
Abstract
Background
Species of Hirudo are used extensively for medicinal purposes, but are currently listed as endangered due to population declines from economic utilization and environmental pollution. In total, five species of Hirudo are currently described throughout Eurasia, with Turkey being one of the major exporters of medicinal leech, primarily H. verbana.
Results
To define the distribution of Hirudo spp. within Turkey, we collected 18 individuals from six populations throughout the country. Morphological characters were scored after dorsal and ventral dissections, and Maximum Likelihood (ML) and Bayesian Inference (BI) analyses resolved phylogenetic relationships using mitochondrial cytochrome c oxidase subunit I (COI), 12S ribosomal RNA (rRNA), and nuclear 18S rRNA gene fragments. Our results identify a new species of medicinal leech, Hirudo sulukii n. sp, in Kara Lake of Adiyaman, Sülüklü Lake of Gaziantep and Segirkan wetland of Batman in Turkey. Phylogenetic divergence (e.g., 10–14 % at COI), its relatively small size, unique dorsal and ventral pigmentation patterns, and internal anatomy (e.g., small and pointed atrium, medium-sized epididymis, relatively long tubular and arc formed vagina) distinguish H. sulukii n. sp. from previously described Hirudo sp.
Conclusions
By ML and BI analyses, H. sulukii n. sp. forms a basal evolutionary branch of Eurasian medicinal leeches. Phylogeographic interpretations of the genus identify a European Hirudo “explosion” during the upper Miocene followed by geological events (e.g., Zanclean flood, mountain building) that likely contributed to range restrictions and regional speciation of extant members of the clade.
Similar content being viewed by others

Background
Hirudinid leeches are parasitic to a variety of vertebrates leading many to regard them with distaste, but their medicinal utility is well established. For centuries, Hirudo medicinalis and related species (e.g., H. verbana, H. troctina) were prescribed to treat virtually every human ailment from arthritis to yellow fever, most without efficacy. In 1830, during their peak usage, a Paris hospital employed more than five million medicinal leeches [30]. Consequently, populations of H. medicinalis in Central Europe were depleted, and non-sustainable collecting led to their extinction in many areas. Pollution and habitat drainage further added to their decline, forcing Europe to import medicinal leeches from the Ottoman Empire (Anatolia), North Africa and Russia [31] to meet demand. By the late 1900’s, the advent of “modern” medicine drastically reduced clinical demand for leeches, allowing some threatened populations to rebound.
Leech therapy languished for most of the 20th century, considered “quackery” by mainstream medical practitioners [66], but the discovery of various bioactive compounds in leech saliva [27, 39], and recognition of the leech’s superior ability to relieve venous congestion (e.g., [58]), has led to renewed interest in clinical applications. Current fields of employment include reconstructive microsurgery, hypertension, and gangrene treatment [24]. In light of 19th century threats to medicinal leech populations as demand increased, considerable conservation steps were implemented to ensure their continued availability. Pursuant to these efforts, much confusion resulted regarding the taxonomic status of different morphological forms [18, 28, 56, 65]. Phylogenetic analysis of nuclear and mitochondrial DNA sequences suggest that the genus Hirudo is monophyletic [60], and that species or morphological varieties can be readily identified by coloration patterns. Molecular studies have shown that European medicinal leeches, although usually marketed as H. medicinalis, comprise a complex of at least three species: H. orientalis, the commonly sold H. verbana and the relatively rare H. medicinalis [4, 37, 54, 55, 60]. Kutschera and Elliott [36] analyzed the behavior of adult H. medicinalis, but could not find differences with respect to its sister taxon H. verbana. Morphological and molecular data demonstrate that commercially available medicinal leeches are generally not H. medicinalis [35, 56, 60], but rather specimens belonging to the Eastern phylogroup H. verbana [61, 62], which is predominantly bred in leech farms and used as a modern ‘medicinal’ stock.
Turkey is rich in wetlands and known to support at least two species of medicinal leech, H. medicinalis and H. verbana. Prior to ~2000, it was believed that medicinal leeches from Turkey’s wetlands were only H. medicinalis [21, 31]. Molecular characterization of Turkish leeches was not performed until the turn of the century, however, and leeches from the Kızılırmak and Yesilirmak Deltas on the Black Sea coast, comprising the majority of leech specimens destined for export, have proven to be to H. verbana [4, 51, 55].
Mapped localities of all Hirudo species show extensive, belt-shaped ranges extending from east to west. To establish the distribution of Hirudo species in Turkey, one of the major exporters of medicinal leeches worldwide, we sampled broadly in three representative localities within the western, eastern and southeastern regions of Turkey. Our data identifies a new species for the genus, H. sulukii n. sp., that forms a basal evolutionary branch among European medicinal leeches and sheds light on the evolutionary history of the genus.
