
Abstract
Background
Characterization of molecular factors regulating ovarian follicular development is critical to understanding its functional mechanism of controlling the estrous cycle, determining oocyte competency, and regulating ovulation. In previous studies, we performed next-gene sequencing to investigate the differentially expressed transcripts of bovine follicular granulosa cells (GCs) at the dominant follicle (DF) and subordinate follicle (SF) stages during the first follicular wave. This study aims to investigate the proteomic characterization of GCs of DF and SF in the bovine estrous cycle.
Results
In total, 3409 proteins were identified from 30,321 peptides obtained from liquid chromatograph-mass spectrometer analysis. Two hundred fifty-nine of these proteins were found to be expressed differently in DF and SF. Out of 259, a total of 26 proteins were upregulated (fold change≥2) and 233 proteins were downregulated (fold change≤0.5) in DF. Gene Ontology (GO) analysis of proteome data revealed the biological process, cellular component and molecular function of expressed proteins in DF and SF, while the Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis showed important signaling pathways associated with follicular development such as the PI3K-Akt, estrogen, and insulin signaling pathways. Immunoblotting results of OGN, ROR2, and HSPB1 confirmed the accuracy of the data. Bioinformatics analysis showed that 13 proteins may be linked to follicular development.
Conclusions
Findings from this study will provide useful information for exploring follicular development and function.
Similar content being viewed by others

Introduction
Bovine ovaries contain many follicles. Of these, only the DF has potential for ovulation. DF is, therefore, a critical source of biomarkers of ovarian follicular development and function. Bovine ovarian follicles grow in a wave-like pattern with typically 2 or 3 follicular waves per estrous cycle. Most of the follicles finally atresia during the process of follicular development, with one larger follicle ovulating and hence achieving ‘dominance’[1, 2]. In general, only the DF eventually ovulates during the last wave of follicular growth. In mammals, ovarian follicle development and atresia are complex, involving cell replication, specialization, differentiation and death. Previous studies reported a variety of hormones and growth factors that are closely associated with follicular development. GCs secrete hormones during follicle growth, particularly steroid hormones close to ovulation, such as progesterone and estradiol, which regulate critical phases of the reproductive cycle, including oocyte growth and maturation [3]. Estrogen plays a beneficial role in follicular development by reducing atresia and promoting GCs proliferation [4,5,6]. Conversely, androgens have been considered inhibiting to folliculogenesis [7, 8]. Follicle stimulating hormone (FSH), an endocrine factor, is critical to ovarian follicles growth because it regulates GCs proliferation and prevents apoptosis and estradiol production [9]. Gonadotropins and luteinizing hormones both regulate ovarian follicle growth and development [10]. Paracrine and autocrine factors such as insulin-like growth factor family, transforming growth factor β family, and fibroblast growth factor family modulate the response of follicle cells to gonadotropin signals [11]. Follistatin is involved in follicle cell proliferation, oocyte maturation, steroidogenesis and corpus luteum function [12, 13]. Cocaine and amphetamine-regulated transcript (CART), a pituitary gonadotropin, is involved in regulation of GCs estradiol production [14].
Despite our detailed knowledge of the hormones and growth factors that control ovarian follicular development (summarized above), the mechanism that regulates the selection of one developing follicle to gain dominance and continue to grow in each follicular wave remains unknown. Complete processing of DF maturation is complex, and multiple proteins are involved in this process to regulate functional differentiation of the follicles, so we used a proteomics approach to characterize the proteomic profile of bovine follicles and identify potential regulatory proteins and signaling pathways involved in follicular development and function. The present study can be used as a reference for further study of regulation mechanisms of follicular development in monotocous species.
Materials and methods
Animal care
All animal procedures were implemented in strict accordance with the principles outlined in “Guide for the care and use of Laboratory Animals” by the National Institute of Health.
Collection of GCs
The ovaries were collected from three Holstein dairy cows in October 2017, and follicular growth was observed and recorded using daily ultrasonography. After 5–7 days of estrus, ovaries were removed from cows when the largest follicle appeared and the growth rate was significantly higher than the growth rates of other follicles. The largest follicle and second largest follicle were collected. Each of the follicles was dissected into two halves, the GCs were removed by gentle scraping of the follicle wall with microsurgery forceps, and detached cells were obtained by washing two times with DPBS, a mixture of DPBS and GCs was centrifuged in Eppendorf tubes, the supernatant was discarded and stored at − 80 °C for further analysis.
