
Abstract
Background
Feathers are a defining feature of birds with multiple functions such as flight, insulation, protection against predation and signaling. Feathers are lost during the annual molt while the rate of such loss at other times of the year and its fitness consequences remain poorly known.
Methods
I used information on the number and the mass of feathers for 160 individuals belonging to 85 species of birds in general linear mixed models to analyze allometry of feathers and to investigate possible factors explaining variation in the number of feathers. A phylogenetic effect was assessed by quantifying the random effect of genus.
Results
The total mass of feathers increased isometrically with body mass, while the total number of feathers and the mean mass of feathers showed negative allometry. Negative allometry implied that small-sized species had relatively many small feathers. There was a negative association between the number of feathers and migration distance. The total number of feathers initially increased during fall and winter, consistent with individuals growing more feathers later during the year or with individuals with fewer feathers selectively disappearing from the population. In contrast, the number of feathers decreased from winter through spring and summer.
Conclusions
These findings suggest that thermoregulation has affected the evolution of the number and the size of feathers, there is selection against feather loss, and that the number of feathers varies across seasons.
Similar content being viewed by others

Background
The number of feathers varies among species of birds from less than 1000 to more than 10000 (Wetmore 1936; Brodkorb 1985), although the factors accounting for this variation remain poorly known. Feathers have a number of functions because they provide insulation (Whittow 1986), protect against predators (Dathe 1955; Mester and Pünte 1959; Tautenhahn 1959; Berger 1960; Höglund 1964; Lindström and Nilsson 1988; Møller et al. 2006), allow for flight and contribute to signalling through size, shape and color (Darwin 1871; Andersson 1982). Down are smaller than contour feathers, but equally important for the maintenance of a high body temperature (Dyck 1985). Several studies have shown that seasonal changes in energy management strategies are related to variation in plumage weight (Saarela et al. 1984; Dove et al. 2007). Furthermore seasonal changes in plumage weight suggest better insulation during colder months (Dawson and Carey 1976; Dawson et al. 1983; Swanson 1991; Cooper 2002). Finally, feather structure could be trade against the number of feathers (Dawson and Carey 1976; Middleton 1986; Walsberg 1988; Lindström, et al. 1993; Dawson et al. 2000; Wolf and Walsberg 2000).
If there is gradual loss of feathers during the annual cycle following the complete molt, we should expect a gradual reduction over time. Brodkorb (1985) reported that house sparrows Passer domesticus loose 11.5% of their feathers from winter to summer, although the data were cross-sectional rather than longitudinal. Hence these data do not allow separation of phenotypic plasticity from selective disappearance. Here I propose predictions that would help discriminate between alternative explanations for seasonal change in the number of feathers. If individuals with more feathers do better in terms of insulation than individuals with few feathers, we should expect an increase in the number of feathers during fall and winter as temperatures fall. If feathers provided protection against predators by allowing for escape, we should expect an increase in the number of feathers over time because individuals that have lost feathers due to predation were more likely to subsequently be captured by a predator or die from insufficient insulation. If the sole advantage of more feathers was simply to provide better insulation, we should expect isometry. If there were relatively more feathers on small species due to their high surface to volume ratio, we should expect negative allometry for the number of feathers. If the total mass of feathers showed isometry, we should expect that the number of feathers and the mean mass of feathers should show isometry.
The objectives of this study were to (1) estimate the allometry of number, total mass of the plumage and mean mass of feathers and (2) to model changes in the number of feathers during the annual cycle to better understand the temporal patterns of feather loss. The number of feathers should reach a maximum at the end of molt, when all feathers including missing ones have been replaced, the only exception being feather loss at the stage of molt. Any subsequent loss of feathers following molt should result in a reduction in the total number of feathers. In contrast, a subsequent increase in the number of feathers later during the year would only be possible if individuals with more feathers survived better than individuals with fewer feathers. Alternatively, individuals may grow more feathers during fall and winter although there is no empirical evidence so far for such seasonal growth of feathers. Thus, a seasonal increase in the number of feathers would represent an estimate of the selective advantage of having more feathers with individuals with few feathers disappearing from the population. The data used for this study mainly derived from Wetmore (1936), who pioneered the study of abundance of feathers on birds.
Methods
Data
Data on the number of feathers and their total mass were extracted from Wetmore (1936) combined with smaller samples from Staebler (1941), Brodkrob (1955) and Markus (1965). Although the mass of different types of feathers would have been useful, no such information was available in the sources consulted here.
