Antibodies with high neutralizing activity to the prior infecting serotype also demonstrate cross-reactivity to other serotypes as well as infection-enhancement activity [7].
After acute infection, a high proportion of the antibodies is directed against the E protein, and some target the prM and NS1 proteins. The virion envelope is a lipid bilayer, which consists of the E and M proteins. A majority of the induced antibodies target the E protein; therefore, virus neutralization may involve neutralization of the E protein epitopes. The antibodies induced consistently react with the E protein, but there is variability in the reactivity to domains I, II, and III; the considerable variability in reactivity in each of the domains isolated from individual patients indicates that virus neutralization may involve several epitopes [28, 29].
In DENV hyperendemic regions, co-circulation of several serotypes demonstrates an epidemiological pattern in which outbreaks arise from a dominant serotype, in a 2–4-year cycle [30, 31]. These cycles persist because infection with one serotype confers only partial protection and may be driven by low immunity to each of the dengue serotypes, which then allows the virus to persist in the region. In hyperendemic regions, most adults are exposed to two or more serotypes and demonstrate seroconversion.
Determination of neutralizing activity of an antibody to DENV
As neutralizing activity of an antibody is considered central in providing protection against dengue, levels of neutralizing antibody have been used as a proxy for disease protection. Determination of levels of neutralizing antibody is also important to analyze the immune response after vaccination. Neutralizing antibody to dengue is determined by using the plaque reduction neutralization test (PRNT). The PRNT is regarded as the gold standard for determination of neutralizing antibodies [32]. High titers of neutralizing antibody titers are associated with protection against subsequent DENV infection. However, infection-enhancement activity has been demonstrated to lower neutralizing activity of antibodies in the presence of FcγR [17, 33]. Thus, the use of non-FcγR-bearing cells for assays poses a challenge in defining the true serological correlate to DENV protection in vivo and in vaccine efficacy trials. Because FcγR-bearing monocyte-lineage cells are the major targets of DENV infection in vivo, these cells may better reflect the biological properties of neutralizing antibodies [34]. Primary and established monocyte-lineage cell lines have also been used in neutralization tests [14, 15, 35–38].
Although assays to determine neutralization antibody titers have been developed, the correlate of protection for dengue has yet to be defined. For JEV, a titer of ≥1:10 at PRNT50 has been suggested as a threshold that demonstrates protection [39, 40]. For DENV, titers of ≥1:10 have been demonstrated in convalescent serum samples, but the cutoff has not been determined. Determination of cutoff titer that demonstrates protection is needed to better predict vaccine efficacy.
Although the PRNT is the most reliable test to determine neutralizing antibodies, it is laborious and requires skilled personnel. Considering the magnitude of the dengue problem, standardized high-throughput assays are needed for both research and industrial settings. Early studies using flow cytometry, peroxidase-antiperoxidase (PAP) and an enzyme-linked immunosorbent assay (ELISA) that employs FcγR-expressing cells have proved useful in the determination of neutralizing activity of DENV antibodies [35–38]. Tools that use the viral protein subunits and E protein epitopes are promising, as these tests are rapid compared to the PRNT [41, 42]. However, further studies are needed to validate these tests and the results compared to those of the conventional PRNT, to establish a robust immune correlate for disease protection [32].
Implications of current findings of the host antibody response for DENV vaccine design
Currently, there are no licensed dengue vaccine which are used clinically. Dengue vaccine research has been carried out for the past 70 years, beginning with the first isolation of the virus during epidemics in Hawaii and Japan. Within a few decades, live-attenuated virus, inactivated vaccine, chimeric flavivirus, DNA, and subunit vaccines have been developed (Table 2). However, vaccine immunogenicity, safety, and efficacy have been recurring issues in their use for control of the disease. Additional challenges hampering clinical development include the lack of a robust immune correlate for dengue, possible inhibitory or enhancing effects that are induced upon natural flavivirus infection and vaccination, and the need for definition of vaccine trial endpoints.