Extraction and digestion of total proteins
GCs were lysed by ultrasonication and homogenization in a cold extraction buffer. Protein concentrations were determined using the Bradford Coomassie® Brilliant Blue G-250 method (BCA) according to manufacturer instructions and using bovine serum albumin (BSA) as a standard. 50 μg of protein from each sample were denatured, followed by the addition of 10 mM dithiothreitol and incubation for 1 h at 56 °C. Proteins were then alkylated with 55 mM iodoacetamide in the dark for 40 min at room temperature and were further subjected to digestion with 1 μg trypsin for 16–17 h at 37 °C.
Mass spectrometry (MS) analysis
The content of each peptide was determined and quantified by Capillary High Performance Liquid Chromatography (Eksigent 425, AB SCIEX) coupled with label-free mass spectrometry (LC-MS/MS). (A) H2O + 0.1% formic acid and (B) acetonitrile + 0.1% formic acid, were parts of the mobile phase. Desalting of the samples was performed online using a reversed-phase C18 trapping column (0.1 mm internal diameter, 20 mm length, 3 μm particle size; Waters). The peptides were then separated using a nano-column (0.75 mm internal diameter, 150 mm length, 5 μm particle size; Waters) at 0.3 μL/min. Peptides were eluted from the column using the following gradient: 5–80% B for 110 min, 80–5% B for 0.1 min, maintained at 5% for 120 min and then returned to the initial conditions. A mass spectrometer (Q-Exactive; Thermo Scientific, USA) was connected to the liquid chromatography apparatus to detect the eluted peptides. The separated peptide fragments were identified using a mass spectrometer operated in positive ion mode with electrospray ionization and collision-induced dissociation (CID). Full-scan MS spectra (350–1750 m/z) was acquired at a resolution of 70,000 with an automatic gain control (AGC) target value of 3e6 by electrospray ionization. The full-scan maximum injection time was 20 ms (millisecond), and the dynamic exclusion was set to 25.0 s. CID spectra were acquired at a resolution of 17,500 with an AGC target value of 2e5 and a maximum injection time of 80 ms. The isolation window was set to 2.0 m/z.
Data analysis
Raw data were imported into the Expressionists software (Proteome Discover 2.0) for processing, after which the quantification was performed based on the peak intensities of the report ions of the only unique peptides in the MS/MS Spectra. MS/MS spectra were searched against the Uniprot-COW FASTA database with the following mascot parameters: peptide mass tolerance for ±15 ppm and fragment mass tolerance for 20 mmu. Trypsin was used as the protein-cleaving enzyme, and the two missed cleavages were accepted. Carbamidomethylation of cysteine was designated as a fixed modification, and oxidation of methionine, acetylation on protein N-term were selected as variable modifications. The peptide confidence was high, peptide length was set to > 4, and peptide false discovery rate (FDR) was set to ≤0.01.
Bioinformatics analysis
GO analysis was used to classify the functions of the differentially expressed proteins (fold change ≥2), which could be categorized into three main categories: biological process, cellular component and molecular function. The most important biochemical metabolic pathways and signal transduction pathways were identified by KEGG pathway analysis. The GO analysis and KEGG pathway analysis were realized using the Protein Analysis Through Evolutionary Relationships (PANTHER) database (http://www.pantherdb.org).
Immunoblotting assay
To validate the global proteomics methodology, three proteins were selected to be measured by immunoblot analysis. The whole protein was extracted from bovine follicles for each group (DF and SF) using a protein extraction kit (Beyotime, China) according to manufacturer instruction. Protein concentrations were measured by the BCA method using BSA as the standard. Extracts were denatured at 97 °C for 5 min. Equal amounts (25 μg/lane) of protein from each sample were separated using 12% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and electro-blotted onto nitrocellulose (NC) membranes. Membranes were then incubated with primary antibodies (polyclonal antibody) at the following dilutions: anti-OGN (HA1136, 1:1000, Huaan, China), anti-ROR2 (HA1137, 1:500, Huaan, China), anti-HSPB1 (HA1138, 1:500, Huaan, China), anti-β-actin (1:1000, CWBIO, China). Primary antibodies were diluted in TBST and incubated overnight at 4 °C. After washing in TBS with 0.1% Tween-20, the membranes were incubated with horseradish peroxidase conjugated secondary antibody (1:10,000, CWBIO, China) for 2 h at room temperature. After washing, the membranes were detected using the eECL Western Blot kit (CWBIO, China) and exposed to film. The intensity of signals for each protein was quantified using Image-Pro Plus Software, version 6.0 (Media Cybernetics, USA) and normalized to values obtained from β-actin. All experiments were performed in triplicates.
