
Abstract
Background
Eurasian Collared Dove (Streptopelia decaocto) is a species distributed in the Eurasian continent and North Africa, and inhabiting mainly in Saitama Prefecture in Japan. Eurasian Collared Dove is one of the most prosperous invaders in the world, and Japanese Eurasian Collared Dove has also been introduced from overseas. The Japanese population has declined to one-hundredth over 30 years and is being protected. In this study, we analyzed its genetic diversity in order to understand the genetic differences between wild populations of Eurasian Collared Dove and those bred in zoos.
Methods
A sequence of about 1.9 kb mtDNA was determined for 20 wild Eurasian Collared Doves living in Saitama, Japan and 20 zoo-bred Eurasian Collared Doves, and population genetic analysis was performed.
Results
In the COI gene, 778 bp had the same sequence in all the 40 individuals examined, and no mutation sites could be confirmed. In the control region, two base substitution sites were confirmed in 1140 bp long sequence. Three haplotypes were detected in 20 individuals in wild, whereas all 20 zoo-bred individuals possessed the same haplotypes possessed in the wild population.
Conclusion
Haplotypes of zoo-bred individuals were also retained among the wild individuals, confirming that no genetic problems could occur if the zoo-bred individuals were released to the wild for the Japanese Eurasian Collared Dove propagation program.
Similar content being viewed by others

Background
The Eurasian Collared Dove (Streptopelia decaocto) is a kind of bird classified in the pigeon family Columbidae, and is morphologically the same in both sexes. The Eurasian Collared Dove is not a migratory bird but is strongly dispersed and widely distributed throughout the world (Smith 1987; Romagosa and McEneaney 1999; Owen 2005; Brazil 2009; Gill and Donsker 2019). The country of origin is thought to be India (Del Hoyo et al. 1997), and today it is mainly distributed on the Eurasian continent and North Africa (Smith 1987; Fujisaki et al. 2010). Molecular phylogenetic analysis reports that it is closely related to African Collared Dove (Streptopelia roseogrise; Johnson et al. 2001).
In Japan, the Eurasian Collared Dove is thought to have been introduced artificially from India in the 16th century (The Ornithological Society of Japan 2012) and it inhabits part of the Kanto region (Saitama, Chiba, Ibaraki and Gunma prefectures). As a result of a survey on the population of Eurasian Collared Doves in Saitama Prefecture conducted in 1968, it was estimated that there were 700 individuals in maximum (Kuroda 1969). In 1980, the estimated population was reported to be 10,000 (Saitama Prefecture 1982). However, the population has been decreasing since then, and it was reported that it estimates from 46 to 68 individuals in the 2008 population survey (Konita et al. 2011). Population surveys have continued since then, but the population remains at double digits (Saitama Prefecture web site). The decrease in the number of Eurasian Collared Dove is thought to be due to the destruction of habitats, the decrease in the number of poultry houses that feed, and the thorough control of poultry houses entry (Saitama Prefecture 2013). For this reason, the Eurasian Collared Dove is designated as endangered IB (EN) by the Japanese Ministry of the Environment’s Red List and is protected (Japanese Ministry of the Environment 2014). The low population size of this species raises concerns about reduced genetic diversity. Assessment of the genetic diversity of this species is an urgent task because it is essential to increase genetic diversity as well as population size in order to protect the species at risk. The genetic diversity of the Eurasian Collared Dove in Pakistan and European countries has already been reported using the Cytochrome c oxidase (COI) gene (Awan et al. 2013; Bagi et al. 2018). In this study, to understand the genetic differences between the wild population and the zoo-bred population in Japan, we extracted DNA from the wild population and samples collected from zoos in Saitama Prefecture, and performed analysis by sequencing and comparing mitochondrial DNA (mtDNA).
Methods
Sampling
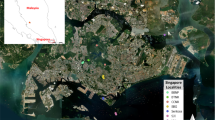
A total of 19 feather samples of the Eurasian Collared Dove used in the present study were collected from the field at seven locations in Saitama Prefecture, Japan, and another 20 feathers were collected from individuals at five zoos in Saitama Prefecture, Japan (Fig. 1). One eggshell of the Eurasian Collared Dove was also obtained from the field in Saitama Prefecture. Wild samples were collected from nests or from the ground within a 3‒4 m radius from just under the nests, and only one sample was collected from each nest to avoid duplication of the same individual. Seven samples were collected in Kuki, six collected in Kawashima, two in Kasukabe and Misato, and one in Kounosu, Satte, and Saitama, respectively. From the zoo, one to nine feather samples were obtained from five zoos. A Ringneck Dove (Streptopelia risoria) sample was also collected from the zoo. All samples were collected between 2017 and 2019, and were sheltered from the sun immediately after collection and were frozen to avoid DNA degradation.