The observation of the development of severe dengue during heterotypic secondary infection implies the need for a vaccine that possesses the ability to simultaneously induce long-term protective immunity to all four serotypes. Multiple vaccine formulations are currently under development, and several candidates are currently in preclinical and clinical stages. Currently, the live-attenuated tetravalent chimeric YFV/DENV vaccine (CYD-TDV) is in the most advanced clinical trial stage (phase III) [43]. The phase I and II studies of the CYD-TDV vaccine have demonstrated that it is safe and tolerated, with confirmation of seroconversion and virological efficacy [44]. In multisite phase III clinical trials conducted in Asia and Latin America, the overall efficacy of the CYD-TDV candidate vaccine ranges from 30 to 64 % against DENV infection. The incidence of severe dengue (>80 % efficacy) and hospitalization was also lower in the vaccine cohort study. Surprisingly, vaccine efficacy against DENV-2 in comparison to the other serotypes was consistently low, ranging from 9 to 42 % [43–46]. The overall efficacy also varied with study site, suggesting that efficacy may be associated with epidemiological patterns and the host immunity baseline. While a lower incidence of severe dengue was seen in the vaccine cohort, results from the study demonstrated that seroconversion did not consistently reflect protective immunity [44].
Recent DENV vaccine clinical trials and patient cohort studies have found that despite the presence of seroconversion (PRNT50 >1:20) in vaccines, protection against DENV-2 was minimal [44]. Although conventional PRNT assays used in these studies are considered reliable in determining neutralizing antibodies as a proxy for disease protection, the assay uses FcγR-negative cells. A novel PRNT assay has demonstrated that neutralizing activity of some serum samples from dengue patients was absent or present at much lower titers when determined using FcγR-expressing BHK cells, as compared to non-FcγR-bearing cells [17]. Interestingly, neutralizing activity to the current infecting serotype was not detected by using FcγR-bearing cells but was present when non-FcγR-bearing cells were used for the assay (Table 1). These studies suggest that DENV-2 infection breakthrough despite seroconversion could be caused by the dominance of antibodies that might not confer virus neutralization in the presence of FcγR.
Alternatively, it is hypothesized that neutralizing activity of an antibody displays differential activity against DENV genotypes. In vitro and animal studies have demonstrated that differential neutralizing activity when Asian and American DENV-2 strains are induced after exposure to DENV-1 [47, 48]. Although multiple genotypes exist in a DENV serotype, the differences in the antigenicity of each genotype in relation to the resulting protection against different genotypes need further characterization.
In addition to protective efficacy issues, hypothetical safety concerns include (1) possible enhancement of clinical response to live-attenuated vaccine in individuals with a flavivirus-immune background, (2) non-protective or waning immunity in vaccinated individuals that could potentially increase the risk of severe dengue, and (3) insufficient attenuation of vaccine strains. The CYD-TDV candidate vaccine trial has demonstrated that the vaccine is safe, and efficacy has been demonstrated through lesser number of virologically confirmed cases within the 2-year study period [46]. However, long-term results would provide insights into waning immunity in vaccinated individuals and the risk of severe dengue [45]. Additionally, further studies on the immune response to dengue vaccines in non-endemic areas would provide a better understanding of the host immune response in individuals without a dengue-immune background.
With half of the world population at risk of DENV infection, an effective dengue vaccine that confers long-term protective immunity to all four dengue serotypes would contribute significantly to disease control. Other considerations in dengue disease control would include suitability of the vaccination program, cost constraints, and identification of disease burden. The CYD-TDV (Dengvaxia) vaccine has been approved as the first registered dengue vaccine in Mexico in December 2015. The CYD-TDV vaccine is also currently under review for clinical introduction in other endemic regions and is anticipated to reduce dengue burden. Follow-up observations on the CYD-TDV vaccine efficacy would however be needed, particularly on breakthrough infections and severe dengue in vaccine recipients. Other promising vaccine candidates in clinical and preclinical trials include virus-vectored and VLP-based vaccines. These next-generation vaccine candidates may offer improved approaches to disease control as they have been demonstrated to be safe due to the non-infectious nature of the vaccine, highly immunogenic, and could be modified rapidly to match recent epidemic strains. With dengue disease burden being the highest in developing countries, a cost-effective, safe vaccine that confers protection with a single dose would be ideal in disease control programs.
Conclusions
Dengue epidemics continue to be a serious public health problem in many areas of the world. Disease control remains a challenge in the absence of a vaccine and specific therapies. Development of tools and robust animal models to advance understanding of the host immune response will facilitate a better understanding of vaccine efficacy and safety issues and contribute to the development of dengue vaccine and treatment programs.