Methods
Specimen collection and maintenance
Leech specimens collected throughout Turkey (Kara Lake, Beyaz Cesme Marsh, Uluabat Lake, Segirkan wetland, Balik Lake, Sülüklü Lake) were transported to Fırat University, Fisheries Faculty (Elazig, Turkey) and maintained in separate 600 L fiberglass tanks based on collection location. Tank bottoms were elevated with peat soil ~10 cm on one side to create a terrestrial to aquatic continuum. Leeches were fed one adult frog (e.g., Pelophylax ridibunda) blood meal per month (others have utilized mammalian blood), and typically survived 2+ years in the laboratory. Specimens were fixed in 70 % ethanol for molecular analysis and some were fixed with 10 % formaldehyde in PBS for dissection. External traits of live specimens were observed by stereomicroscopy. Preserved specimens were dissected dorsally and ventrally, with representative sketches of internal morphology derived directly from the type specimen.
DNA extraction
Tissue samples from live specimens were obtained by placing the leech in a 10 % ethanol sedating solution until it was unresponsive to touch. Approximately half of the caudal sucker was removed with a scalpel, and tissue cuttings were immediately processed using the E.Z.N.A.™ Tissue DNA kit (Omega Bio-Tek) following the manufacturer’s instructions. Whenever possible, tissue from postmortem specimens was taken from the caudal sucker to avoid contamination from gut contents.
DNA sequence amplification of target genes
Nuclear 18S rRNA, mitochondrial 12S rRNA and partial cytochrome c oxidase subunit 1 (COI) DNA fragments were amplified from genomic DNA using the polymerase chain reaction (PCR). All 12S sequences were obtained under conditions described by Borda and Siddall [8]. PCR amplification protocols were conducted as described by Wirchansky and Shain [67] employing primers listed in Table 1. PCR products were purified using the Wizard SV Gel and PCR Clean-Up System kit (Promega, Inc.) according to the manufacturer’s protocol.
DNA sequencing and editing
Purified PCR products were shipped to GeneWiz, Inc. (South Plainfield, NJ) for Sanger DNA sequencing using an ABI 3730xl DNA analyzer. Each PCR product was sequenced in both directions using amplification primers, and sequence chromatograms were viewed and manually adjusted in ChromasPro (Technelysium, Queensland, Australia) or BioEdit [26]. Sequence alignments were made with MUSCLE [17] or CLUSTAL W [29, 38]. Accession numbers for all CO1, 12S and 18S sequences are listed in Suppl. Data (Table 1).
Phylogeny
Maximum-likelihood (ML) analyses were performed for all DNA comparisons, using the pipeline sequence MEGA 7 [34] to align corresponding sequences from multiple individuals or homologous DNA across species, Gblocks [9] for alignment curation, PhyML [25] for tree building and TreeDyn [11] for tree drawing, as configured in the Phylogeny.fr platform [14]. The aLRT statistical test (approximation of the standard Likelihood Ratio Test; [3]) embedded in PhyML determined branch support values. Default settings were used for all parameters.
Bayesian Inference (BI) analysis was performed on the combined data set (morphological parameters, 18S, 12S, COI in Nexus format) in MrBayes v. 3.2.1x64 [48, 49]. Data were partitioned for 18S and 12S, and by codon position for COI. ModelTest [47] via FindModel was used to determine the optimal model of evolution for each gene under the Akaike Information Criterion (AIC; [46]). The general time reversible (GTR) model with a gamma distributed rate parameter was used for COI, 12S and 18S. Two analyses were run simultaneously with all parameter sets unlinked by partition for two million generations each, sampling every 100 generations, with a burn-in achieved by <50,000 generations. Setting the burn-in to 500,000 generations left a total of 7413 trees sampled for assessment of posterior probabilities. Gaps were treated as missing data, and default settings were used for all other parameters.
Results
Specimens of Hirudo were collected from multiple locations in Turkey (Fig. 1; Tables 2 and 3). These localities are separated by 1312 km (Uluabat Lake to Kara Lake), 1306 km (Uluabat Lake to Beyaz Cesme Marsh) and 289 km (Kara Lake to Beyaz Cesme Marsh). Leeches were typically found in muddy bottoms, as well as underwater and in aquatic/terrestrial vegetation (typically reedbeds), with banks of water proving the most prevalent habitat.

Specimens were scored for morphological characters according to Borda and Siddall [8], Utevsky and Trontelj [65], Klemm [33], Sawyer [53], Nesemann and Neubert [42], Saglam [50] and Govedich et al. [23], Elliott and Dobson [19] (Additional file 1). By these criteria, 10 leeches were identified as H. verbana, while six specimens did not match characters described for any known Hirudo species. Specifically, external pigmentation was unique, along with internal distinctions of the epididymis and vagina (see below).