Map of Japan (a) and Saitama Prefecture (b) showing the locations where samples were collected. The number in parenthesis represents the number of samples
DNA extraction, PCR and sequencing
DNA was extracted from the feathers through the spin-column method (NucleoSpin Tissue Kit; MACHEREY–NAGEL, Düren, Germany). The feathers containing soft tissues were cut into small pieces using sterilized scissors, placed in 200 μL of DNA extraction buffer and incubated overnight at 56 °C, resulting to an eluted final volume of 50 μL. For the eggshell sample, the inside of the eggshell was wiped with a cotton swab soaked in DNA extraction buffer and incubated. A partial mtDNA fragment of COI and the Control Region (CR) was amplified through polymerase chain reaction (PCR) using the following primers specifically designed for this study; Sd-COI-F (5′-GTGACCCTAATCAATCGATG-3′) and Sd-COI-R2 (5′-TATGTAGCCGAAGGGTTC-3′) were used for COI, and SD-CR-F15665 (5′-CCCTGCATCTGTGTCCTATG-3′) and SD-CR-R3 (5′-CATTATTAATGGTTTGTCAGCGG-3′) were used for CR, respectively. The mtDNA control region has been widely applied to the population genetics of various animal species including birds because its mutation rate is approximately 2.8–5 times faster than other sequence segments (Avise et al. 1987; Nagata et al. 1998; Avise 2000).
Each amplification process was conducted using a reaction mixture of 20 μL containing 1 μL template DNA, 10 mM deoxynucleotides (2.5 mM each), 5 nM of each primer, 2 μL buffer, and 1 U of Ex Taq polymerase (TaKaRa, Shiga, Japan). The thermal cycling conditions involved an initial denaturation step at 94 °C for 5 min, 45 cycles of denaturation at 95 °C for 10 s, annealing at 57 °C for COI and 62 °C for CR for 30 s, elongation at 72 °C for 60 s, and final elongation at 72 °C for 2 min. The PCR products were confirmed by 2% agarose gel electrophoresis and purified using Nucleo Spin Gel and PCR Clean-up kit (MACHEREY–NAGEL). The purified product was eluted with 20 μL elution buffer. All the amplified fragments were sequenced with both forward and reverse primers. The PCR products were used as the template for the 10-μL cycle sequencing reactions using BigDye Terminator v3.1 Cycle Sequencing Kit (Applied Biosystems, Foster City, CA, USA). The sequences were generated using a four-capillary 3500 Genetic Analyzer (Applied Biosystems) and the specific primer.
Population genetic analyses
Multiple sequence alignment was then performed using CLUSTAL W (Thompson et al. 1994). The sequence waveform data were checked manually to avoid mistakes due to sequence reactions. Haplotype and nucleotide diversities were calculated based on the 1140-bp fragment of the CR using DnaSP version 6.12 (Rozas et al. 2017). The haplotype network was constructed using TCS 1.21 (Clement et al. 2000). The mtDNA CR sequences were deposited in the DNA Data Bank of Japan (DDBJ), European Molecular Biology Laboratory (EMBL), and GenBank under the accession numbers LC529745–LC529747, and LC530631, respectively.