Results
Top 40 highly expressed proteins in DF and SF
The top 40 highly expressed proteins in GCs of bovine DF and SF are shown below (Table 1). Many of them are known to be critical for follicular growth and development, including VIM, GSTA3, ATP5B, HSPA8, GAPDH, ATP5A1, EEF1A1, CYP11A1, KRT8, etc.
Identification of differentially expressed proteins
Based on a fold change greater than 2 with p < 0.05, 259 differentially expressed proteins were identified in GCs between DF and SF, including 26 upregulated proteins (fold change≥2) and 233 downregulated proteins (fold change≤0.5) in DF (Additional file 1).
GO analysis
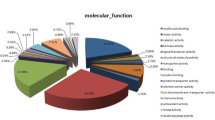
A total of 3409 proteins were identified from the 30,321 peptide sequences obtained from LC-MS/MS (FDR ≤ 0.01). Two thousand eight hundred ninety-five proteins and three thousand one hundred two proteins were identified in DF and SF, respectively. The expressed proteins in DF and SF were categorized under 3 major GO classifications: biological process, cellular component and molecular function. Among the biological processes, many biological processes were associated with follicular development, such as regulation of the apoptotic signaling pathway, the steroid biosynthetic process, the sterol biosynthetic process, the male gamete generation process, the G-protein coupled receptor signaling pathway, and the cell-cell signaling process (Fig. 1a). Regarding most molecular functions, the number of proteins in DF was similar to the number of proteins in SF. For example, 79 proteins in DF and 80 proteins in SF were involved in GTPase activity, 51 proteins in DF and 52 proteins in SF were involved in lyase activity, 37 proteins in DF and 39 proteins in SF that possessed the function of protein transporter activity were detected, etc. (Fig. 1b). Among cellular components, most of the proteins were assigned to the endopeptidase complex, an integral component of the organelle membrane, late endosome, mitochondrial ribosome and NADH dehydrogenase complex (Fig. 1c).

The expressed protein GO annotation. Classification of the expressed proteins in GCs of DF and SF in cattle based on the GO annotation, respectively. a: biological processes, b: molecular function, c: cellular components
Biological processes classifications of differentially expressed proteins
The differentially expressed proteins were categorized into 31 biological processes. Besides cellular component biogenesis and cellular macromolecular complex assembly, each of the other biochemical pathways includes either only up-regulated proteins or only down-regulated proteins (Fig. 2).

Biological processes of upregulated proteins and downregulated proteins
KEGG pathway analysis
KEGG pathway analysis showed that some important biochemical metabolic pathways and signal transduction pathways were associated with follicular development (Fig. 3), such as the Wnt signaling pathway, the estrogen signaling pathway, oocyte meiosis, progesterone-mediated oocyte maturation, the insulin signaling pathway, the FGF signaling pathway, the EGF receptor signaling pathway, apoptosis signaling pathway, the FoxO signaling pathway, and the p53 signaling pathway. Most importantly, the PI3K-Akt signaling pathway is a classical signaling pathway in follicular development regulation, including 8 differentially expressed proteins (HSP90AB1, GNG10, YWHAG, YWHAH, YWHAB, YWHAQ, RPS6, CDC37).

Differentially expressed proteins KEGG pathways analysis. The x-coordinate is the signaling pathway names, the y-coordinate is the number of proteins
Immunoblot validation
Three randomly selected proteins including OGN, ROR2, and HSPB1 were analyzed via immunoblotting. OGN increased significantly (P < 0.05) in DF compared with SF, ROR2 increased extremely significantly (P < 0.01) in SF compared with DF, and HSPB1 increased significantly (P < 0.05) in SF compared with DF. The expression tendency of the three proteins were consistent with MS data (Fig. 4) (Additional file 2).