Hirudo sulukii n. sp
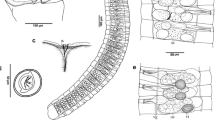
Based on morphological and genetic criteria, we formally propose the new species designation, Hirudo sulukii n. sp. (LSID: urn:lsid:zoobank.org:act:C338A26A-A205-4894-AB01-AA012293DD25), for leech specimens collected near Adiyaman, Batman and Gaziantep in southeastern Anatolia (Tables 2 and 3). The name “sulukii” is derived from the Turkish word “sülük” in reference to “leech”. Description based on holotype (specimen HS5 from Sülüklü Lake, catalogue ANSP G1 19488 in the Academy of Natural Sciences, Philadelphia, PA, USA). Paratypes deposited in the Academy of Natural Sciences (ANSP G1 19489) and Cukurova University Parasitology Museum, Adana, Turkey (CUPM-HIR/2016-1). Description: adult 64.06 ± 23.06 mm (27–105 mm) mean long, 6.71 ± 2.61 mm (4–12 mm) mean wide, mean width of anterior sucker 3.36 ± 1.10 mm (2–5.2 mm), mean width of posterior sucker 4.53 ± 1.33 mm (2–7 mm) (Fig. 2). Dorsum (Figs. 3 and 4a) pigmentation variably olive green, two orange paramedian stripes thin, two orange paramarginal stripes broad and encompassing black, segmentally-arranged united ellipsoid and elongated spots, dorsal lateral margins of body with yellow stripes encompassing zigzagged black longitudinal; covered with numerous papillae of body surface; background pigmentation of ventral (Figs. 3 and 4b) surface light greenish and covered with small number irregular dark markings. With classic Hirudo arc eyespot pattern [53], containing five pairs bilateral eyespots. Eyespots, five pairs on II, III, IV a1, V a1 and VI a2, forming a parabolic arc (Fig. 5). Number of annuli per somite: I-II-III: one, IV-V: two, VI-VII: three, VIII: four, IX: five (b1, b2, a2, b5, b6). Gonopores situated in furrow between annuli, separated by five annuli, male pore in the furrow XI b5/b6, female pore in the furrow XII b5/b6. Jaws trignathous, monostichodont, papillated.

Dorsal (a) and ventral (b) views of Hirudo sulukii n. sp. Holotype HS5 from Sülüklü Lake, Turkey (catalogue ANSP G1 19488)


Hirudo sulukii n. sp. Dorsal view (a) and ventral view (b); mg, male gonopore; fg, female gonopore. Based on holotype HS5 from Sülüklü Lake, Turkey (catalogue ANSP G1 19488)

View of anterior-dorsal and eyes of Hirudo sulukii n. sp. e, eyes. Based on holotype HS5 from Sülüklü Lake, Turkey (catalogue ANSP G1 19488)
Male reproductive apparatus notably large, with thick muscular penis sheath terminating in a bulbous prostate, located at ganglion in segment XI. Epididymis medium-sized, spherical, more than twice size of pearlescent-sheened ejaculatory bulb, tightly packed masses of ducting standing upright on either side of the atrium. Testisacs ovoid and larger than ovisacs, located posterior to ganglion in segment XIII. Female reproductive system relatively coiled tubing. The pearlescent-sheened vagina long and upright, evenly bowed tube entering directly into ventral body wall. Oviducts a thin duct forming several coiled and covered with a thick layer of glandular tissue, bi-lobed ovaries. Ovisacs globular ovoid or small bean seed-shaped (Fig. 6).

Hirudo sulukii n. sp. Dorsal view of reproductive system. a, Atrium; e, epididymis; eb, ejaculatory bulb; g, ganglion; o, ovisac; od, oviduct; ps, penis sheath; t, testisac, v, vagina. Based on holotype HS5 from Sülüklü Lake, Turkey (catalogue ANSP G1 19488)
Remarks
Despite similarities between Hirudo sulukii n. sp. and other Hirudo species, the former can be distinguished from its closest relatives using internal and external features. Hirudo sulukii n. sp. differs from H. medicinalis and H. orientalis by the form of black spots on the dorsal, paramedian stripes of the body. Hirudo sulukii n. sp. has black, segmentally-arranged united ellipsoid and elongated spots, and dorsal lateral margins of body a pair of zigzagged black dorsolateral longitudinal stripes (Fig. 4a). The ventral coloration pattern of H. sulukii n. sp. has a variable number of irregular spots (Fig. 4b); H. orientalis has black, dorsal rounded or quadrangular spots while H. medicinalis has elongated spots. The marginal spots of H. medicinalis are fused to form distinct black stripes. The ventral of H. medicinalis has an irregular dark mesh-like pattern while that of H. orientalis is more regular, formed by segmentally-arranged pairs of light markings on a predominantly black background. Hirudo verbana has broad, diffuse paramedian stripes orange in color. The ventral pattern of H. verbana is unicolored greenish to yellow, bounded by a pair of black ventrolateral stripes. Hirudo troctina has a pair of zigzag-shaped, black ventrolateral longitudinal stripes [65]. Hechtel and Sawyer [28] considered external pigmentation to be not only the most useful, but also arguably the best character to distinguish species of Hirudo.