Mismatch distribution analysis (Rogers and Harpending 1998) was performed with DnaSP 6 and Arlequin 3.5 (Excoffier and Lischer 2010) to test for signatures of recent population expansion. DnaSP 6 was used to develop figures and Arlequin 3.5 was used to calculate the values. Tajima’s D (Tajima 1983) and Fu’s FS tests (Fu 1997) of selective neutrality were performed using Arlequin 3.5. Negative Tajima’s D values suggest that a population has recently increased in size. The fit to a sudden expansion model was judged using the value of the sum of squared deviations (SSD). The lack of significant differences in the SSD values indicated that the population assumed an expansion model. The Harpending’s raggedness index (rg) was used to distinguish between expanded and stationary populations with the observed mismatched distributions (Harpending et al. 1993). The value of rg is typically low and inconsequential in expanding populations but high and significant in stationary populations (Harpending 1994). The phylogenetic relationships among the sequences were traced through the maximum likelihood (ML) method (Yang 1994) with MEGA 7 software (Kumar et al. 2016). The ML analysis was performed using the GTR + I + G model. The model was selected with the highest parameter of maximum likelihood fits using the Find Best-Fit Substitution Model of MEGA 7. A closely related S. chinensis and S. orientalis sample (DDBJ/EMBL/NCBI Accession number: KP636801 and NC031447) were used as the outgroup and the previously reported mtDNA of Chinese Eurasian Collared Dove was used for analysis in this study (KX372273). Their corresponding CR sequences were aligned for the phylogenetic analysis.
Results
In this study, we determined a total of 1918 bp of mtDNA from a total of 40 wild and zoo-reared individuals: 778 bp of the COI gene and 1140 bp of the CR. The 778 bp of the COI gene was the same sequence in all 40 individuals, and no mutation sites could be confirmed. In the CR, two base substitution sites were confirmed in 1140 bp, they were transitions, and neither transversion nor indel was confirmed. In the wild, three haplotypes were detected in 20 individuals, whereas all 20 zoo individuals possessed the same haplotype (Table 1). SD-A, SD-B, and SD-C were detected in 6, 11, and 3 individuals, respectively, indicating that SD-B was the major haplotype in wild populations. All zoo animals possessed SD-A.
Phylogenetic analysis based on the obtained sequences showed that the three haplotypes of Eurasian Collared Dove were clustered, and the sequences of Eurasian Collared Dove from China registered in the NCBI database were identical to those of SD-A (Fig. 2). Phylogenetic analysis using CR showed conventional phylogenetic relationships (Johnson et al. 2001). The results of network analysis showed a star-shape formed by a single base difference centered on SD-A, and showed a difference of 9 bases from Ringneck Dove (Fig. 3).

Phylogenetic tree constructed with the haplotypes of the Eurasian Collared Dove using the ML method. Bootstrap values were calculated from 1000 replicates and are indicated at the branches (> 50%). CHN shows a sample of China registered in the database

TCS network tree of three haplotypes among the wild Eurasian Collared Dove in Japan and Ringneck Dove. The numbers in the center of the circles or square represent the haplotype ID number
The haplotype diversity of the wild individual was 0.616 ± 0.077, and the nucleotide diversity was 0.00077 (Table 2). The mismatch distribution analysis for wild showed a unimodal shape (Fig. 4). The SSD value of the wild did not have significant difference. In the selective neutrality test, Tajima’s D and Fu’s FS were neither significantly different.

Mismatch distribution of all haplotypes among the wild Eurasian Collared Dove in Japan
Discussion
This study revealed that wild Eurasian Collared Dove harbors three haplotypes. There was no mutation in the COI gene, only in the CR. This may be due to the fact that the CR generally had a faster base substitution rate than the COI gene, and this was also the case in this species. In contrast to our results, sequencing of 658 bp of the COI gene from 134 individuals of the Eurasian Collared Dove from around Europe revealed 42 haplotypes (Bagi et al. 2018). It is estimated that European Doves have multiplied to approximately 20‒40 million individuals in 50‒60 years, and the high genetic diversity of doves in Europe is due to a remarkable population. The Japanese wild Eurasian Collared Dove had a genetic diversity of 0.616, but the California Condor (Gymnogyps californianus), which is feared to become extinct, was 0.543, and the Eastern White Eagle (Aquila adalberti) was 0.322 (Martinez-Cruz et al. 2004; Adams and Villablanca 2007). Compared with these, the values are not considered to be low. As a result of the mismatch distribution