References
Bhatt S, Gething PW, Brady OJ, Messina JP, Farlow AW, Moyes CL, et al. The global distribution and burden of dengue. Nature. 2013;496(7446):504–7.
World Health Organization. Dengue and severe dengue haemorrhagic fever. Available at: http://www.who.int/mediacentre/factsheets/fs117/en/. Accessed 23 February 2016.
Marchand E, Prat C, Jeannin C, Lafont E, Bergmann T, Flusin O, et al. Autochthonous case of dengue in France, October 2013. Euro Surveill. 2013;18(50):20661.
Lin CC, Huang YH, Shu PY, Wu HS, Lin YS, Yeh TM, et al. Characteristic of dengue disease in Taiwan: 2002–2007. Am J Trop Med Hyg. 2010;82(4):731–9.
Gjenero-Margan I, Aleraj B, Krajcar D, Lesnikar V, Klobučar A, Pem-Novosel I, et al. Autochthonous dengue fever in Croatia, August-September 2010. Euro Surveill. 2011;16:9.
Moi ML, Kotaki A, Tajima S, Ikeda M, Yagasaki K, Lim CK, Kinoshita H, Nakayama E, Saito Y, Kurane I, Oishi K, Saijo M, Takasaki T. Trends of imported dengue fever cases in Japan, 2010 to 2013. Dengue Bulletin, in press, 2015
Kutsuna S, Kato Y, Moi ML, Kotaki A, Ota M, Shinohara K, et al. Autochthonous dengue fever, Tokyo, Japan, 2014. Emerg Infect Dis. 2015;21(3):517–20.
World Health Organization. Dengue haemorrhagic fever: diagnosis, treatment, prevention and control. 2nd ed. Geneva: World Health Organization; 1997.
Kurane I, Matsutani T, Suzuki R, Takasaki T, Kalayanarooj S, Green S, et al. T-cell responses to dengue virus in humans. Trop Med Health. 2011;39(4 Suppl):45–51.
Mathew A, Kurane I, Green S, Vaughn DW, Kalayanarooj S, Suntayakorn S, et al. Impaired T cell proliferation in acute dengue infection. J Immunol. 1999;162(9):5609–15.
Graham RR, Juffrie M, Tan R. A prospective seroepidemiologic study on dengue in children four to nine years of age in Yogyakarta, Indonesia. I. Studies in 1995–1996. Am J Trop Med Hyg. 1999;61:412–9.
Sangkawibha N, Rojanasuphot S, Ahandrik S, et al. Risk factors in dengue shock syndrome: a prospective epidemiologic study in Rayong, Thailand. I. The 1980 outbreak. Am J Epidemiol. 1984;120:653–69.
Sabin AB. Research on dengue during World War II. Am J Trop Med Hyg. 1952;1(1):30–50.
Kliks SC, Nisalak A, Brandt WE, Wahl L, Burke DS. Antibody-dependent enhancement of dengue virus growth in human monocytes as a risk factor for dengue hemorrhagic fever. Am J Trop Med Hyg. 1989;40:444–51.
Kliks SC, Nimmanitya S, Nisalak A, Burke DS. Evidence that maternal dengue antibodies are important in the development of dengue hemorrhagic fever in infants. Am J Trop Med Hyg. 1988;38:411–9.
Moi ML, Takasaki T, Saijo M, Kurane I. Dengue virus infection-enhancing activity of undiluted sera obtained from patients with secondary dengue virus infection. Trans R Soc Trop Med Hyg. 2013;107(1):51–8.
Moi ML, Lim CK, Chua KB, Takasaki T, Kurane I. Dengue virus infection-enhancing activity in serum samples with neutralizing activity as determined by using FcγR-expressing cells. PLoS Negl Trop Dis. 2012;6(2), e1536.
Moi ML, Lim CK, Kotaki A, Takasaki T, Kurane I. Detection of higher levels of dengue viremia using FcγR-expressing BHK-21 cells than FcγR-negative cells in secondary infection but not in primary infection. J Infect Dis. 2011;203(10):1405–14.