Western blots for validation of OGN, ROR2 and HSPB1 abundance in DF and SF. β-actin was used as a loading control, and the abundance of proteins was corrected relatively to β-actin. On the left side of a-c: immunoblot results of OGN, ROR2, HSPB1 and β-actin in DF and SF GCs; on the right side of a-c: protein relative expression level of OGN, ROR2 and HSPB1 in DF and SF GCs. Superscript single and double asterisk indicate significantly different at the 0.05 and 0.01 levels, respectively. (n = 3 each; least square mean ± SE)
Discussion
This study identified numerous new candidate proteins potentially interacting with the follicular development mechanism. A total of 3409 proteins were identified in both types of follicles and 259 of them were identified as differentially expressed between DF and SF.
Our study provided novel information in bovine follicles and identified specific highly differentially expressed proteins in GCs such as HSPA5, HSP90B1, HSPA8, HSPB1, HSPD1, ATP5B, ATP5A1 and EEF1A1. Heat shock protein (HSP) expression is fundamental in the reproductive system of mammals, and different HSP members were also found in ovaries [15]. Out of 40 highly expressed proteins, 5 HSPs (HSPA5, HSP90B1, HSPA8, HSPB1, HSPD1) were detected. As ATP5A1 and ATP5B are highly expressed in glioblastoma tumor cells [16], they may be closely related to cell proliferation. Eukaryotic translation elongation factor 1 alpha (EEF1A) is known to participate in protein synthesis, and one isoform of EEF1A is EEF1A1. It is possible that the 8 proteins mentioned above promote follicular growth by associating with cell proliferation.
Most differentially expressed proteins are down-regulated, consisting mainly of ribosomal proteins, eukaryotic translation initiation factors, and tubulin, and these proteins could promote cell growth and proliferation. SF and DF in earlier waves eventually undergo atresia, whereas they are down regulated only in DF. The question is uncertain but meaningful, so it needs to be further research.
GO analysis of biological processes of expressed proteins in DF and SF showed that numbers of expressed proteins associated with steroid biosynthetic process in DF were greater than in SF. It is known that GCs secrete steroid hormones, which are related to follicle maturation. It is suggested that more steroids may be needed for ovulation in DF than in SF. In addition, 28 proteins in DF were identified in the male gamete generation process, whereas no proteins in SF were detected in this biological process, so it is possible that DF begins the preparation for fertilization. Biological process classifications of differentially expressed proteins showed most of the upregulated proteins were involved in the negative regulation of the epithelial cell apoptotic process, negative regulation of the extrinsic apoptotic signaling pathway, positive regulation of the hormone secretion, and the positive regulation of peptide hormone secretion. Most of the downregulated proteins were, instead, related to cell communication, signal transduction, ribosome biogenesis, and regulation of translational initiation (EIF4A1, EIF5, EIF3I, EIF1). In addition, the FSH receptor [17, 18], estrogen receptor [19] and CART [20] receptor are all G protein-coupled receptors (GPCRs). We found that some proteins were involved in GPCR signaling pathways in both types of follicles, suggesting GPCRs play an essential role during follicular development. In addition, GTPases play vital functions in cellular trafficking, cell division, and translation [21], and many proteins involved in GTPase activity were found by the GO analysis results of molecular functions.
The eminent event in DF and SF during a bonine follicular wave is size difference, as DF continues to grow, while SF begins to regress. KEGG analysis revealed differentially expressed proteins (YWHAG, YWHAH, YWHAB, YWHAQ) involved in the Hippo signaling pathway that play an essential role in maintaining organizational size [22,23,24]. YWHAG, YWHAH, YWHAB, and YWHAQ belong to the 14–3-3 family of proteins, which are considered as anti-apoptotic and critical regulatory proteins in cell division and apoptosis. These proteins may also play an important role in regulating cellular activities by associating with cytoskeletal proteins [22]. It is clear that Hippo and Akt signaling pathways regulate follicle growth, but most ovarian follicles are restricted to growth under physiological conditions due to local Hippo signaling [25].
Oxidative stress may be related to folliculogenesis and oogenesis [26] in the bovine species. SOD2 – mitochondrial can act directly on superoxide anion radicals – for example, estradiol inhibited SOD2 mRNA expression in rat luteal cells [27]. KEGG analysis showed that SOD2 participate in the FoxO signaling pathway, and SOD2 was upregulated in DF. These results may indicate that SOD2 plays an important role in DF.
Conclusions
The present study characterized the GCs proteome of bovine follicles at specific stages and screened 259 differentially expressed proteins in DF and SF. Many of these proteins may be linked to follicular development such as HSPA5, HSP90B1, HSPA8, HSPB1, HSPD1, ATP5B, ATP5A1, EEF1A1, 14–3-3 proteins (YWHAG, YWHAH, YWHAB, YWHAQ), and SOD2. Our findings will not only enrich the regulation theory of follicular development and dominance, but also have great significance to the application of propagation.