analysis, a unimodal was formed with low nucleotide diversity value, and there was no significant difference in SSD. Furthermore, network analysis showed a star shape. These results indicate that the Eurasian Collared Dove has experienced population expansion in the past, but in fact, the current population has been decimated. This pattern of Eurasian Collared Dove may be the immediate result of a series of demographic events over a very short period of time, such as “introduction–population expansion–population decline—bottleneck”. All 20 zoo-bred individuals possessed the same haplotype. All the animals currently bred at the zoo excluding zoo E are from the Saitama Prefectural Agriculture and Forestry Research Center, and were bred from four individuals caught in 1966 (Saitama Eurasian Collared Dove Protection Plan). Therefore, if it is inferred that the four individuals were derived from the same maternal line, and there is no particular contradiction with this result. The breeding population of Zoo L breeds wild individuals, and the founder individuals differ from other zoos. In this study, it was found that they had the same haplotype as those raised in other zoos. The major haplotype of the wild population was SD-B, and all zoo-bred individuals retained SD-A. If all zoo-bred individuals were released into the wild, the major haplotype in wild would be SD-A, but even in wild, SD-A is not numerous, so there seems to be no particular problem as genetic pollution. Since only three SD-Cs in the wild population are detected, protection of the haplotype SD-C is an issue that needs to be given the highest priority in order to maintain the genetic diversity of Eurasian Collared Dove. In the future, it will be necessary to maintain the population so as not to further reduce the population of the Eurasian Collared Dove, and to secure a founder holding SD-C and propagate it in a zoo for protection of this dove species. One of the most important factors of the population decrease of the Eurasian Collared Dove may be that the invasion of wild birds into poultry houses has been controlled by avian influenza countermeasures (Saitama Prefecture 2013). Providing such an environment artificially may be necessary for the protection of Eurasian Collared Dove. It should be interpreted with some caution that only maternally derived mtDNA was used in this study. Future microsatellite genotyping may elucidate the genetic diversity of Eurasian Collared Dove in detail.
Conclusion
We sequenced the 778 bp COI gene and the 1140 bp CR in 20 wild Eurasian Collared Doves and 20 zoo-bred in Japan. Comparative analysis revealed three haplotypes from wild individuals in CR, and all zoo-bred individuals had the same haplotype. Also, the haplotypes of the zoo-bred individuals were retained in wild individuals. Phylogenetic analysis revealed that the three haplotypes were genetically related and had the same DNA sequence as Chinese Eurasian Collared Dove individual. In addition, the study confirms that no genetic problems as genetic pollution can occur if a zoo-bred individual is released into the wild for a Japanese Eurasian Collared Dove propagation program.
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
References
Adams MS, Villablanca FX. Consequences of a genetic bottleneck in California condors: a mitochondrial DNA perspective. In: Mee A, Hall LS, editors. California condors in the 21st century. Nuttall Ornithological Club and American Ornithologist’s Union, Cambridge. 2007. p. 35–55.
Avise JC. Phylogeography: the history and formation of species. Cambridge: Harvard University Press; 2000.
Avise JC, Arnold J, Ball RM. Intraspecific phylogeography: the mitochondrial DNA bridge between population genetics and systematics. Ann Rev Ecol Syst. 1987;18:489–522.
Awan AR, Umar E, Haq MZ, Firyal S. Molecular classification of Pakistani collared dove through DNA barcoding. Mol Biol Rep. 2013;40:6329–31.
Bagi Z, Dimopoulos EA, Loukovitis D, Eraud C, Kusza S. MtDNA genetic diversity and structure of Eurasian Collared Dove (Streptopelia decaocto). PLoS ONE. 2018;13:e0193935.
Brazil M. Birds of East Asia. Princeton: Princeton University Press; 2009.
Clement MD, Posada D, Crandall KA. TCS: a computer program to estimate gene genealogies. Mol Ecol. 2000;9:1657–9.
Del Hoyo J, Elliot A, Sargatal J. Handbook of the birds of the world. Sandgrouse to cuckoos, vol. 4. Barcelona: Lynx Edicions; 1997.
Excoffier L, Lischer HEL. Arlequin suite ver 3.5: a new series of programs to perform population genetics analyses under Linux and Windows. Mol Ecol Resour. 2010;10:564–7.
Fujisaki I, Pearlstine EV, Mazzotti FJ. The rapid spread of invasive Eurasian collared doves Streptopelia decaocto in the continental USA follows human altered habitats. Ibis. 2010;152:622–32.
Gill F, Donsker D. IOC world bird list (v9.1). 2019. https://doi.org/10.14344/ioc.ml.9.1.