I extracted information on body mass from Dunning (2008). Migration distance was estimated as the mean of the northernmost and southernmost latitudes during the breeding season minus the mean of the northernmost and southernmost latitudes during winter relying on Cramp and Perrins (1977–1994) and Poole et al. (1993–2002). I also used northernmost and southernmost latitudes during the breeding season and northernmost and southernmost latitudes during winter to test for possible latitudinal effects. These tests were restricted to North American species to avoid problems of differences in latitudinal effects on climate among continents. The species investigated here generally molt in late summer-early fall (Cramp and Perrins 1977–1994; Poole et al. 1993–2002). Therefore, I related the total number of feathers to time since molt estimated as the number of months since August. All data are reported in Additional file 1: Table S1.
Statistical analyses
I log10-transformed body mass, total number of feathers, mass of all feathers, mean mass of feathers and migration distance to achieve normally distributed frequency distributions.
I analyzed allometric relationships between the total mass of all feathers, the total number of feathers and the mean mass of feathers, respectively, in relation to body mass using log10-transformed variables while including species as a random effect to account for differences in sample size among species. I tested whether the allometric relationships were isometric or negative or positive allometric by testing with t-tests whether allometry coefficients differed significantly from unity.
I developed general linear mixed models with species as a random effect thereby controlling for differences in sample size among species while body mass, time since molt and migration distance were fixed factors. Both a linear and a quadratic effect of time since molt were entered into the models to account for non-linear effects of relative time. Furthermore, I included sex and northernmost and southernmost breeding and wintering latitude in these general linear mixed models with species as a random effect thereby controlling for differences in sample size among species while body mass, time since molt and migration distance were fixed factors. Finally, I tested for phylogenetic effects in a general linear mixed model with species and genus as random effects controlling for differences in sample size among species while body mass, time since molt and migration distance were fixed factors. A phylogenetic effect would imply a significant effect of genus. All analyses were made with JMP (SAS 2012).
Results
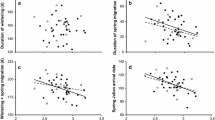
A total of 91% of the variance in the number of feathers was due to differences among species with the remaining 9% of the variance occurring within species (F = 9.42, d.f. = 84, 75, r 2 = 0.91, p < 0.0001). The allometric relationships for the number of feathers, the mass of the plumage and the mean mass of feathers are reported in Table 1. The total mass of all feathers increased isometrically with body mass since the allometry coefficient did not differ significantly from one (t = 0.77, d.f. = 74.46, p = 0.45). The total number of feathers showed negative allometry (Figure 1A), with an allometry coefficient less than one (t = 24.47, d.f. = 85.64, p < 0.001). The average mass of individual feathers likewise increased with body mass showing negative allometry (Figure 1B; Table 2), and the allometry coefficient was less than one (t = 5.27, d.f. = 73.1, p < 0.001).

(A) The number of feathers in different birds in relation to body mass (g), and (B) the mean mass of feathers in different birds in relation to body mass (g).
The relationship between the number of feathers, total mass of feathers and mean mass of feathers and log migration distance was significantly negative for number of feathers, but not for mass of feathers or mean mass of feathers (no. feathers: F = 8.60, d.f. = 1, 81, p = 0.044, estimate (SE) = −0.075 (0.026); mass of feathers: F = 0.00, d.f. = 1, 60.74, p = 0.98; mean mass of feathers: F = 0.14, d.f. = 1, 60.67, p = 0.71). Thus species that migrated longer distances had fewer feathers for a given body size. The total number of feathers was significantly related to time since molt and time since molt squared (Table 2). This implies that the number of feathers reached a peak halfway through the year after molt (Figure 2). In addition there was a significant effect of body mass with more feathers in larger species (Table 2).

Total number of feathers in relation to time (months since August) in different birds. Note the curvilinear relationship.
The predicted number of feathers at the end of the molting period from the model in Table 2 was 1259, increasing to 1950 half a year later (a rate of increase by 3.8 feathers per day), and subsequently falling to 1350 by the start of the molting period 12 months later (a rate of loss of 3.3 feathers per day).
There was no significant effect of sex in the analysis presented in Table 2 (F = 0.42, d.f. = 1, 142, p = 0.52). Furthermore, there were no significant effects of northernmost or southernmost latitude during breeding or during winter (breeding northern latitude: F = 1.07, d.f. = 1, 63.33, p = 0.30; breeding southern latitude: F = 1.49, d.f. = 1, 84.81, p = 0.23; winter northern latitude: F = 0.22, d.f. = 1, 68.87, p = 0.64; winter southern latitude: F = 0.29, d.f. = 1, 87.44, p = 0.59). Finally, there was no effect of genus on the number of feathers in a mixed model (variance ratio = 0.088, 95% confidence intervals −0.00038, 0.00066, percentage variance explained = 6.23).