Ito M, Katakai Y, Ono F, Akari H, Mukai RZ, Takasaki T, et al. Serotype-specific and cross-reactive neutralizing antibody responses in cynomolgus monkeys after infection with multiple dengue virus serotypes. Arch Virol. 2011;156(6):1073–7.
Littaua R, Kurane I, Ennis FA. Human IgG Fc receptor II mediates antibody-dependent enhancement of dengue virus infection. J Immunol. 1990;144:3183–6.
Endy TP, Nisalak A, Chunsuttitwat S, Vaughn DW, Green S, Ennis FA, et al. Relationship of preexisting dengue virus (DV) neutralizing antibody levels to viremia and severity of disease in a prospective cohort study of DV infection in Thailand. J Infect Dis. 2004;189(6):990–1000.
Libraty DH, Endy TP, Houng HS, Green S, Kalayanarooj S, Suntayakorn S, et al. Differing influences of virus burden and immune activation on disease severity in secondary dengue-3 virus infections. J Infect Dis. 2002;185(9):1213–21.
Vaughn DW, Green S, Kalayanarooj S, Innis BL, Nimmannitya S, Suntayakorn S, et al. Dengue viremia titer, antibody response pattern, and virus serotype correlate with disease severity. J Infect Dis. 2000;181(1):2–9.
Moi ML, Takasaki T, Omatsu T, Nakamura S, Katakai Y, Ami Y, et al. Demonstration of marmosets (Callithrix jacchus) as a non-human primate model for secondary dengue virus infection: high levels of viraemia and serotype cross-reactive antibody responses consistent with secondary infection of humans. J Gen Virol. 2014;95(Pt 3):591–600.
De Alwis R, Williams KL, Schmid MA, Lai CY, Patel B, Smith SA, et al. Dengue viruses are enhanced by distinct populations of serotype cross-reactive antibodies in human immune sera. PLoS Pathog. 2014;10(10), e1004386.
Pinto AK, Brien JD, Lam CY, Johnson S, Chiang C, Hiscott J, et al. Defining new therapeutics using a more immunocompetent mouse model of antibody-enhanced dengue virus infection. mBio. 2015;6(5):e01316–15.
Ng JK, Zhang SL, Tan HC, Yan B, Martinez JM, Tan WY, et al. First experimental in vivo model of enhanced dengue disease severity through maternally acquired heterotypic dengue antibodies. PLoS Pathog. 2014;10(4), e1004031.
Tsai WY, Lai CY, Wu YC, Lin HE, Edwards C, Jumnainsong A, et al. High-avidity and potently neutralizing cross-reactive human monoclonal antibodies derived from secondary dengue virus infection. J Virol. 2013;87(23):12562–75.
Beltramello M, Williams KL, Simmons CP, Macagno A, Simonelli L, Quyen NT, et al. The human immune response to dengue virus is dominated by highly cross-reactive antibodies endowed with neutralizing and enhancing activity. Cell Host Microbe. 2010;8(3):271–83.
Wilder-Smith A, Yoksan S, Earnest A, Subramaniam R, Paton NI. Serological evidence for the co-circulation of multiple dengue virus serotypes in Singapore. Epidemiol Infect. 2005;133(4):667–71.
Endy TP, Nisalak A, Chunsuttiwat S, Libraty DH, Green S, Rothman AL, et al. Spatial and temporal circulation of dengue virus serotypes: a prospective study of primary school children in Kamphaeng Phet, Thailand. Am J Epidemiol. 2002;156(1):52–9.
Roehrig, J. Guidelines for plaque reduction neutralizing testing of human antibodies to dengue viruses. Geneva: World Health Organization; 2007. whqlibdoc.who.int/hq/2007/who_ivb_07.07_eng.pdf. Accessed 6 December 2015.
Moi ML, Lim CK, Kotaki A, Takasaki T, Kurane I. Discrepancy in dengue virus neutralizing antibody titers between plaque reduction neutralizing tests with Fcgamma receptor (FcgammaR)-negative and FcgammaR-expressing BHK-21 cells. Clin Vaccine Immunol. 2010;17(3):402–7.
Moi ML, Lim CK, Kotaki A, Takasaki T, Kurane I. Development of an antibody-dependent enhancement assay for dengue virus using stable BHK-21 cell lines expressing Fc gammaRIIA. J Virol Methods. 2010;163(2):205–9.