In this study we used the approach of Hechtel and Sawyer [28] and Utevsky and Trontelj [65] regarding the size of the epididymis in relation to the ejaculatory duct. The epididymes of Hirudo sulukii n. sp. (Fig. 6) and H. orientalis are medium-sized. In contrast, the epididymes of H. verbana are relatively small, whereas H. troctina and H. medicinalis have massive epididymes [65]. The vagina of Hirudo sulukii n. sp. is relatively long tubular and arc formed (Fig. 6), while in H. orientalis the vagina is tubular and evenly curved. The former two species do not show the central swelling and sharp folding typical for H. verbana. In H. medicinalis, the vagina can have two conditions: straight and tubular, or terminally curved [65]. Hirudo troctina has a bulbous vagina [28].
Moquin-Tandon [40] described at least five species of Hirudo including H. verbana and H. medicinalis, but later concluded that they were all varieties of the same leech species. The medicinal leech, H. sulukii n. sp., considered here was determined to be morphologically different than all species described by Moquin-Tandor [40, 41].
Phylogenetic analyses
To determine the relationship of specimens to other Hirudo species, we subjected them to the comparative analysis of CO1 (cytochrome c oxidase subunit 1) and 12S rRNA from mitochondria, and nuclear 18S rRNA. Combined COI, 12S and 18S rRNA analysis contained 13 terminals with 1514 aligned characters. Maximum Likelihood of the combined data set yielded five equally parsimonious trees with 500–1000 steps (Fig. 7; Additional file 1); concordant trees were generated independently with COI data (Fig. 8; Additional file 1). Collectively, H. sulukii n. sp., formed a basal branch among European medicinal leeches with strong bootstrap support, while resolution among H. medicinalis, H. orientalis and H. verbana lineages was ambiguous, as noted in previous studies [45, 56]. Population structure was shallow among the collected specimens (<2 % divergence at CO1; Table 4), suggesting recent invasions into field sites sampled in the current study (see Fig. 1). The Asian species, H. nipponia, fell outside the Hirudo clade in combined sequence analyses (Fig. 7), suggesting a deep ancestral split with European species, and calling into question the designation of H. nipponia within the Hirudo phylogroup. Interestingly, H. nipponia was equidistant to European Hirudo species (~22–25 % at CO1), the latter of which were approximately equidistant to each other (i.e., ~10–14 % at CO1; Table 4). Inferring a divergence rate of ~2 % per million years at the CO1 locus based on combined geological and molecular data within Oligochaeta [10, 15, 67], we estimate a lower Miocene split between lineages leading to H. nipponia and European Hirudo sp., and radiation of the latter species during the upper Miocene. Branch patterns of remaining species were consistent with those reported previously [45].

Maximum Likelihood analysis of a combined COI, 12S and 18S data set (1514 total positions). Bootstrap values are indicated. European Hirudo species form a distinct clade with H. sulukii n. sp. as a basal member

Maximun Likelihood analysis of COI mtDNA haplotypes (542 total positions). The tree is drawn to scale, with branch lengths measured in the number of substitutions per site
Discussion
Maximum Likelihood and Baysian Inference analyses yielded trees with concordant topologies and strong support for H. sulukii as a basal branch of the European medicinal leeches. Relationships between H. medicinalis, H. verbana and H orientalis were less conclusive, consistent with confusion regarding their morphological identification [45, 56]. The relatively small size of H. sulukii, unique dorsal and ventral pigmentation patterns, and internal anatomy (e.g., small and pointed atrium, medium-sized epididymis, relatively long tubular and arc formed vagina) are distinguishing features of this previously undescribed leech. Note that H. sulukii has thus far been collected only from relatively high elevation field sites (i.e., Kara Lake-Adiyaman 1233 m, Sülüklü Lake-Gaziantep 877 m, and Segirkan wetland- Batman 525 m), and its small size in comparison with other Hirudo species may reflect an adaptation to this environment (e.g., reduced foraging season/food supply), as suggested for other annelid species (e.g., [15]).
Previously, only two medicinal leeches were thought to occur in Turkey, H. verbana and H. medicinalis, while a total of five are currently described throughout Eurasia. The range of H. verbana occurs to the south of H. medicinalis in an almost parapatric fashion with little overlap [5, 32, 42, 43, 51]. The former is subdivided into an Eastern (southern Ukraine, North Caucasus, Turkey and Uzbekistan) and Western phylogroup (Balkans and Italy) that do not overlap, suggesting distinct postglacial colonization from separate refugia [61, 64]. Easternmost records are from Samarqand Province in Uzbekistan [61, 64, 65], resulting in an east-to-west extent of ~4600 km. Leeches supplied by commercial facilities belong to the Eastern phylogroup, originating mostly from Turkey and the Krasnodar Territory in Russia, two leading areas of leech export.
Hirudo medicinalis is distributed from Britain and southern Norway to the southern Urals and probably as far as the Altai Mountains, occupying the deciduous arboreal zone [6, 12, 16, 21, 22, 31, 43, 51, 52, 59, 63, 68]. Hirudo orientalis is associated with mountainous areas in the sub-boreal eremial zone and occurs in Transcaucasian countries, Iran and Central Asia, while H. troctina has been found in northwestern Africa and Spain in the Mediterranean zone [64]. Hirudo verbana and H. medicinalis have recently experienced range expansions while H. orientalis has remained geographically isolated within arid and alpine areas of Central Asia and Transcaucasia [61].