Harpending HC. Signature of ancient population growth in a low-resolution mitochondrial DNA mismatch distribution. Hum Biol. 1994;66:591–600.
Harpending HC, Sherry ST, Rogers AR, Stoneking M. The genetic structure of ancient human populations. Curr Anthropol. 1993;34:483–96.
Johnson KP, de Cort S, Dinwoodey K, Mateman AC, ten Cate C, Lessells CM, et al. A molecular phylogeny of the dove genera Streptopelia and Columba. Auk. 2001;118:874–87.
Konita Y, Ura T, Ikeno S, Kawachi H, Takahashi K, Wakatabe S, Mizuno A. Density of the Eurasian Collared Dove Streptopelia decaocto of the Kanto region in 2008 in comparison with historical records. Strix. 2011;27:135–40 (in Japanese).
Kumar S, Stecher G, Tamura K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol Biol Evol. 2016;33:1870–4.
Kuroda N. Recovery of Japanese Streptopelia decaocto from a reduced small population. J Yamashina Inst Ornithol. 1969;49:131–42 (in Japanese).
Martınez-Cruz B, Godoy JA, Negro JJ. Population genetics after fragmentation: the case of the endangered Spanish imperial eagle (Aquila adalberti). Mol Ecol. 2004;13:2243–55.
Ministry of the Environment. Red data book 2014—Threatened wildlife of Japan, vol. 2. Aves. Tokyo: Gyosei; 2014 (in Japanese).
Nagata J, Masuda R, Kaji K, Kaneko M, Yoshida MC. Genetic variation and population structure of the Japanese sika deer (Cervus nippon) in Hokkaido Island based on mitochondrial D-loop sequences. Mol Ecol. 1998;7:871–7.
Owen J. The ecology of a garden: The first fifteen years. Cambridge: University Press; 2005.
Rogers AR, Harpending H. Population growth waves in the distribution of pairwise genetic differences. Mol Biol Evol. 1998;9:552–69.
Romagosa CM, McEneaney T. Eurasian collared dove in North America and the Caribbean. N Am Birds. 1999;53:348–53.
Rozas J, Ferrer-Mata A, Sánchez-DelBarrio JC, Guirao-Rico S, Librado P, Ramos-Onsins SE, et al. DnaSP 6: DNA sequence polymorphism analysis of large datasets. Mol Biol Evol. 2017;34:3299–302.
Saitama Prefecture. Urgent investigation report of natural monuments: Eurasian Collared Dove in Koshigaya. Saitama Prefecture Historic Site, Place of Scenic Beauty and Natural Monument Survey Report Vol.3 1982. (in Japanese).
Saitama Prefecture. Saitama Eurasian Collared Dove conservation plan. 2013. https://www.pref.saitama.lg.jp/a0508/sub-tayouseihozen/documents/614427.pdf. Accessed 15 Oct 2019. (in Japanese).
Smith PW. The Eurasian Collared-dove arrives in the Americas. Am Birds. 1987;41:1371–9.
Tajima F. Evolutionary relationship of DNA sequences in finite populations. Genetics. 1983;105:437–60.
The Ornithological society of Japan. Check-list of Japanese birds. 7th revised edition. Sanda: The Ornithological Society of Japan; 2012.
Thompson JD, Higgins DG, Gibson TJ. CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994;22:4673–80.
Yang Z. Maximum likelihood phylogenetic estimation from DNA sequences with variable rates over sites: approximate methods. J Mol Evol. 1994;39:306–14.
Acknowledgements
This study was conducted as part of a survey in Saitama Prefecture. The authors also thank the anonymous reviewers and editor for their expert criticism and valuable suggestions.
Funding
Not applicable.
Author information
Authors and Affiliations
Contributions
KN performed the experiments, analyzed the data, prepared the draft, figures and/or tables, KT, KU, YT, TA and FN collected samples, FN conceived and designed the study, and improved the drafts. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
This study followed all applicable institutional guidelines for the care and use of animals and complied with the current laws of the country in which they were performed.
Consent for publication
Not applicable.
Competing interests
The authors have no conflicts of interest to report.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Nagai, K., Takahashi, Y., Tokita, Ki. et al. Genetic diversity in Japanese populations of the Eurasian Collared Dove. Avian Res 11, 21 (2020). https://doi.org/10.1186/s40657-020-00207-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40657-020-00207-8