Discussion
The number of feathers showed negative allometry while the mass of feathers showed isometry. The number of feathers showed a hump-shaped relationship with time since molt in August, first showing an increase during fall and winter followed by a decrease during spring and summer. This pattern could be explained as a consequence of selection against feather loss during fall and winter followed by a steady loss during spring and summer. Here I have only analyzed patterns of abundance of feathers and feather loss in a sample of mainly passerine and related birds. Thus the results provided here do not necessarily apply to other orders of birds.
Feathers play an important role in thermoregulation. Here I have shown that the total mass of feathers increased isometrically with body mass. In contrast, the total number of feathers and the mean mass of feathers showed negative allometry implying that small species have relatively more and light feathers. These latter patterns are consistent with expectations from the higher surface to volume ratio of smaller species. Furthermore, they suggest that smaller species have relatively many small feathers.
I did not find a linear decline in the number of feathers from the molting period onwards, raising questions about gain and loss of feathers. Feathers emerge during the annual (or bi-annual) molt, and once the molting period is finished, there is little or no feather growth with the exception of feathers being replaced if completely lost. Such replacement is used as a research tool in studies of ptilochronology (Grubb 2006). An alternative explanation for an increase in the number of feathers during fall and winter is that birds grow more feathers during fall and winter although there is no empirical evidence for such seasonal growth. Hence there appears to be no means by which the number of feathers can increase during fall and winter unless individuals that have already lost feathers are selected against. Here, I have indirectly shown selection for individuals with a larger number of feathers in fall because the number of feathers increased with time during fall, followed by gradual loss of feathers in spring as the importance of thermoregulation diminished when ambient temperatures increased. There was a negative association between the number of feathers and migration distance, while there was no significant relationship between the mass of feathers and the mean mass of individual feathers, respectively, and migration distance. Thus migrants had fewer feathers than residents as expected if the cost of thermoregulation is higher in resident species at higher latitudes. In contrast, there were no significant associations between the number of feathers and northernmost or southernmost latitudinal range during breeding or winter. Gradual loss of feathers in spring and summer when ambient temperatures increase allows for the maintenance of high body temperature in the face of normal activity. Such feathers lost during spring and summer may be used as nest material and provide efficient insulation for developing embryos and nestlings.
Feathers are commonly lost in connection with predation attempts (Dathe 1955; Mester and Pünte 1959; Tautenhahn 1959; Berger 1960; Höglund 1964; Lindström and Nilsson 1988; Møller et al. 2006). The frequency of such loss differs among species that vary in susceptibility to predation with the most predation-prone species more readily losing feathers (Møller et al. 2006). The ease of feather loss even differs between neighboring urban and rural populations (Møller and Ibáñez-Álamo 2012), implying that there is scope for local adaptation in terms of feather loss among habitats with different predator communities.
The present study raises a number of perspectives for the future. First, are the patterns in non-passerines similar to what has been described here? Second, do tropical species have fewer feathers than temperate species as a consequence of their higher ambient temperature? Third, is feather loss and number of feathers related to susceptibility to predation? Fourth, what is the mechanism that maintains the balance between difficulty of loss of feathers to maintain flying ability and insulation and ease of loss when attacked by a predator? Finally, how does this balance change as feathers become worn and lost prior to molt?
Conclusion
Larger species of birds have more and heavier feathers, and these feathers are lost at a rate that indicates selection against feather loss in fall and winter, while the rate of feather loss remains constant during spring and summer with the number of feathers reaching its lowest level in summer just before the start of molt.