Availability of data and materials
Availability of data and materials are included in the manuscript, figures, and tables.
Abbreviations
- AGC:
-
Automatic gain control
- BCA:
-
Bradford Coomassie® Brilliant Blue G-250 method
- BSA:
-
Bovine serum albumin
- CART:
-
Cocaine and amphetamine-regulated transcript
- CID:
-
Collision-induced dissociation
- DF:
-
Dominant follicle
- EEF1A:
-
Eukaryotic translation elongation factor 1 alpha
- FDR:
-
False discovery rate
- FSH:
-
Follicle stimulating hormone
- GCs:
-
Granulosa cells
- GO:
-
Gene Ontology
- GPCRs:
-
G protein-coupled receptors
- HSP:
-
Heat shock protein
- KEGG:
-
Kyoto Encyclopedia of Genes and Genomes
- LC-MS/MS:
-
Capillary High Performance Liquid Chromatography coupled with label-free mass spectrometry
- MS:
-
Mass spectrometry
- NC:
-
Nitrocellulose
- PANTHER:
-
Protein Analysis Through Evolutionary Relationships
- SDS-PAGE:
-
Sodium dodecyl sulfate polyacrylamide gel electrophoresis
- SF:
-
Subordinate follicle
References
Ginther OJ, Bergfelt DR, Beg MA, et al. Follicle Selection in Cattle: Relationships among Growth Rate, Diameter Ranking, and Capacity for Dominance1[J]. Biol Reprod. 2001;65(2):345.
Manikkam M, Calder MD, Salfen BE, et al. Concentrations of steroids and expression of messenger RNA for steroidogenic enzymes and gonadotropin receptors in bovine ovarian follicles of first and second waves and changes in second wave follicles after pulsatile LH infusion[J]. Anim Reprod Sci. 2001;67(3–4):189–203.
Urrego R, Herrera-Puerta E, Chavarria NA, et al. Follicular progesterone concentrations and messenger RNA expression of MATER and OCT-4 in immature bovine oocytes as predictors of developmental competence[J]. Theriogenology. 2015;83(7):1179–87.
Payne RW, Hellbaum AA. The effect of estrogens on the ovary of the hypophysectomized rat[J]. Endocrinology. 1955;57(2):193.
Fortune JE. Ovarian follicular growth and development in mammals[J]. Biol Reprod. 1994;50(2):225–32.
Badinga L, Driancourt MA, Savio JD, et al. Endocrine and ovarian responses associated with the first-wave dominant follicle in cattle[J]. Biol Reprod. 1992;47(5):871–83.
Conway BA, Mahesh VB, Mills TM. Effect of dihydrotestosterone on the growth and function of ovarian follicles in intact immature female rats primed with PMSG[J]. J Reprod Fertil. 1990;90(1):267–77.
Mahajan DK. Steroidogenesis in human polycystic ovary[J]. Endocrinol Metab Clin North Am. 1988;17(4):751–69.
Hunzicker-Dunn M, Maizels ET. FSH signaling pathways in immature granulosa cells that regulate target gene expression: branching out from protein kinase a[J]. Cell Signal. 2006;18(9):1351–9.
Richards JAS, Russell DL, Ochsner S, et al. Ovulation: new dimensions and new regulators of the inflammatory-like response[J]. Annu Rev Physiol. 2002;64(1):69–92.
Buratini J, Price CA. Follicular somatic cell factors and follicle development[J]. Reprod Fertil Dev. 2011;23(1):32–9.
Silva CC, Knight PG. Modulatory actions of activin-a and follistatin on the developmental competence of in vitro-matured bovine oocytes[J]. Biol Reprod. 1998;58:558–65.
Glister C, Groome NP, Knight PG. Bovine follicle development is associated with divergent changes in activin-a, inhibin-a and follistatin and the relative abundance of different follistatin isoforms in follicular fluid[J]. J Endocrinol. 2006;188:215–25.
Smith GW, Sen A, Folger JK, et al. Putative role of cocaine- and amphetamine-regulated transcript (CARTPT) in dominant follicle selection in cattle[J]. Soc Reprod Fertil Suppl. 2010;67(8):105.