By molecular clock inference using divergence estimates at the CO1 locus [10, 15, 67], our data suggest a deep, ancestral split between European and Asian (i.e., the lineage leading to H. nipponia) medicinal leeches somewhere in the lower Miocene, followed by an “explosion” of Hirudo species upon their putative arrival to the European continent during the upper Miocene, 5–10 mya (Fig. 9). The possible misclassification of H. nipponia does not affect this evolutionary scenario since it represents a basal, sister branch to the European Hirudo phylogroup (see Fig. 8). This evolutionary timeline is supported by tree topologies and relative genetic distance of European Hirudo species to each other at the COI locus (i.e., 10–14 % divergence; see Table 4). The time frame of these events suggest the presence of an open habitat corresponding with, for example, formation of Levantine land bridges, which may have facilitated mammalian-based, passive dispersal of an ancestral Hirudo archetype throughout Europe. Thereafter, tectonic activity at the onset of the Pliocene ~5.3 mya broke the land bridge between Morocco and Spain causing the Zanclean Flood that filled the Mediterranean basin, and in combination with mountain building throughout the European continent [7], appears to have restricted panmixia among extant Hirudo lineages, leading in part to their speciation and current geographic ranges. For instance, concurrent with the closing of the Tethys Sea by continental drift of the African and Arabian plates, mountain building events occurred in Southern Turkey forming the Taurus Mountain chain [13]. At present, the Anatolia region has been isolated completely by the southeastern Taurus Mountains [1], likely isolating H. sulukii from other European Hirudo populations.

Proposed biogeographical history of Eurasian Hirudo species based on phylogeny and current distribution patterns. Colored regions indicate reported geographic ranges of respective Hirudo species; the arrow topology reflects relationships between species based on Maximum Likelihood and Baysian phylogenies
Species of Hirudo have had broad applications in medicine, ranging from reconstructive surgeries (e.g., facial, finger reattachment, ear flap) to anticoagulants/analgesics secreted from salivary glands [2, 24]. Thus the discovery of a new Hirudo species, particularly a basal member of this phylogroup, has considerable value in the context of medical potential. Specifically, natural variants of known bioactive factors (e.g., hirudin, antistasin, etc.) are logical candidates to explore for their potentially enhanced or novel pharmaceutical properties. The current study has prompted a more systematic survey of Hirudo throughout Turkey and surrounding regions with the collective aims of refining the evolutionary history of the genus, facilitating conservation efforts, and identifying species that may expand the repertoire of medicinal applications for this important Hirudinid genus.
Conclusions
By phylogenetic and morphological criteria, specimens collected from Kara Lake of Adiyaman, Sülüklü Lake of Gaziantep and Segirkan wetland of Batman in Turkey comprise a new species, Hirudo sulukii. Geographic isolation by the Taurus Mountain chain has likely contained H. sulukii within the regional sampling area. By ML and BI analyses, H. sulukii n. sp. forms a basal evolutionary branch of Eurasian medicinal leeches, preceded by a deeper ancestral split with the Asian medicinal leech. H. nipponia. Phylogeographic interpretations of the genus identify a European Hirudo “explosion” during the upper Miocene followed by geological events (e.g., Zanclean flood, mountain building) that likely contributed to range restrictions and regional speciation of extant members of the European clade.
References
Altinli E. Geology of Eastern and Southeastern Anatolia. Bull Mineral Res Explor Inst Turk. 1966;66:35–76.
Abdualkader AM, Ghawi AM, Alaama M, Awang M, Merzouk A. Leech Therapeutic Applications. Indian J Pharm Sci. 2013;75:127–37.
Anisimova M, Gascuel O. Approximate likelihood-ratio test for branches: A fast, accurate, and powerful alternative. Syst Biol. 2006;55:539–52.
Apakupakul K, Siddall ME, Burreson EM. Higher level relationships of leeches (Annelida: Clitellata: Euhirudinea) based on morphology and gene sequences. Mol Phylogenet Evol. 1999;12:350–9.
Balık S, Ustaoğlu MR, Sarı HM, Özdemir Mis D, Aygen C, Taşdemir A, Yıldız S, Topkara ET, Sömek H, Özbek M, İlhan A. A preliminary study on the biological diversity of Bozalan Lake (Menemen- İzmir). E.U. J Fish Aquat Sci. 2006;23:291–4.
Bat L, Akbulut M, Culha M, Sezgin M. The macrobenthic fauna of Sırakaraağaçlar Stream flowing into the Black Sea at Akliman, Sinop. Turk J Mar Sci. 2000;6:71–86.
Blondel J, Aronson J, Bodiou JY, Boeuf G. The Mediterranean region, biological diversity in space and time. 2nd ed. New York: Oxford University Press Inc; 2010. p. 376.