References
Andersson M (1982) Female choice selects for extreme tail length in a widowbird. Nature 299:818–820
Berger G (1960) Schreckmauser bei der Grosstrappe (Otis tarda L.). Beitr Vogelkde 7:126–129
Brodkorb P (1955) Number of feathers and weights of various systems in a Bald Eagle. Wilson Bull 67:142
Brodkorb P (1985) Feathers, number of. In: Campbell B, Lack E (eds) A Dictionary of Birds. T & AD Poyser, London, pp 209–210
Cooper SJ (2002) Seasonal metabolic acclimatization in mountain chickadees and juniper titmice. Physiol Biochem Zool 75:386–395
Cramp S, Perrins CM (eds) (1977–1994) The Birds of the Western Palearctic, Vols. 1–9. Oxford University Press, Oxford, UK
Darwin C (1871) The Descent of Man, and Selection in Relation to Sex. John Murray, London, UK
Dathe H (1955) Über die Schreckmauser. J Ornithol 96:5–14
Dawson WR, Carey C (1976) Seasonal acclimatization to temperature in cardueline finches. I. Insulative and metabolic adjustments. J Comp Physiol 112:317–333
Dawson WR, Marsh RL, Buttemer WA, Carey C (1983) Seasonal and geographic variation of cold resistance in house finches Carpodacus mexicanus. Physiol Zool 56:353–369
Dawson A, Hinsley SA, Ferns PN, Bonser RHC, Eccleston L (2000) Rate of moult affects feather quality: a mechanism linking current reproductive effort to future survival. Proc R Soc Lond B 267:2093–2098
Dove CJ, Rijke AM, Wang X, Andrews LS (2007) Infrared analysis of contour feathers: the conservation of body heat radiation in birds. J Therm Biol 32:42–47
Dunning JB (2008) CRC Handbook of Avian Body Masses, 2nd edn. CRC Press, Boca Raton, FL
Dyck J (1985) Feather. In: Campbell B, Lack E (eds) A Dictionary of Birds. T & AD Poyser, London, pp 206–209
Grubb TC Jr (2006) Ptilochronology: Feather Tme and the Biology of Birds. Oxford University Press, New York
Höglund NH (1964) Fright moult in Tetraonids. Viltrevy 2:419–422
Lindström Å, Nilsson J-Å (1988) Birds doing it the octopus way: fright moulting and distraction of predators. Ornis Scand 19:165–166
Lindström Å, Visser GH, Daan S (1993) The energetic cost of feather synthesis is proportional to basal metabolic rate. Physiol Zool 66:490–510
Markus MB (1965) The number of feathers on birds. Ibis 107:394
Mester H, Pünte W (1959) Bemerkungen über die Schreckmauser. Vogelwelt 80:179–180
Middleton ALA (1986) Seasonal changes in plumage structure and body composition of the American goldfinch, Carduelis tristis. Can Field-Nat 100:545–549
Møller AP, Ibáñez-Álamo JD (2012) Escape behaviour of birds provides evidence of predation being involved in urbanization. Anim Behav 84:341–348
Møller AP, Nielsen JT, Erritzøe J (2006) Losing the last feather: feather loss as an antipredator adaptation in birds. Behav Ecol 17:1046–1056
Poole A, Stettenheim P, Gill F (eds) (1993–2002) The birds of North America. The American Ornitologists’ Union and the Academy of Natural Sciences of Philadelphia, Philadelphia
Saarela S, Rintamäki H, Saarela M (1984) Seasonal variation in the dynamics of ptiloerection and shivering correlated changes in the metabolic rate and body temperature of the pigeon. J Comp Physiol B 154:47–53
SAS (2012) JMP version 10.0. SAS Institute Inc, Cary, NC
Staebler AE (1941) Number of contour feathers in the English sparrow. Wilson Bull 53:126–127
Swanson DL (1991) Seasonal adjustments in metabolism and insulation in the dark-eyed junco. Condor 93:538–545
Tautenhahn W (1959) Schreckmauser bei Amsel. Vogelwelt 80:122
Walsberg GE (1988) Heat flow through avian plumages: the relative importance of conduction, convection, and radiation. J Therm Biol 13:89–92
Wetmore A (1936) The number of feathers in passeriform and related birds. Auk 53:159–169
Whittow GC (1986) Regulation of body temperature. In: Sturkie PD (ed) Avian Physiology. Springer, New York, pp 221–252
Wolf BO, Walsberg GE (2000) The role of the plumage in heat transfer processes of birds. Am Zool 40:575–584
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The author declares that he has no competing interests.
Additional file
Additional file 1: Table S1.
Species, genus, migration distance (° latitude), month, sex, number of feathers, mass of feathers (g), body mass (g), number of months after molt, northernmost breeding latitude, southernmost breeding latitude, northernmost winter latitude, and southernmost winter latitude in different species of birds. See Methods for further details.
Rights and permissions
This article is published under an open access license. Please check the 'Copyright Information' section either on this page or in the PDF for details of this license and what re-use is permitted. If your intended use exceeds what is permitted by the license or if you are unable to locate the licence and re-use information, please contact the Rights and Permissions team.
About this article

Cite this article
Møller, A.P. The allometry of number of feathers in birds changes seasonally. Avian Res 6, 2 (2015). https://doi.org/10.1186/s40657-015-0012-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40657-015-0012-3