Rodrigo WW, Alcena DC, Kou Z, Kochel TJ, Porter KR, Comach G, et al. Difference between the abilities of human Fcgamma receptor-expressing CV-1 cells to neutralize American and Asian genotypes of dengue virus 2. Clin Vaccine Immunol. 2009;16(2):285–7.
Konishi E, Tabuchi Y, Yamanaka A. A simple assay system for infection-enhancing and -neutralizing antibodies to dengue type 2 virus using layers of semi-adherent K562 cells. J Virol Methods. 2010;163:360–7.
Guy B, Chanthavanich P, Gimenez S, et al. Evaluation by flow cytometry of antibody-dependent enhancement (ADE) of dengue infection by sera from Thai children immunized with a live-attenuated tetravalent dengue vaccine. Vaccine. 2004;22(27-28):3563–74.
Laoprasopwattana K, Libraty DH, Endy TP, et al. Dengue virus (DV) enhancing antibody activity in preillness plasma does not predict subsequent disease severity or viremia in secondary DV infection. J Infect Dis. 2005;192(3):510–9.
World Health Organization. Japanese encephalitis vaccine. Wkly Epidemiol Rec. 2006;81(34/35):331–40.
Oya A. Immunity and protection of epidemic in Japanese encephalitis. Naika. 1966;17:905–9 (in Japanese).
Libraty DH, Zhang L, Obcena A, Brion JD, Capeding RZ. Anti-dengue virus envelope protein domain III IgG ELISA among infants with primary dengue virus infections. Acta Trop. 2015;142:103–7.
Liu L, Wen K, Li J, Hu D, Huang Y, Qiu L, et al. Comparison of plaque- and enzyme-linked immunospot-based assays to measure the neutralizing activities of monoclonal antibodies specific to domain III of dengue virus envelope protein. Clin Vaccine Immunol. 2012;19(1):73–8.
Capeding MR, Tran NH, Hadinegoro SR, Ismail HI, Chotpitayasunondh T, Chua MN, et al. Clinical efficacy and safety of a novel tetravalent dengue vaccine in healthy children in Asia: a phase 3, randomised, observer-masked, placebo-controlled trial. Lancet. 2014;384(9951):1358–65.
Sabchareon A, Wallace D, Sirivichayakul C, Limkittikul K, Chanthavanich P, Suvannadabba S, et al. Protective efficacy of the recombinant, live-attenuated, CYD tetravalent dengue vaccine in Thai schoolchildren: a randomised, controlled phase 2b trial. Lancet. 2012;380(9853):1559–67.
Hadinegoro SR, Arredondo-García JL, Capeding MR, Deseda C, Chotpitayasunondh T, Dietze R, et al. Efficacy and long-term safety of a dengue vaccine in regions of endemic disease. N Engl J Med. 2015;373(13):1195–206.
Villar L, Dayan GH, Arredondo-García JL, Rivera DM, Cunha R, Deseda C, et al. Efficacy of a tetravalent dengue vaccine in children in Latin America. N Engl J Med. 2015;372(2):113–23.
Kochel TJ, Watts DM, Halstead SB, Hayes CG, Espinoza A, Felices V, et al. Effect of dengue-1 antibodies on American dengue-2 viral infection and dengue haemorrhagic fever. Lancet. 2002;360(9329):310–2.
Bernardo L, Yndart A, Vázquez S, Morier L, Guzmán MG. Antibody responses to Asian and American genotypes of dengue 2 virus in immunized mice. Clin Diagn Lab Immunol. 2005;12(2):361–2.
Funding
This work was supported in part by a research grant, Research on Emerging and Re-emerging Infectious Diseases (H26-shinkou-jitsuyouka-007) from the Japan Agency for Medical Research and Development (AMED), and a Grant-in-Aid for Young Scientists (B) from JSPS (26870872).
Disclosure statement
None.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare no competing interest.
Authors’ contributions
MLM drafted the article. TT and IK evaluated, edited, and critically revised the manuscript for intellectual content. All authors have given final approval to the version to be submitted.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Moi, M., Takasaki, T. & Kurane, I. Human antibody response to dengue virus: implications for dengue vaccine design. Trop Med Health 44, 1 (2016). https://doi.org/10.1186/s41182-016-0004-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s41182-016-0004-y