Borda E, Siddall ME. Arhynchobdellida (Annelida: Oligochaeta: Hirudinida): phylogenetic relationships and evolution. Mol Phylogenet Evol. 2004;30:213–25.
Castresana J. Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis. Mol Biol Evol. 2000;17(4):540–52.
Chang CH, Lin SM, Chen JH. Molecular systematics and phylogeography of the gigantic earthworms of the Metaphire formosae species group (Clitellata, Megascolecidae). Mol Phylogenet Evol. 2008;49:958–68.
Chevenet F, Brun C, Banuls AL, Jacq B, Chisten R. TreeDyn: towards dynamic graphics and annotations for analyses of trees. BMC Bioinformatics. 2006;7:439.
Demirsoy A, Kasparek M, Akbulut A, Durmus Y, Emir Akbulut N, Çaliskan M. Phenology of the medicinal leech, Hirudo medicinalis L. in north-western Turkey. Hydrobiologia. 2001;462:19–24.
Dercourt J, Zonenshain LP, Ricou LE, Kazmin VG, Le Pichon X, Knipper AL, Grandjacquet C, Sbortshikov IM, Geyssant J, Lepvrier C, Pechersky DH, Boulin J, Sibuet J-C, Savostin LA, Sorokhtin O, Westphal M, Bazhenov ML, Lauer JP, Biju Duval B. Geological evolution of the Tethys belt from the Atlantic to the Pamirs since the Lias. Tectonophysics. 1986;123:241–315.
Dereeper A, Guignon V, Blanc G, Audic S, Buffet S, Chevenet F, Dufayard JF, Guindon S, Lefort V, Lescot M, Claverie JM, Gascuel O. Phylogeny.fr: robust phylogenetic analysis for the non-specialist. Nucleic Acids Res. 2008;36:W465–9.
Dial CR, Dial RJ, Saunders R, Lang SA, Tetreau MD, Lee B, Wimberger P, Dinapoli MS, Egiazarov AS, Gipple SL, Maghirang MR, Swartley-McArdle DJ, Yudkovitz SR, Shain DH. Historical biogeography of the North American glacier ice worm, Mesenchytraeus solifugus (Annelida: Oligochaeta: Enchytraeidae). Mol Phylogenet Evol. 2012;63:577–84.
Duran M, Akyıldız GK, Özdemir A. Gökpınar Çayı’nın Büyük Omurgasız Faunası ve Su Kalitesinin Değerlendirilmesi. Türk Sucul Yaşam Dergisi. 2007;5:577–83.
Edgar RC. MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004;32(5):1792–7.
Elliott JM, Kutschera U. Medicinal leeches: historical use, ecology, genetics and conservation. Freshw Rev. 2011;4:21–41.
Elliott JM, Dobson M. Freshwater Leeches of Britain and Ireland. Keys to the Hirudinea and a Review of their Ecology. Freshwater Biological Association Scientific Publication No: 69. 2015. p. 1–108.
Folmer O, Black M, Hoeh W, Lutz R, Vrijenhoek R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol Mar Biol Biotechnol. 1994;3:294–9.
Geldiay R. Çubuk Barajı ve Emir Gölünün Makro ve Mikro Faunasının Mukayeseli İncelenmesi. Ankara Üniv Fen Fak Mecm. 1949;2:106.
Geldiay R, Tareen IU. Bottom Fauna of Gölcük Lake. 1. Population study of Chironomids, Chaoborus and Oligochaeta. İzmir: E.Ü.F.F. İlmi Raporlar Serisi No:137; 1972. p. 15.
Govedich FR, Bain BA, Moser WE, Gelder SR, Davies RW, Brinkhurst RO. Annelida (Clitellata): Oligochaeta, Branchiobdellida, Hirudinida, and Acanthobdellida. In: Thorp JH, Covich AP, editors. Ecology and classification of North American freshwater invertebrates 3rd edition, Academic press. 2009. p. 385–410.
Gödekmerdan A, Arusan S, Bayar B, Sağlam N. Tıbbi Sülükler ve Hirudoterapi. Turkiye Parazitol Derg. 2011;35:234–9. doi:10.5152/tpd.2011.60.
Guindon S, Gascuel O. A simple, fast, and accurate algorithm to estimate large phylogenies by maximum likelihood. Syst Biol. 2003;52(5):696–704.
Hall TA. BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp Ser. 1999;41:95–8.
Haycraft JB. On the action of secretion obtained from the medicinal leech on coagulation of the blood. Proc R Soc Lond. 1884;36:478.
Hechtel FOP, Sawyer RT. Toward a taxonomic revision of the medicinal leech Hirudo medicinalis Linnaeus, 1758 (Hirudinea: Hirudinidae): re-description of Hirudo troctina Johnson, 1816 from North Africa. J Nat Hist. 2002;36(11):1269–89.
Higgins D, Thompson J, Gibson T, Thompson JD, Higgins DG, Gibson TJ. CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994;22:4673–80.