Velázquez MM, Salvetti NR, Amweg AN, et al. Changes in the expression of heat shock proteins in ovaries from bovines with cystic ovarian disease induced by ACTH[J]. Res Vet Sci. 2013;95(3):1059–67.
Xu G, Li JY. ATP5A1 and ATP5B are highly expressed in glioblastoma tumor cells and endothelial cells of microvascular proliferation[J]. J Neuro-Oncol. 2016;126(3):405–13.
Lazari MF, Liu X, Nakamura K, et al. Role of G protein-coupled receptor kinases on the agonist-induced phosphorylation and internalization of the follitropin receptor[J]. Mol Endocrinol. 1999;13(6):866.
Marion S, Robert F, Crepieux P, et al. G protein-coupled receptor kinases and beta arrestins are relocalized and attenuate cyclic 3′,5′-adenosine monophosphate response to follicle-stimulating hormone in rat primary Sertoli cells[J]. Biol Reprod. 2002;66(1):70–6.
Sharma G, Prossnitz ER. G-protein-coupled estrogen receptor (GPER) and sex-specific metabolic homeostasis[J]. Adv Exp Med Biol. 2017;1043:427.
Lv L, Jimenezkrassel F, Sen A, et al. Evidence supporting a role for cocaine- and amphetamine-regulated transcript (CARTPT) in control of granulosa cell estradiol production associated with dominant follicle selection in Cattle1[J]. Biol Reprod. 2009;81(3):580.
Vega FM, Thomas M, Reymond N, et al. The rho GTPase RhoB regulates cadherin expression and epithelial cell-cell interaction[J]. Cell Commun Signal. 2015;13:6.
Pan D. Hippo signaling in organ size control[J]. Genes Dev. 2007;21(8):886.
Halder G, Johnson RL. Hippo signaling: growth control and beyond[J]. Development. 2011;138(1):9.
Hergovich A. Mammalian hippo signalling: a kinase network regulated by protein-protein interactions[J]. Biochem Soc Trans. 2012;40(1):124.
Kawamura K, Cheng Y, Suzuki N, et al. Hippo signaling disruption and Akt stimulation of ovarian follicles for infertility treatment[J]. Proc Natl Acad Sci U S A. 2013;110(43):17474–9.
Combelles CMH, Holick EA, Paolella LJ, et al. Profiling of SOD isoenzymes in compartments of the developing bovine antral follicle[J]. Reproduction. 2010;139(5):871.
Sugino N, Hirosawatakamori M, Zhong L, et al. Hormonal regulation of copper-zinc superoxide dismutase and manganese superoxide dismutase messenger ribonucleic acid in the rat corpus luteum: induction by prolactin and placental lactogens[J]. Biol Reprod. 1998;59(3):599–605.
Acknowledgements
Authors are grateful of Prof. Sandeep Kumar Rajput and Prof. James Richard Pursley in Michigan State University in the USA for their directions of research designs and manuscript preparation.
Funding
This study was supported by Shanxi Scholarship Council of China Grant no. 2014-key 5, Shanxi Sci-technological Collaboration Grant no. 201603D421006, Shanxi Talent Introduction and Sanjin Talent Program, Shanxi Provincial Talent Introduction and SXAU (Shanxi Agricultural University) Major Research Achievement Cultivation Grant no. zdpy 201403/201503 to Lyu; Chinese Natural Science Foundation Grant no. 31873002, Shanxi Key Research and Development Plan (general) Agriculture Project Grant no. 201703d221020–1 to Li.
Author information
Authors and Affiliations
Contributions
HQ and LP created the idea for this study. LW and LL collected samples. HQ, ZZ, and XD performed lab work. HQ and LP wrote the manuscript. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
We confirm that this study did not involve relevant clause of the Ethics Committee, and all animal procedures were performed in strict accordance with the recommendations in the Guide for the Care and Use of Laboratory Animals of the National Institute of Health.
Consent for publication
All authors read and approved the final manuscript and consented for publication.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Additional files
Additional file 1:
259 differentially expressed proteins. (XLSX 54 kb)
Additional file 2:
Original immunoblotting results. (XLSX 9 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Hao, Q., Zhu, Z., Xu, D. et al. Proteomic characterization of bovine granulosa cells in dominant and subordinate follicles. Hereditas 156, 21 (2019). https://doi.org/10.1186/s41065-019-0097-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s41065-019-0097-5