Kaestner A. Invertebrate zoology, vol. I. New York: Interscience; 1967. p. 597.
Kasparek M, Demirsoy A, Akbulut A, Emir Akbulut N, Çaliskan M, Durmus Y. Distribution and status of the medicinal leech (Hirudo medicinalis L.) in Turkey. Hydrobiologia. 2000;441:37–44.
Kazancı N, Ekingen P, Türkmen G. A study on Hirudinea fauna of Turkey and habitat quality of the species. Rev Hydrobiol. 2009;1:81–95.
Klemm DJ. The leeches (Annelida: Hirudinea) of North America. Cincinnati: Aquatic Biology Section, Environmental Monitoring and Support Laboratory, United States Environmental Protection Agency; 1982.
Kumar S, Stecher G, Tamura K. MEGA7: molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol Biol Evol. 2016;33:1870–4. doi:10.1093/molbev/msw054
Kutschera U. Leeches underline the need for Linnaean taxonomy. Nature. 2007;447:775.
Kutschera U, Elliott JM. The European medicinal leech Hirudo medicinalis L.: morphology and occurrence of an endangered species. Zoosyst Evol. 2014;91:271–80. doi:10.3897/zse.90.8715.
Kvist S, Oceguera-Figueroa A, Siddall ME, Erseus C. Barcoding, types and the Hirudo files: using information content to critically evaluate the identity of DNA barcodes. Mitochondrial DNA. 2010;21:198–205. doi:10.3109/19401736. 2010.529905.
Larkin MA, Blackshields G, Brown NP, Chenna R, McGettigan PA, McWilliam H, Valentin F, Wallace IM, Wilm A, Lopez R, Thompson JD, Gibson TJ, Higgins DG. ClustalW and ClustalX, version 2. Bioinformatics. 2007;23(21):2947–8.
Markwardt F. Untersuchungen über Hirudin. Naturwissenschaften. 1955;52:537.
Moquin-Tandon A. Monographie de la Famille des Hirudinees. Montpellier: Maison de Commerce; 1827.
Moquin-Tandon A. Monographie de la Famille des Hirudinees. Paris: Balliere; 1846.
Nesemann H, Neubert E. Annelida: Clitellata: Branchiobdellida, Acanthobdellea, Hirudinea. In: Süßwasserfauna von Mitteleuropa, 6/2. Heidelberg, Berlin: Spektrum Akad Verl; 1999.
Özbek M, Sarı HM. Batı Karadeniz Bölgesi’ndeki Bazı Göllerin Hirudinea (Annelida) Faunası. E.Ü. Su Ürünleri Dergisi. 2007;24:83–8.
Palumbi SR. Nucleic acids II: the polymerase chain reaction. In: Hillis DM, Moritz C, Mable BK, editors. Molecular systematics. Sunderland: Sinauer & Associates Inc.; 1996. p. 205–47.
Phillips AJ, Siddall ME. Poly-paraphyly of Hirudinidae: many lineages of medicinal leeches. BMC Evol Biol. 2009;9:246. doi:10.1186/1471-2148-9-246.
Posada D, Buckley TR. Model selection and model averaging in phylogenetics: advantages of Akaike Information Criterion and Bayesian approaches over likelihood ratio tests. Syst Biol. 2004;53:793–808.
Posada D, Crandall KA. MODELTEST: testing the model of DNA substitution. Bioinformatics. 1998;14:817–8.
Ronquist F, Huelsenbeck J, Teslenko M. Draft MrBayes version 3.2 manual: tutorials and model summaries. 2011. p. 172.
Ronquist F, Huelsenbeck JP. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics. 2003;19:1572–4.
Saglam N. Key of freshwater and marine leeches. Elazığ, Turkey: Fırat Üniversitesi Basım Evi; 2004. p. 38.
Saglam N. Protection and sustainability, exportation of some species of medicinal leeches (Hirudo medicinalis L., 1758 and Hirudo verbana Carena, 1820). J FisheriesSciencescom. 2011;5(1):1–15.
Saglam N, Dorucu M, Ozdemir Y, Seker E, Sarieyyupoglu M. Distribution and economic importance of medicinal leech, Hirudo medicinalis (Linnaeus, 1758) in Eastern Anatolia/Turkey. Lauterbornia. 2008;65:105–18.
Sawyer RT. Leech biology and behavior. New York: Oxford University Press; 1986.
Siddall ME, Apakupakul K, Burreson EM, Coates KA, Erséus C, Gelder SR, Källersjö M, Trapido-Rosenthal H. Validating livanow: molecular data agree that leeches, branchiobdellidans, and acanthobdella peledina form a monophyletic group of oligochaetes. Mol Phylogenet Evol. 2001;21:346–51.
Siddall ME, Burreson EM. Phylogeny of leeches (Hirudinea) based on mitochondrial cytochrome c oxidase subunit I. Mol Phylogenet Evol. 1998;9:156–62.
Siddall ME, Trontelj P, Utevsky SY, Nkamany M, Macdonald KS. Diverse molecular data demonstrate that commercially available medicinal leeches are not Hirudo medicinalis. Proc R Soc B Biol Sci. 2007;274:1481–7.
Simon C, Paabo S, Kocher TD, Wilson AC. Evolution of mitochondrial ribosomal RNA in insects as shown by the polymerase chain reaction. In: Clegg M, Clark S, editors. Molecular evolution, U.C.L.A. Symposia on molecular and CellularBiology, New series, vol. 122. New York: Alan R. Liss, Inc; 1990. p. 235–44.
Soucacos PN, Beris QE, Malizos KN, Xenakis TA, Georgoulis A. Successful treatment of venous congestion in free skin flaps using medical leeches. Microsurgery. 1994;15(7):496–501.
Taşdemir A, Yıldız S, Topkara ET, Özbek M, Balık S, Ustaoğlu MR. Benthic fauna of Yayla Lake (Buldan-Denizli). Türk Sucul Yaşam Dergisi. 2004;2:182–90.
Trontelj P, Utevsky SY. Celebrity with a neglected taxonomy: molecular systematics of the medicinal leech (genus Hirudo). Mol Phylogenet Evol. 2005;34:616–24.
Trontelj P, Utevsky SY. Phylogeny and phylogeography of medicinal leeches (genus Hirudo): Fast dispersal and shallow genetic structure. Mol Phylogenet Evol. 2012;63:475–85.
Trontelj P, Sotler M, Verovnik R. Genetic differentiation between two species of the medicinal leech, Hirudo medicinalis and the neglected H. verbana, based on random-amplified polymorphic DNA. Parasitol Res. 2004;94:118–24. doi:10.1007/s00436-004-1181-x.
Ustaoğlu MR, Balık S, Özbek M, Sarı HM. The Freshwater leeches (Annelida- Hirudinea) of the Gediz catchment area (İzmir region). Zool Middle East. 2003;29:118–20.
Utevsky S, Zagmajster M, Atemasov A, Zınenko O, Utevska O, Utevsky A, Trontelj P. Distribution and status of medicinal leeches (genus Hirudo) in the Western Palaearctic: anthropogenic, ecological, or historical effects? Aquat Conserv Mar Freshwat Ecosyst. 2010;20:198–210.
Utevsky SY, Trontelj P. A new species of the medicinal leech (Oligochaeta, Hirudinida, Hirudo) from Transcaucasia and an identification key for the genus Hirudo. Parasitol Res. 2005;98:61–6.
Whitaker IS, Rao J, Izadi D, Butler PE. Historical Article: Hirudo medicinalis: ancient origins of, and trends in the use of medicinal leeches throughout history. Br J Oral Maxillofac Surg. 2004;42:133–7.
Wirchansky BA, Shain DH. A new species of Haemopis (Annelida: Hirudinea): Evolution of North American terrestrial leeches. Mol Phylogenet Evol. 2010;54:226–34.
Yıldırım N. Fırnız Çayı (Kahramanmaraş)’nın Fiziko-Kimyasal ve Bazı Biyolojik (Bentik makroinvertebrat) Özellikleri. In: Fen Bilimleri Enstitüsü, Biyoliji Ana Bilim Dalı. Kahramanmaraş: Kahramanmaraş Sütçü İmam Ünivrsitesi; 2006. p. 32.
Acknowledgements
We thank Mark Siddall for helpful comments.
Funding
Supported by The Scientific and Technological Research Council of Turkey (TUBITAK) to NS, and Busch Biomedical and GAIA grants to DHS.
Availability of data and material
Holotype and paratype Hirudo sulukii specimens deposited in the Academy of Natural Sciences, Philadelphia, PA, USA (catalogue ANSP G1 19488, 19489). Additional paratypes deposited in Cukurova University Parasitology Museum, Adana, Turkey (catalogue CUPM-HIR/2016-1). All DNA sequences deposited into GenBank (Additional file 1: Table S1).
Authors’ contributions
NS collected leech specimens, conducted experimental analyses including specimen dissections, and contributed to writing the manuscript; RS conducted DNA analyses and contributed to writing; SAL assisted with DNA analyses, phylogenies and writing; DHS oversaw experimental analyses and writing of the manuscript. All authors read and approved the final manuscript.
Competing interests
The authors declare that they have no competing interests.
Consent for publication
Not applicable.
Ethics approval and consent to participate
Ethical approval is not required at Rutgers University for research conducted on invertebrates such as medicinal leeches used in our study.
Endnotes
Not applicable
Author information
Authors and Affiliations
Corresponding author
Additional file
Additional file 1:
Supplementary Data Hirudo. (DOCX 311 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Saglam, N., Saunders, R., Lang, S.A. et al. A new species of Hirudo (Annelida: Hirudinidae): historical biogeography of Eurasian medicinal leeches. BMC Zool 1, 5 (2016). https://doi.org/10.1186/s40850-016-0002-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40850-016-0002-x